
The Obesity Amelioration Effect in High-Fat-Diet Fed Mice of a Homogeneous Polysaccharide from Codonopsis pilosula

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
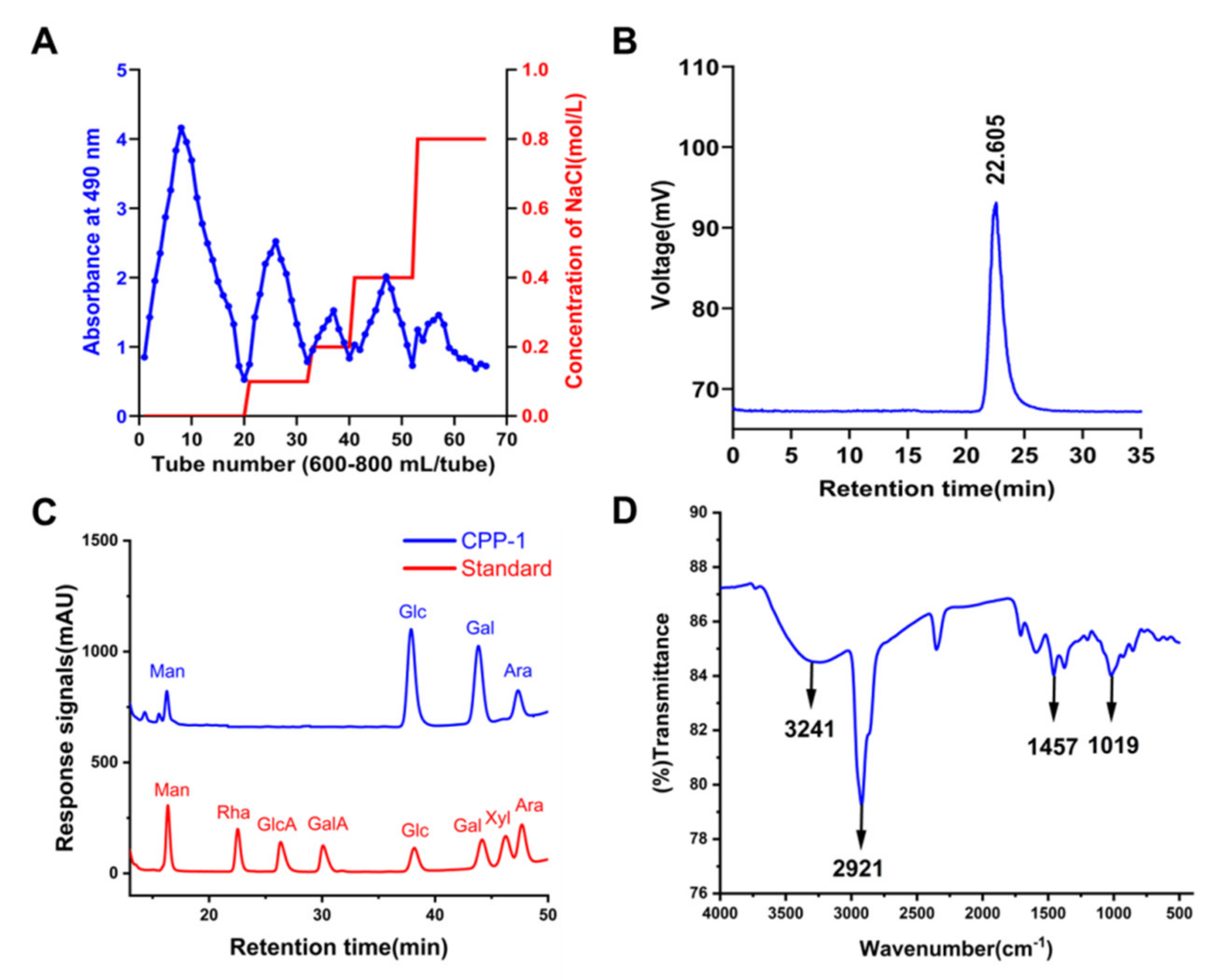
2.1. Extraction, Isolation, and Purification of CPP−1
2.2. Monosaccharide Composition Analysis of CPP−1
2.3. Methylation Analysis
2.4. Fourier Transform Infrared Spectroscopy (FT-IR) Analysis
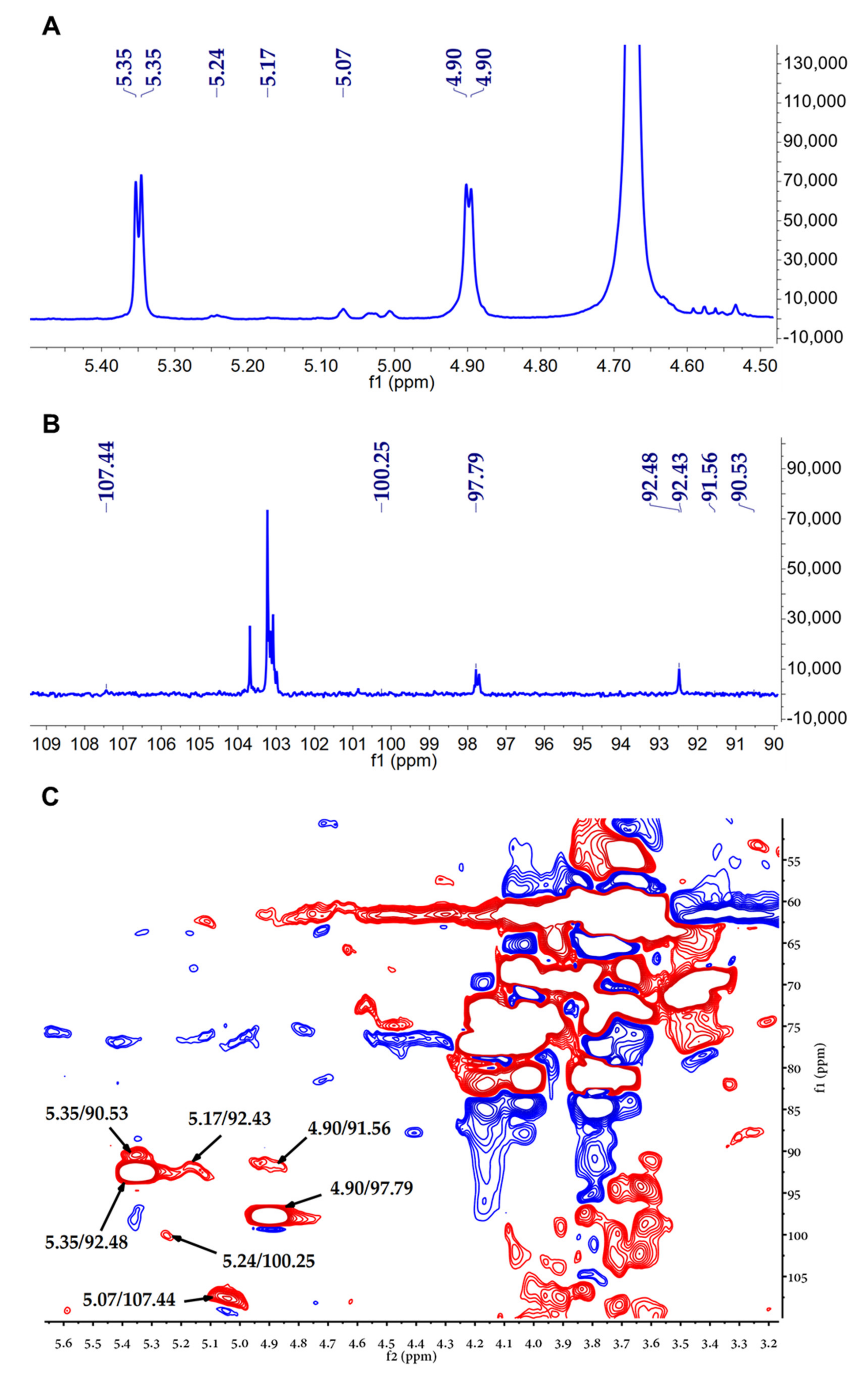
2.5. Nuclear Magnetic Resonance (NMR) Spectroscopy
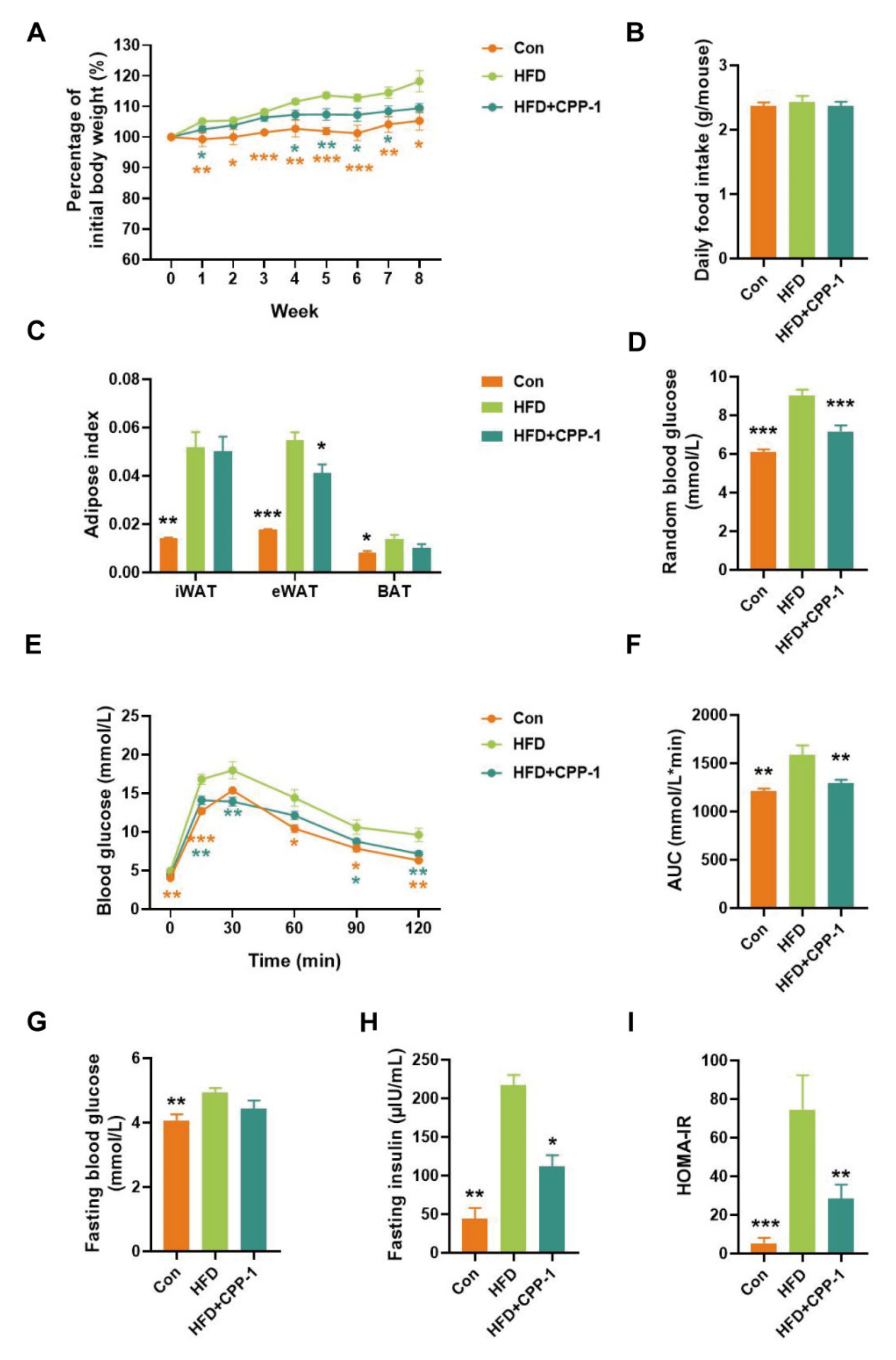
2.6. Effect of CPP−1 Supplementation on Body Weight Gain in HFD-Fed Mice
2.7. CPP−1 Supplementation Improves Blood Glucose Metabolism
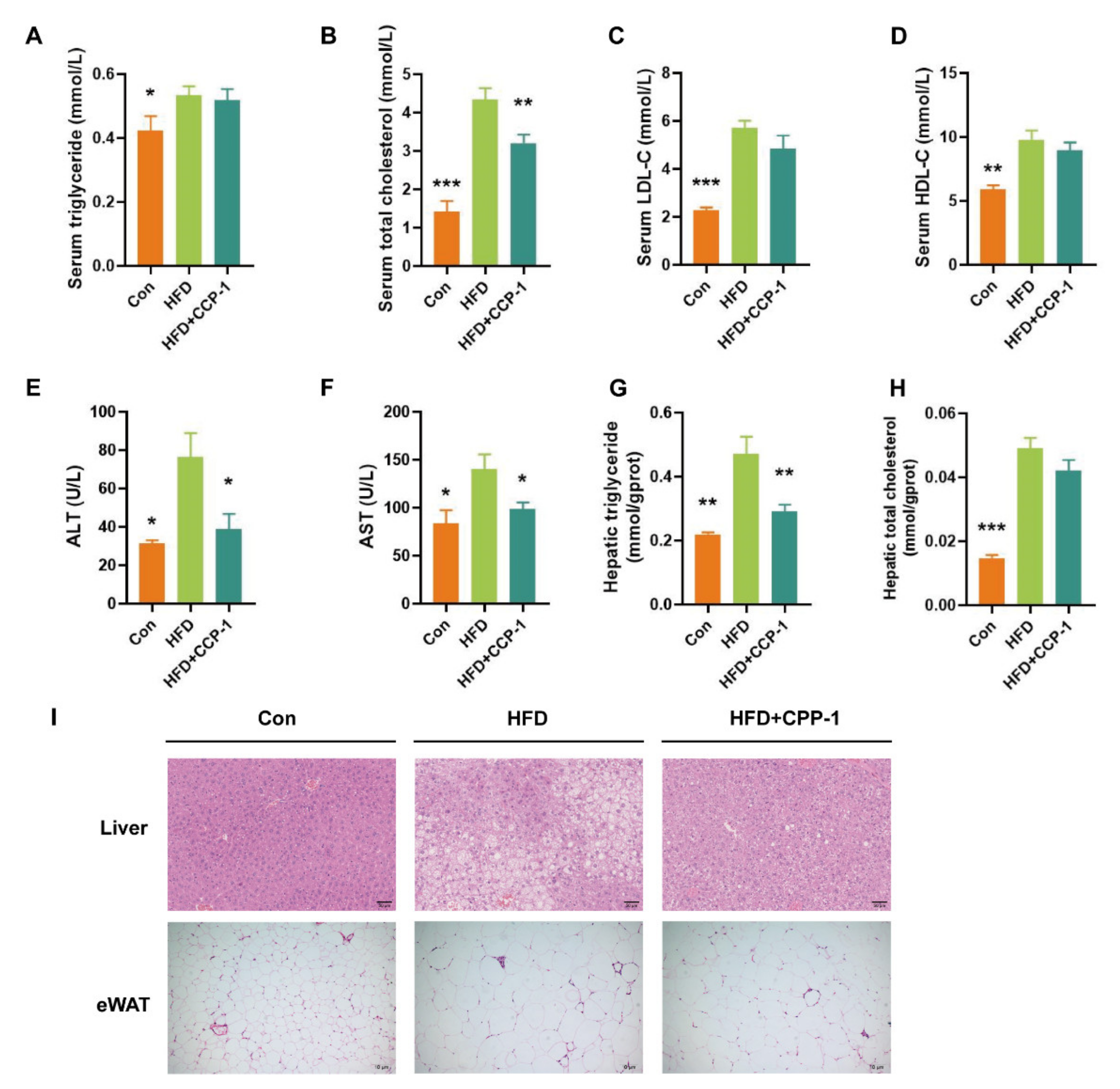
2.8. CPP−1 Supplementation Improves Lipid Metabolism
2.9. CPP−1 Supplementation Alleviates Lipid Accumulation in the Liver and eWAT
3. Materials and Methods
3.1. Materials
3.2. Extraction, Purification and Fractionation of Polysaccharides
3.3. Homogeneity Evaluation and Molecular Weight Determination
3.4. Monosaccharide Composition
3.5. Fourier-Transform Infrared (FT-IR) Spectroscopy
3.6. Methylation Analysis
3.7. Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
3.8. Animal Studies
3.9. Random Glucose Test
3.10. Glucose Tolerance Test
3.11. Fasting Blood and Fasting Insulin Assays
3.12. Biochemical Analyses
3.13. Histology and Microscopy
3.14. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Endalifer, M.L.; Diress, G. Epidemiology, Predisposing Factors, Biomarkers, and Prevention Mechanism of Obesity: A Systematic Review. Int. J. Obesity 2020, 2020, 6134362. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, E. Obesity and cardiovascular disease. Minerva Pediatrics 2015, 67, 25–32. [Google Scholar]
- Han, T.S.; Lean, M.E. A clinical perspective of obesity, metabolic syndrome and cardiovascular disease. JRSM Cardiovasc. Dis. 2016, 5, 2048004016633371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almendros, I.; Martinez-Garcia, M.A.; Farré, R.; Gozal, D. Obesity, sleep apnea, and cancer. Int. J. Obesity 2020, 44, 1653–1667. [Google Scholar] [CrossRef]
