
Hepatoprotective Activity of Ethanol Extract of Rice Solid-State Fermentation of Ganoderma tsugae against CCl4-Induced Acute Liver Injury in Mice

Abstract
:1. Introduction
2. Results
2.1. Screening of Optimal G. tsugae Strain with High Triterpenoid Content
2.2. Optimization of Culture Conditions of G42
2.2.1. Effects of Single-Factor on Triterpenoid Content of G42
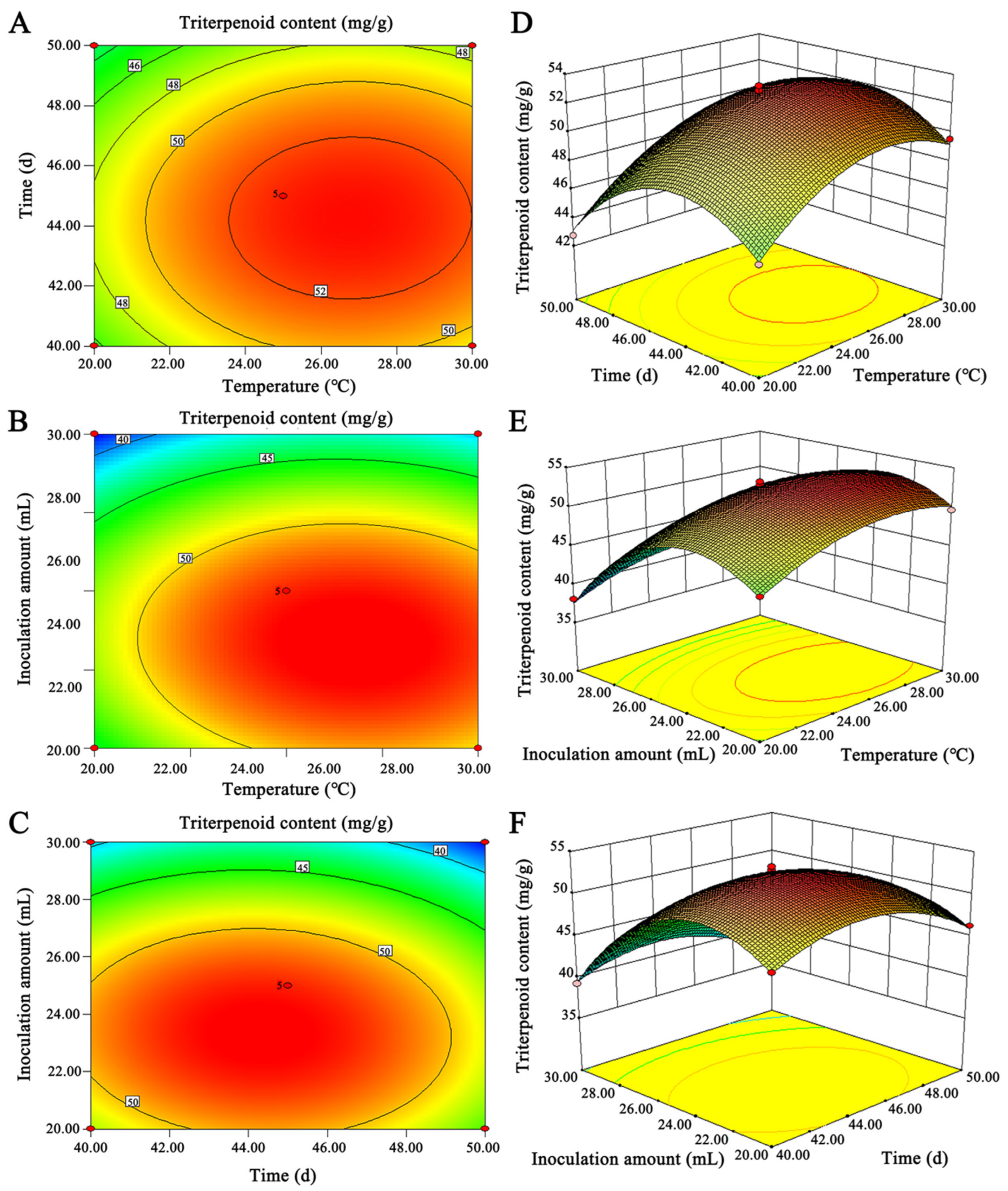
2.2.2. Optimization of Culture Conditions of G42 by Response Surface Methodology
2.3. Evaluation of the Hepatoprotective Activity of the Ethanol Extract
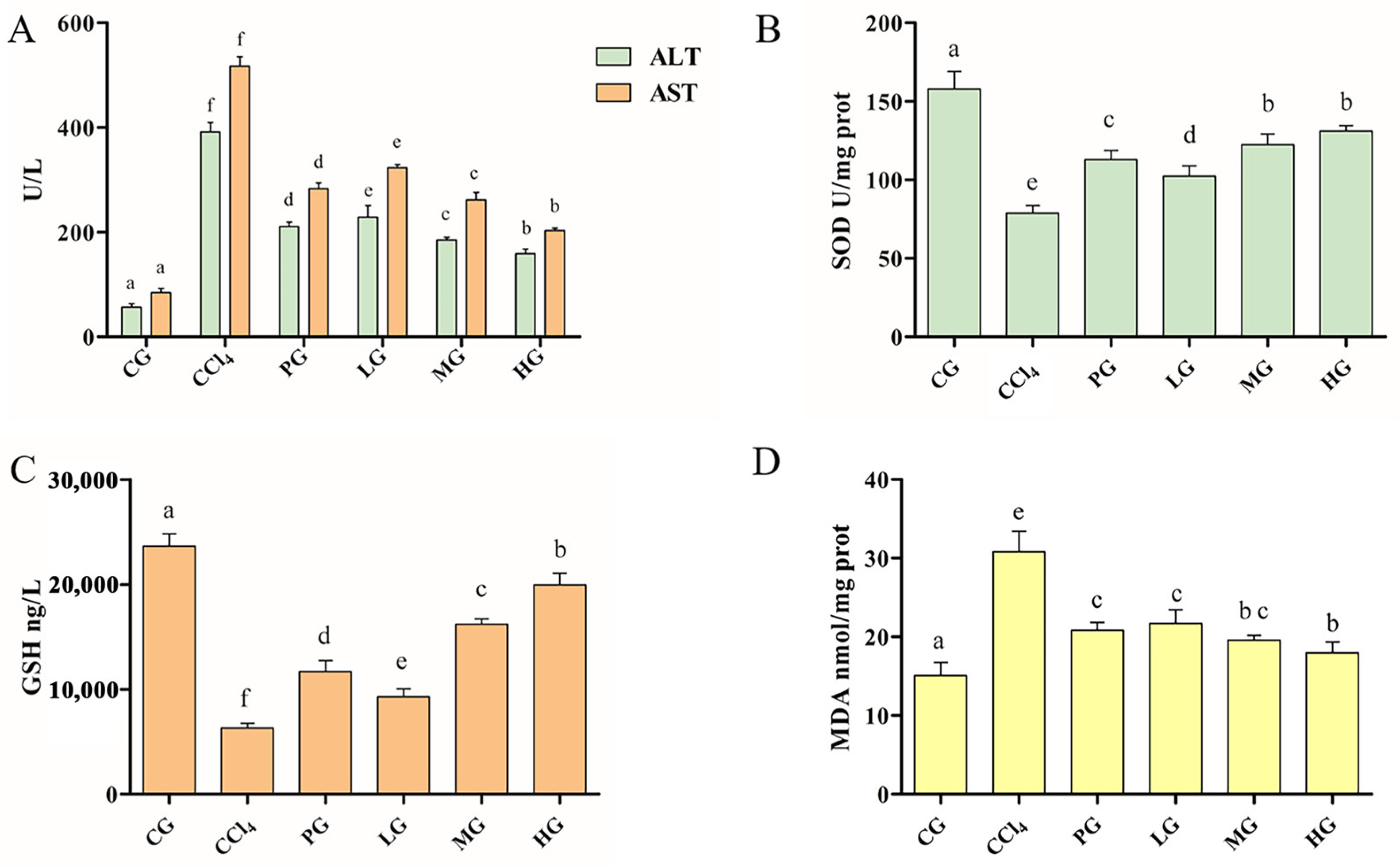
2.3.1. Effects of the Ethanol Extract on Serum ALT and AST Activities
2.3.2. Effect of the Ethanol Extract on the Antioxidant Enzyme Activity of Liver Tissue
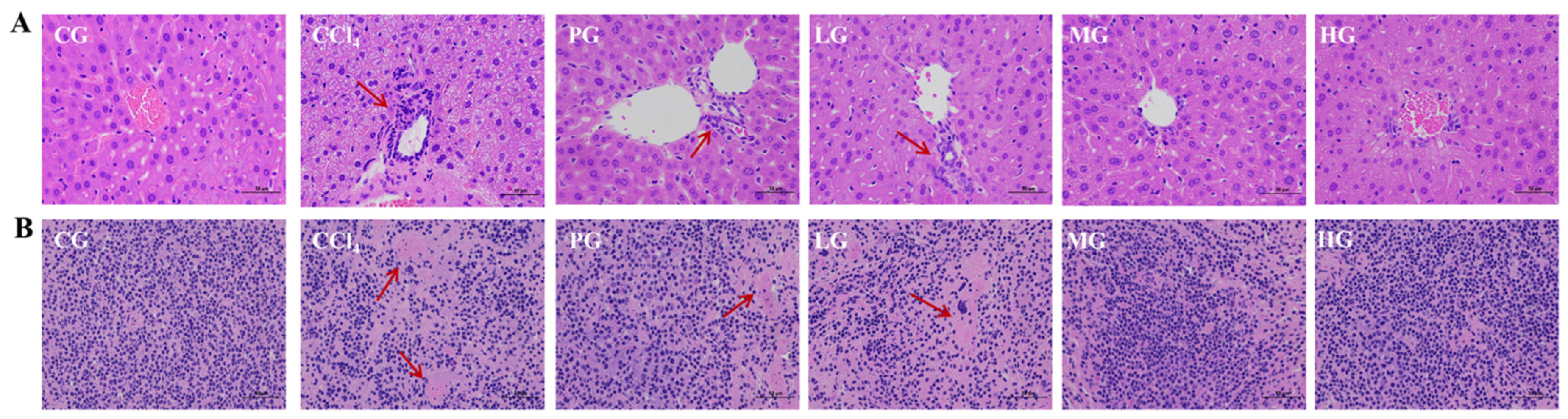
2.3.3. Histopathological Analysis of Liver and Kidney
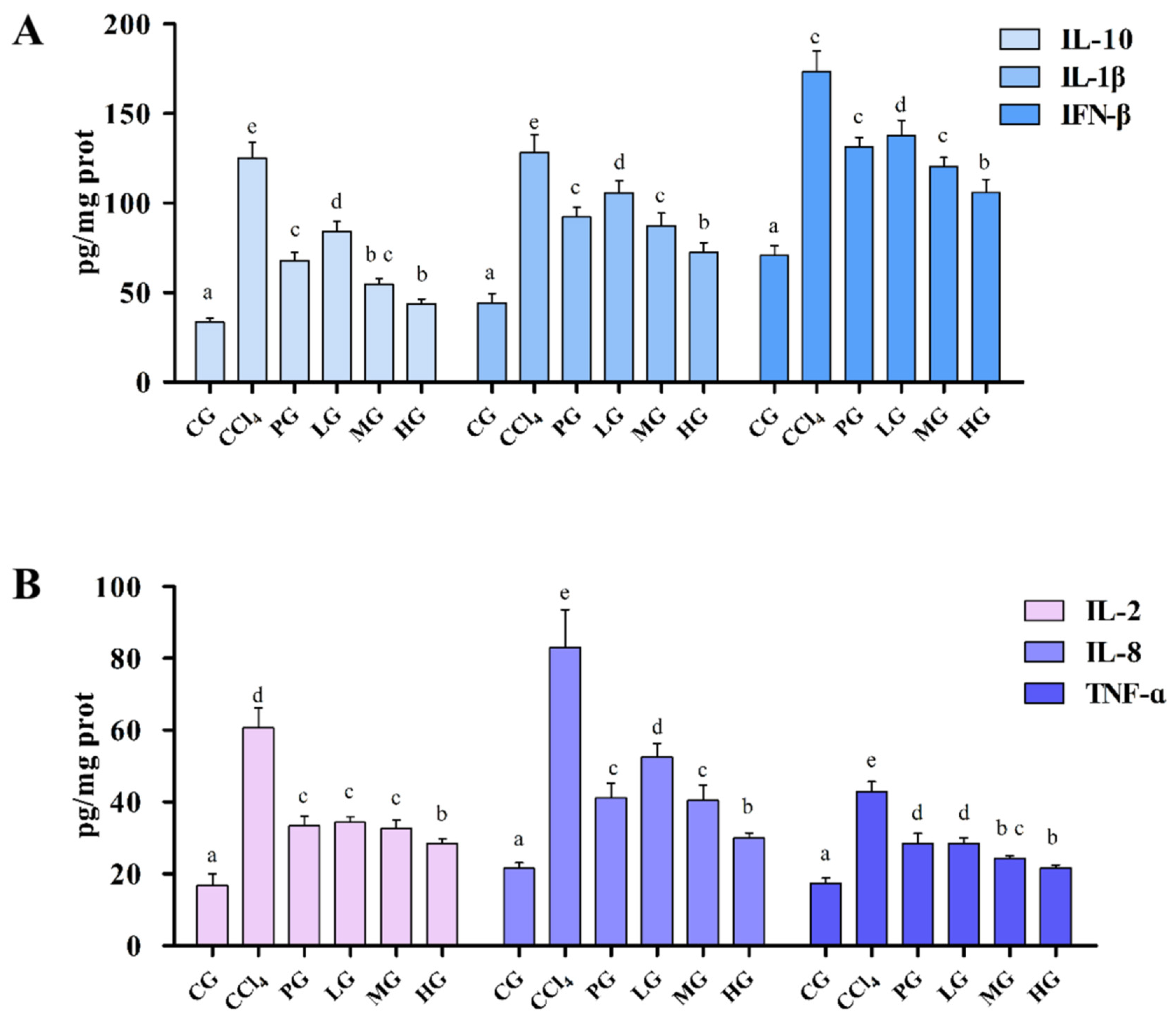
2.3.4. Effects of the Ethanol Extract on Inflammatory Factors of Liver Tissue
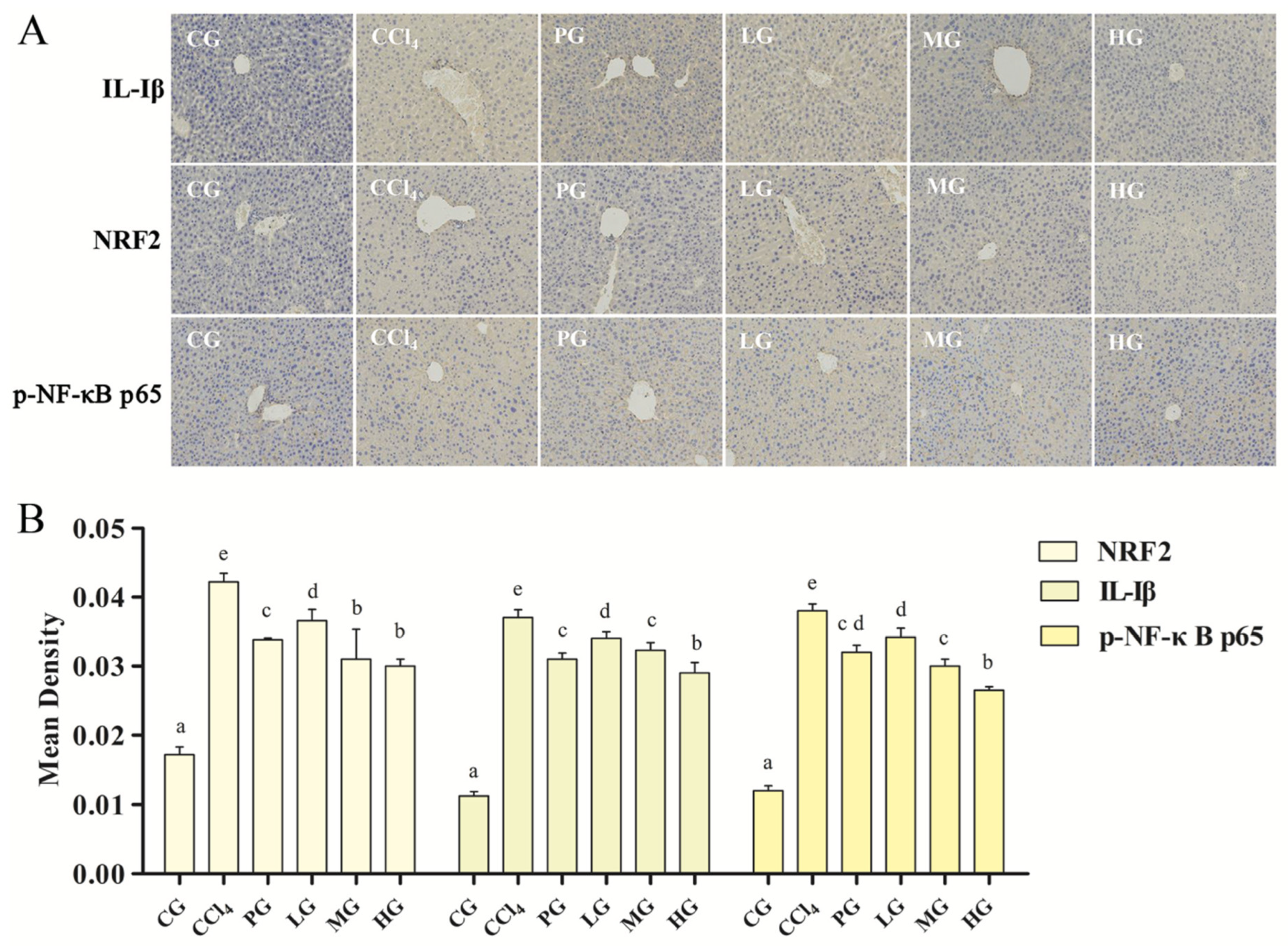
2.3.5. Effect of the Ethanol Extract on the IHC of Liver Tissue
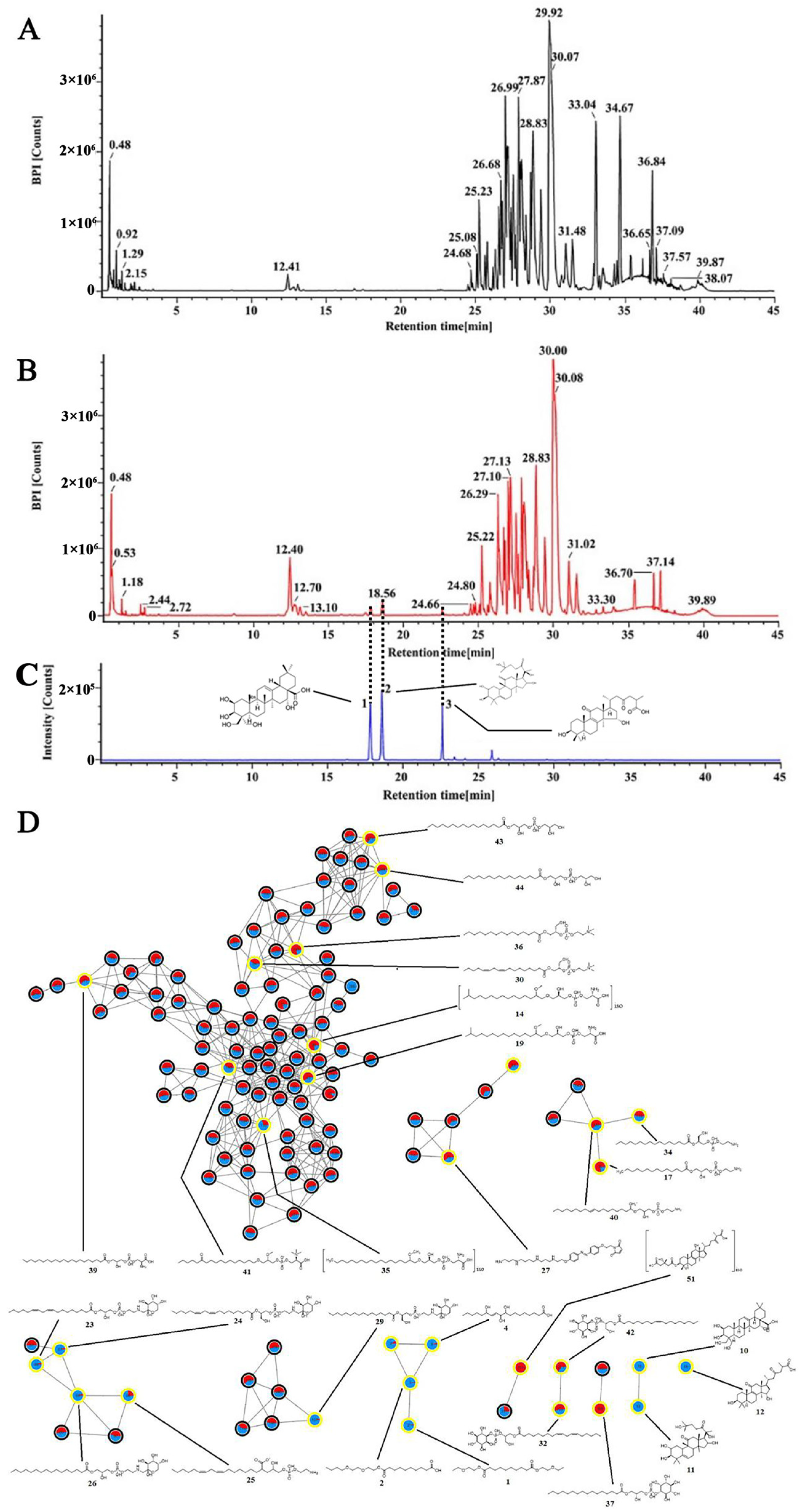
2.4. Chemical Identification of the Ethanol Extract of G42
3. Discussion
4. Materials and Methods
4.1. Strains and Reagents
4.2. Screening of High-Yield Triterpenoids Strains of G. tsugae from Rice by SSF
4.2.1. SSF Culture of G. tsugae Strains
4.2.2. Triterpenoid Content in Fermentation Products
4.2.3. Antioxidant Activity In Vitro of Ethanol Extract of Fermentation Products