- Vink, R.G.; Roumans, N.J.T.; Arkenbosch, L.A.J.; Mariman, E.C.M.; van Baak, M.A. The effect of rate of weight loss on long-term weight regain in adults with overweight and obesity. Obesity 2016, 24, 321–327. [Google Scholar] [CrossRef]
- Bessesen, D.H.; Van Gaal, L.F. Progress and challenges in anti-obesity pharmacotherapy. Lancet Diabetes Endocrinol. 2018, 6, 237–248. [Google Scholar] [CrossRef]
- Su, Y.; Li, J.; Wu, L.; Kuang, H. Polysaccharides from TCM herbs exhibit potent anti-obesity effect by mediating the community structure of gut microbiota. Die Pharm. 2021, 76, 473–479. [Google Scholar]
- Zhang, Y.; Xie, Q.; You, L.; Cheung, P.C.K.; Zhao, Z. Behavior of Non-Digestible Polysaccharides in Gastrointestinal Tract: A Mechanistic Review of its Anti-Obesity Effect. eFood 2021, 2, 59–72. [Google Scholar] [CrossRef]
- Lee, H.B.; Kim, Y.S.; Park, H.Y. Pectic polysaccharides: Targeting gut microbiota in obesity and intestinal health. Carbohydr. Polym. 2022, 287, 119363. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Jin, M.L.; Morris, G.A.; Zha, X.Q.; Chen, H.Q.; Yi, Y.; Li, J.E.; Wang, Z.-J.; Gao, J.; Nie, S.-P. Advances on bioactive polysaccharides from medicinal plants. Crit. Rev. Food Sci. 2016, 56, S60–S84. [Google Scholar] [CrossRef]
- Yang, Y.; Chang, Y.; Wu, Y.; Liu, H.; Liu, Q.; Kang, Z.; Wu, M.; Yin, H.; Duan, J. A homogeneous polysaccharide from Lycium barbarum: Structural characterizations, anti-obesity effects and impacts on gut microbiota. Int. J. Biol. Macromol. 2021, 183, 2074–2087. [Google Scholar] [CrossRef]
- Li, R.; Xue, Z.; Jia, Y.; Wang, Y.; Li, S.; Zhou, J.; Liu, J.; Zhang, M.; He, C.; Chen, H. Polysaccharides from mulberry (Morus alba L.) leaf prevents obesity by inhibiting pancreatic lipase in high-fat diet induced mice. Int. J. Biol. Macromol. 2021, 192, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Meng, C.; Chen, W.; Chen, H.; Chen, W. Ganoderma amboinense polysaccharide prevents obesity by regulating gut microbiota in high-fat-diet mice. Food Biosci. 2021, 42, 101107. [Google Scholar] [CrossRef]
- Cheng, Z.; Hu, M.; Tao, J.; Yang, H.; Yan, P.; An, G.; Wang, H. The protective effects of Chinese yam polysaccharide against obesity-induced insulin resistance. J. Funct. Foods 2019, 55, 238–247. [Google Scholar] [CrossRef]
- Bai, R.; Cui, F.; Li, W.; Wang, Y.; Wang, Z.; Gao, Y.; Wang, N.; Xu, Q.; Hu, F.; Zhang, Y. Codonopsis pilosula oligosaccharides modulate the gut microbiota and change serum metabolomic profiles in high fat diet induced obese mice. Food Funct. 2022, 13, 8143–8157. [Google Scholar] [CrossRef]
- Liu, W.; Lv, X.; Huang, W.; Yao, W.; Gao, X. Characterization and hypoglycemic effect of a neutral polysaccharide extracted from the residue of Codonopsis Pilosula. Carbohydr. Polym. 2018, 197, 215–226. [Google Scholar] [CrossRef]
- Hu, Y.R.; Xing, S.L.; Chen, C.; Shen, D.Z.; Chen, J.L. Codonopsis pilosula polysaccharides alleviate Aβ1-40-induced PC12 cells energy dysmetabolism via CD38/NAD+ signaling pathway. Curr. Alzheimer Res. 2021, 18, 208–221. [Google Scholar]
- Li, J.; Zhang, X.; Cao, L.; Ji, J.; Gao, J. Three inulin-type fructans from Codonopsis pilosula (Franch.) Nannf. Roots and their prebiotic activity on Bifidobacterium longum. Molecules 2018, 23, 3123. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Du, C.; Zhai, X.; Li, J.; Meng, J.; Shao, Y.; Gao, J. Codonopsis pilosula Polysaccharide Improved Spleen Deficiency in Mice by Modulating Gut Microbiota and Energy Related Metabolisms. Front. Pharmacol. 2022, 13, 862763. [Google Scholar] [CrossRef]
- Engin, A. Obesity and Lipotoxicity; Springer Cham: New York, NY, USA, 2017; pp. 1–17. [Google Scholar]