4.3. Optimization Conditions of G42 with High Triterpenoid Content by Response Surface Methodology
4.3.1. Effects of Culture Conditions on Triterpenoid Content
4.3.2. Response Surface Design of G42 from Rice by SSF
4.4. Evaluation of the Hepatoprotective Activity of Ethanol Extract of G42
4.4.1. Preparation of G42 Extracts
4.4.2. Experimental Design of Animals
4.4.3. Determination of Serum ALT and AST Levels
4.4.4. Inflammatory Cytokines and Antioxidant Activity of Liver Tissue
4.4.5. Histopathological Analysis
4.4.6. Immunohistochemistry (IHC)
4.5. Chemical Identification of the Ethanol Extract of G42 by Mass Spectrometry
4.5.1. Liquid Chromatography and Mass Spectrometry (LC-MS)
4.5.2. Data Processing and Compound Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Losser, M.R.; Payen, D. Mechanisms of liver damage. Semin. Liver Dis. 1996, 16, 357. [Google Scholar]
- Chiu, H.F.; Fu, H.Y.; Lu, Y.Y.; Han, Y.C.; Shen, Y.C.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C.K. Triterpenoids and polysaccharide peptides-enriched Ganoderma lucidum: A randomized, double-blind placebo-controlled crossover study of its antioxidation and hepatoprotective efficacy in healthy volunteers. Pharm. Biol. 2017, 55, 1041–1046. [Google Scholar] [CrossRef] [Green Version]
- Sudheesh, N.P.; Ajith, T.A.; Mathew, J.; Nima, N.; Janardhanan, K.K. Ganoderma lucidum protects liver mitochondrial oxidative stress and improves the activity of electron transport chain in carbon tetrachloride intoxicated rats. Hepatol. Res. 2012, 42, 181–191. [Google Scholar]