- Gao, S.M.; Liu, J.S.; Wang, M.; Cao, T.T.; Qi, Y.D.; Zhang, B.G.; Sun, X.B.; Liu, H.T.; Xiao, P.G. Traditional uses, phytochemistry, pharmacology and toxicology of Codonopsis: A review. J. Ethnopharmacol. 2018, 219, 50–70. [Google Scholar] [CrossRef]
- Commission, C.P. Pharmacopoeia of People’ s Republic of China; China Medical Science Press: Beijing, China, 2020. [Google Scholar]
- Zou, Y.F.; Zhang, Y.Y.; Paulsen, B.S.; Fu, Y.P.; Huang, C.; Feng, B.; Li, L.X.; Chen, X.F.; Jia, R.Y.; Song, X.; et al. Prospects of Codonopsis pilosula polysaccharides: Structural features and bioactivities diversity. Trends Food Sci. Technol. 2020, 103, 1–11. [Google Scholar] [CrossRef]
- Wakana, D.; Kawahara, N.; Goda, Y. Two New Pyrrolidine Alkaloids, Codonopsinol C and Codonopiloside A, Isolated from Codonopsis pilosula. Chem. Pharm. Bull. 2013, 61, 1315–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, F.Y.; Cheng, L.Z.; Yan, Y.M.; Liu, B.H.; Cheng, Y.X. Two Novel Proline-Containing Catechin Glucoside from Water-Soluble Extract of Codonopsis pilosula. Molecules 2018, 23, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.P.; Guo, Q.L.; Liu, Y.F.; Shi, J.G. Codonopiloneolignanin A, a polycyclic neolignan with a new carbon skeleton from the roots of Codonopsis pilosula. Chin. Chem. Lett. 2016, 27, 55–58. [Google Scholar] [CrossRef]
- Hu, X.Y.; Qin, F.Y.; Lu, X.F.; Zhang, L.S.; Cheng, Y.X. Three New Polyynes from Codonopsis pilosula and Their Activities on Lipid Metabolism. Molecules 2018, 23, 887. [Google Scholar] [CrossRef] [Green Version]
- Su, J.S.; Qin, F.Y.; Liu, Y.; Zhang, Y. Four new polyynes from Codonopsis pilosula collected in Yunnan province, China. Nat. Prod. Res. 2021, 35, 3548–3555. [Google Scholar] [CrossRef]
- Zheng, T.; Cheng, L.Z.; Yan, Y.M.; Liu, B.H.; Qin, F.Y.; Xu, F.R.; Cheng, Y.X. Two New Triterpenoids from the Roots of Codonopsis pilosula. Molecules 2018, 23, 383. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Wang, R.Y.; Zhao, Y.; Yu, Y.F.; Zhang, Z.X.; Hu, F.D.; Gao, K.; Fei, D.Q. Triterpenoids with alpha-glucosidase inhibitory activities from the roots of Codonopsis pilosula var. modesta. J. Chem. Res. 2021, 45, 635–638. [Google Scholar] [CrossRef]
- Cao, M.; Yu, C.; Yao, Z.; Gao, X.; Wu, S. Atractylodesin III maintains mitochondrial function and inhibits caspase-3 activity to reverse apoptosis of cardiomyocytes in AMI rats. Int. J. Clin. Exp. Pathol. 2019, 12, 198–204. [Google Scholar]
- Ming, I.; Chen, Y.; Yao, F.K.; Shi, J.T.; Yang, J.J.; Du, H.X.; Wang, X.Y.; Wang, Y.X.; Liu, J.G. Phosphorylated Codonopsis pilosula polysaccharide could inhibit the virulence of duck hepatitis A virus compared with Codonopsis pilosula polysaccharide. Int. J. Biol. Macromol. 2017, 94, 28–35. [Google Scholar] [CrossRef]
- Wang, Q.; Niu, L.L.; Liu, H.P.; Wu, Y.R.; Li, M.Y.; Jia, Q. Structural characterization of a novel polysaccharide from Pleurotus citrinopileatus and its antitumor activity on H22 tumor-bearing mice. Int. J. Biol. Macromol. 2021, 168, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.F.; Zhang, Y.Y.; Zhu, Z.K.; Fu, Y.P.; Paulsen, B.S.; Huang, C.; Feng, B.; Li, L.X.; Chen, X.F.; Jia, R.Y.; et al. Characterization of inulin-type fructans from two species of Radix Codonopsis and their oxidative defense activation and prebiotic activities. J. Sci. Food Agric. 2021, 101, 2491–2499. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.L.; Li, Y.X.; Cui, Y.S.; Jiang, S.L.; Dong, C.X.; Du, J. Structural characterization of three polysaccharides from the roots of Codonopsis pilosula and their immunomodulatory effects on RAW264.7 macrophages. Int. J. Biol. Macromol. 2019, 130, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-P.; Li, L.-X.; Zhang, B.-Z.; Paulsen, B.S.; Yin, Z.-Q.; Huang, C.; Feng, B.; Chen, X.-F.; Jia, R.-R.; Song, X.; et al. Characterization and prebiotic activity in vitro of inulin-type fructan from Codonopsis pilosula roots. Carbohydr. Polym. 