- Zhao, C.; Fan, J.L.; Liu, Y.Y.; Guo, W.L.; Cao, H.; Xiao, J.; Wang, Y.; Liu, B. Hepatoprotective activity of Ganoderma lucidum triterpenoids in alcohol-induced liver injury in mice, an iTRAQ-based proteomic analysis. Food Chem. 2019, 15, 148–156. [Google Scholar]
- Wu, Y.W.; Chen, K.D.; Lin, W.C. Effect of Ganoderma tsugae on chronically carbon tetrachloride-intoxicated rats. Am. J. Chin. Med. 2004, 32, 841–850. [Google Scholar]
- Zhou, X.; Lin, J.; Yin, Y.; Zhao, J.; Sun, X.; Tang, K. Ganodermataceae: Natural products and their related pharmacological functions. Am. J. Chin. Med. 2007, 35, 559–574. [Google Scholar] [CrossRef]
- Li, P. Extractions; Isolations; Purification and Bioactivities of Polysaccharides from Ganoderma tsugae. Ph.D. Thesis, Jilin University, Changchun, China, 2016. [Google Scholar]
- Lin, K.W.; Maitraie, D.; Huang, A.M.; Wang, J.P.; Lin, C.N. Triterpenoids and an alkamide from Ganoderma tsugae. Fitoterapia 2016, 108, 73–80. [Google Scholar] [CrossRef]
- Su, H.J.; Fann, Y.F.; Chung, M.I.; Won, S.J.; Lin, C.N. New lanostanoids of Ganoderma tsugae. J. Nat. Prod. 2000, 63, 514–516. [Google Scholar]
- Lin, K.W.; Chen, Y.T.; Yang, S.C.; Wei, B.L.; Hung, C.F.; Lin, C.N. Xanthine oxidase inhibitory lanostanoids from Ganoderma tsugae. Fitoterapia 2013, 89, 231–238. [Google Scholar]
- Chien, R.C.; Yen, M.T.; Tseng, Y.H.; Mau, J.L. Chemical characteristics and anti-proliferation activities of Ganoderma tsugae polysaccharides. Carbohydr. Polym. 2015, 5, 90–98. [Google Scholar] [CrossRef]