2018, 193, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Xia, Y.; Luo, H.; Huang, S.; Wang, Y.; Shentu, Y.; Mahaman, Y.A.R.; Huang, F.; Ke, D.; Wang, Q.; et al. Codonopsis pilosula Polysaccharide Attenuates Tau Hyperphosphorylation and Cognitive Impairments in hTau Infected Mice. Front. Mol. Neurosci. 2018, 11, 437. [Google Scholar] [CrossRef]
- Wan, X.; Jin, X.; Wu, X.; Yang, X.; Lin, D.; Li, C.; Fu, Y.; Liu, Y.; Liu, X.; Lv, J. Structural characterisation and antitumor activity against non-small cell lung cancer of polysaccharides from Sanghuangporus vaninii. Carbohydr. Polym. 2022, 276, 118798. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, C.; Jing, L.; Feng, M.; Li, R.; Yang, Y. The structural characterization and immune modulation activitives comparison of Codonopsis pilosula polysaccharide (CPPS) and selenizing CPPS (sCPPS) on mouse in vitro and vivo. Int. J. Biol. Macromol. 2020, 160, 814–822. [Google Scholar] [CrossRef]
- Kuang, M.T.; Xu, J.Y.; Li, J.Y.; Yang, L.; Hou, B.; Zhao, Q.; Hu, J.M. Purification, structural characterization and immunomodulatory activities of a polysaccharide from the fruiting body of Morchella sextelata. Int. J. Biol. Macromol. 2022, 213, 394–403. [Google Scholar] [CrossRef]
- Buettner, R.; Schölmerich, J.; Bollheimer, L.C. High-fat diets: Modeling the metabolic disorders of human obesity in rodents. Obesity 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Bays, H.E.; Toth, P.P.; Kris-Etherton, P.M.; Abate, N.; Aronne, L.J.; Brown, W.V.; Gonzalez-Campoy, J.M.; Jones, S.R.; Kumar, R.; La Forge, R.; et al. Obesity, adiposity, and dyslipidemia: A consensus statement from the National Lipid Association. J. Clin. Lipidol. 2013, 7, 304–383. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.N.; Zhang, X.F. Polysaccharide extracted from Huperzia serrata using response surface methodology and its biological activity. Int. J. Biol. Macromol. 