- You, B.J.; Lee, H.Z.; Chung, K.R.; Lee, M.H.; Huang, M.J.; Tien, N.; Chan, C.W.; Kuo, Y.H. Enhanced production of ganoderic acids and cytotoxicity of Ganoderma lucidum using solid-medium culture. Biosci. Biotechnol. Biochem. 2012, 76, 1529–1534. [Google Scholar] [CrossRef]
- Yang, L.C.; Fu, T.J.; Yang, F.C. Biovalorization of soybean residue (okara) via fermentation with Ganoderma lucidum and Lentinus edodes to attain products with high anti-osteoporotic effects. J. Biosci. Bioeng. 2020, 129, 514–518. [Google Scholar] [CrossRef]
- Costa, T.M.; Kaufmann, V.; Paganelli, C.J.; Siebert, D.A.; Micke, G.A.; Alberton, M.D.; Tavares, L.B.B.; de Oliveira, D. Kinetic identification of phenolic compounds and potential production of caffeic acid by Ganoderma lipsiense in solid-state fermentation. Bioprocess. Biosyst. Eng. 2019, 8, 1325–1332. [Google Scholar] [CrossRef]
- Costa, T.M.; Lenzi, J.; Paganelli, C.J.; Filho, H.H.D.S.; Alberton, M.D.; Tavares, L.B.B.; de Oliveira, D. Liposoluble compounds from Ganoderma lipsiense grown on solid red rice medium with antiparasitic and antibacterial properties. Biotechnol. Appl. Biochem. 2020, 67, 180–185. [Google Scholar] [CrossRef]
- Wang, Y.; Li, B.; Liu, D.; Yang, X.; Li, S.; Hao, G.; Wen, H.; Liu, H. Two new sesquiterpenes and six norsesquiterpenes from the solid culture of the edible mushroom Flammulina velutipes. Tetrahedron 2012, 68, 3012–3018. [Google Scholar] [CrossRef]
- Perez De Souza, L.; Alseekh, S.; Brotman, Y.; Fernie, A.R. Network-based strategies in metabolomics data analysis and interpretation: From molecular networking to biological interpretation. Expert. Rev. Proteom. 2020, 17, 243–255. [Google Scholar] [CrossRef]