2020, 157, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, G. Extraction, characterization and antioxidant activities of pumpkin polysaccharide. Int. J. Biol. Macromol. 2018, 118, 770–774. [Google Scholar] [CrossRef]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Chen, F.; Zheng, F. Comparison of determination of sugar-PMP derivatives by two different stationary phases and two HPLC detectors: C-18 vs. amide columns and DAD vs. ELSD. J. Food Compos. Anal. 2021, 96, 103715. [Google Scholar] [CrossRef]
- Hakomori, S. A Rapid Permethylation of Glycolipid, and Polysaccharide Catalyzed by Methylsulfinyl Carbanion in Dimethyl Sulfoxide. J. Biochem. 1964, 55, 205–208. [Google Scholar]
- Liu, Y.; Ye, Y.; Hu, X.; Wang, J. Structural characterization and anti-inflammatory activity of a polysaccharide from the lignified okra. Carbohydr. Polym. 2021, 265, 118081. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Haboubi, H.; Haboubi, N. Adult obesity complications: Challenges and clinical impact. Ther. Adv. Endocrinol. 2020, 11. [Google Scholar] [CrossRef]
- Song, X.; Wang, L.; Liu, Y.; Zhang, X.; Weng, P.; Liu, L.; Zhang, R.; Wu, Z. The gut microbiota–brain axis: Role of the gut microbial metabolites of dietary food in obesity. Food Res. Int. 2022, 153, 110971. [Google Scholar] [CrossRef]
- Uchiyama, K.; Naito, Y.; Takagi, T. Intestinal microbiome as a novel therapeutic target for local and systemic inflammation. Pharmacol. Ther. 2019, 199, 164–172. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, H.; Zhang, K.; Lu, Y.; Wu, Q.; Chen, J.; Li, Y.; Wu, Q.; Chen, Y. Extraction, purification, structural characterization, and gut microbiota relationship of polysaccharides: A review. Int. J. Biol. Macromol. 2022, 213, 967–986. [Google Scholar] [CrossRef]
- Fang, Q.; Hu, J.; Nie, Q.; Nie, S. Effects of polysaccharides on glycometabolism based on gut microbiota alteration. Trends Food Sci. Technol. 2019, 92, 65–70. [Google Scholar] [CrossRef]