- Kuloglu, N.; Sönmez, M.F. A biochemical and immunohistochemical study of the protective effects of carnosine for carbon tetrachloride induced liver injury in rats. Biotech. Histochem. 2015, 90, 608–614. [Google Scholar] [CrossRef]
- Go, J.; Kim, J.E.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, Y.H.; Lim, Y.; Hong, J.T.; Hwang, D.Y. Protective effect of gallotannin-enriched extract isolated from galla rhois against CCl4-induced hepatotoxicity in ICR Mice. Nutrients 2016, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Palliyaguru, D.L.; Chartoumpekis, D.V.; Wakabayashi, N.; Skoko, J.J.; Yagishita, Y.; Singh, S.V.; Kensler, T.W. Withaferin A induces Nrf2-dependent protection against liver injury: Role of Keap1-independent mechanisms. Free Radic. Biol. Med. 2016, 101, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.M.; Wang, Y.; Tan, H.S.; Yu, T.; Fan, X.M.; Chen, P.; Zeng, H.; Huang, M.; Bi, H.C. Schisandrol B protects against acetaminophen-induced acute hepatotoxicity in mice via activation of the NRF2/ARE signaling pathway. Acta Pharmacol. Sin. 2016, 37, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Li, H.; Liu, Q.; Liu, F.; Tang, L.; Li, C.; Yuan, Y.; Zhan, Y.; Xu, W.; Li, W.; et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell Signal. 2011, 23, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bao, L.; Yang, X.; Li, L.; Li, S.; Gao, H.; Yao, X.S.; Wen, H.; Liu, H.W. Bioactive sesquiterpenoids from the solid culture of the edible mushroom Flammulina velutipes growing on cooked rice. Food Chem. 2012, 132, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Cheng, M.J.; Wu, M.D.; Chen, J.J.; Chen, Y.L.; Chang, H.S.; Chen, K.P. Secondary metabolites from the fermented rice of the fungus Monascus purpureus and their bioactivities. Nat. Prod. Res. 2019, 33, 3541–3550. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.S.; Asatiani, M.D.; Sharvit, L.E.; Trabelcy, B.; Barseghyan, G.S.; Wasser, S.P. Chemical Composition and Medicinal Value of the New Ganoderma tsugae var. jannieae CBS-120304 Medicinal Higher Basidiomycete Mushroom. Int. J. Med. Mushrooms 2015, 17, 735–747. [Google Scholar] [CrossRef]
- Tan, Y.; Li, Y.; Zhou, F.; Guo, J.; Wang, T.; Shi, Y.; Yang, Y.; Lu, J.; Pei, G. Administration of a mixture of triterpenoids from yeyachun and phenolic acids from danshen ameliorates carbon tetrachloride-induced liver fibrosis in mice by the regulation of intestinal flora. J. Pharmacol. Sci. 2020, 143, 165–175. [Google Scholar] [CrossRef]
- Buffler, M.; Becker, C.; Windisch, W.; Schümann, K. Inflammation neither increases hepatic hepcidin nor affects intestinal (59)Fe-absorption in two murine models of bowel inflammation, hemizygous TNF(ΔARE/+) and homozygous IL-10(−/−) mice. J. Trace. Elem. Med. Biol. 2015, 32, 162–167. [Google Scholar] [CrossRef]
- Zhao, Z.; Gong, S.; Wang, S.; Ma, C. Effect and mechanism of evodiamine against ethanol-induced gastric ulcer in mice by suppressing Rho/NF-кB pathway. Int. Immunopharmacol. 2015, 28, 588–595. [Google Scholar] [CrossRef]