- Luan, F.; Ji, Y.; Peng, L.; Liu, Q.; Cao, H.; Yang, Y.; He, X.; Zeng, N. Extraction, purification, structural characteristics and biological properties of the polysaccharides from Codonopsis pilosula: A review. Carbohydr. Polym. 2021, 261, 117863. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time | Methylated | Type of Linkage | Major Mass Fragments (m/z) | Percentage(%) |
|---|---|---|---|---|
| 8.739 | 2,3,4,6-Me4-Glcp | T-Glcp | 43, 59, 102, 118, 129, 161, 205 | 20.53 |
| 9.856 | 2,4,6-Me3-Glcp | 1,3-Glcp | 43, 59, 87, 101, 118, 129, 161, 202, 234, 277 | 4.80 |
| 9.990 | 2,3,6-Me3-Galp | 1,4-Galp | 43, 59, 71, 87, 102, 118, 129, 162, 233 | 34.84 |
| 10.113 | 6-Me-Glcp | 1,2,3,4-Glcp | 87, 99, 115, 129, 157, 185, 218, 160, 333 | 10.36 |
| 10.698 | 2,3,4,6-Me4-Manp | T-Manp | 43, 59, 102, 118, 129, 161, 205 | 5.17 |
| 10.933 | 2,6-Me2-Glcp | 1,3,4-Glcp | 43, 59, 87, 118, 129, 160, 185, 305 | 15.25 |
| 12.308 | 2,3,5-Me3-Araf | T-Araf | 59, 71, 87, 102, 118, 129, 161, 162 | 9.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Q.; Huo, J.; Wang, Y.; Zhou, Y.; Luo, D.; Hou, J.; Zhang, Z.; Long, H.; Zhong, X.; Xie, C.; et al. The Obesity Amelioration Effect in High-Fat-Diet Fed Mice of a Homogeneous Polysaccharide from Codonopsis pilosula. Molecules 2022, 27, 5348. https://doi.org/10.3390/molecules27165348
Su Q, Huo J, Wang Y, Zhou Y, Luo D, Hou J, Zhang Z, Long H, Zhong X, Xie C, et al. The Obesity Amelioration Effect in High-Fat-Diet Fed Mice of a Homogeneous Polysaccharide from Codonopsis pilosula. Molecules. 2022; 27(16):5348. https://doi.org/10.3390/molecules27165348
Chicago/Turabian StyleSu, Qi, Jiangyan Huo, Yibin Wang, Yang Zhou, Dan Luo, Jinjun Hou, Zijia Zhang, Huali Long, Xianchun Zhong, Cen Xie, and et al. 2022. "The Obesity Amelioration Effect in High-Fat-Diet Fed Mice of a Homogeneous Polysaccharide from Codonopsis pilosula" Molecules 27, no. 16: 5348. https://doi.org/10.3390/molecules27165348