- Jiang, T.; Tian, F.; Zheng, H.; Whitman, S.A.; Lin, Y.; Zhang, Z.; Zhang, N.; Zhang, D.D. Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-κB-mediated inflammatory response. Kidney Int. 2014, 85, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Yuan, F.; Li, H.; Feng, Y.; Zhang, Y.; Zhang, C.; Zhang, J.; Song, Z.; Jia, L. The ameliorations of Ganoderma applanatum residue polysaccharides against CCl4 induced liver injury. Int. J. Biol. Macromol. 2019, 137, 1130–1140. [Google Scholar] [CrossRef]
- Chen, Y.S.; Chen, Q.Z.; Wang, Z.J.; Hua, C. Anti-Inflammatory and Hepatoprotective Effects of Ganoderma lucidum polysaccharides against Carbon Tetrachloride-Induced liver Injury in Kunming Mice. Pharmacology 2019, 103, 143–150. [Google Scholar] [CrossRef]
- Hu, Z.; Du, R.; Xiu, L.; Bian, Z.; Ma, C.; Sato, N.; Hattori, M.; Zhang, H.; Liang, Y.; Yu, S.; et al. Protective effect of triterpenes of Ganoderma lucidum on lipopolysaccharide-induced inflammatory responses and acute liver injury. Cytokine 2020, 127, 154917. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Kang, J.; Wu, C.H.; Li, Y.; Chen, R.Y. Triterpenoids from Ganoderma theaecolum. China J. Chin. Mater. Med. 2016, 41, 1075–1080. [Google Scholar]
- Che, X.Q.; Li, S.P.; Zhao, J. Ganoderma triterpenoids from aqueous extract of Ganoderma lucidum. China J. Chin. Mater. Med. 2017, 42, 1908–1915. [Google Scholar]
- Yang, Z.; Liu, B.; Yang, L.E.; Zhang, C. Platycodigenin as potential drug candidate for Alzheimer’s Disease via modulating microglial polarization and neurite Regeneration. Molecules 2019, 24, 3207. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, T.; Ren, L.; Jing, S.; Li, Z.; Zuo, P.; Li, T.; Wang, Y.; Zhang, J.; Wei, Z. Cucurbitacin IIb induces apoptosis and cell cycle arrest through regulating EGFR/MAPK pathway. Environ. Toxicol. Pharmacol. 2021, 81, 103542. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Huang, Q. Revised method for determining Ganoderma lingzhi terpenoids by UV-Vis spectrophotometry based on colorimetric vanillin perchloric acid reaction. Mycosystema 2018, 37, 1792–1801. [Google Scholar]
- Zhang, G.L.; Shen, L.; Hang, B.Q.; Lai, Y.H.; Zhou, M.; Zou, J.M. Main Active Components and Antioxidant Activity of Extracts from Ganoderma lucidum Solid Fermentation Longan Seed. Sci. Technol. Food Ind. 2019, 40, 53–57. [Google Scholar]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 9, 828–837. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Biomass (g) | Triterpenoid Content (mg/g) | Total Reducing Power | DPPH Clearance Rate (%) |
|---|---|---|---|---|
| G10 | 166.11 ± 1.23 | 31.79 ± 1.25 | 0.91 | 72.91 |
| G12 | 169.54 ± 2.21 | 30.17 ± 2.03 | 0.28 | 45.89 |
| G17 | 169.37 ± 1.57 | 19.85 ± 1.74 | 0.77 | 84.89 |
| G20 | 171.2 ± 3.45 | 19.43 ± 1.65 | 0.40 | 54.75 |
| G27 | 171.89 ± 2.36 | 35.03 ± 1.76 | 0.85 | 65.48 |
| G40 | 174.86 ± 4.35 | 24.70 ± 2.14 | 0.55 | 65.11 |
| G41 | 167.38 ± 3.66 | 34.15 ± 0.96 | 1.08 | 83.13 |
| G42 | 187.78 ± 2.96 | 48.94 ± 2.26 | 1.24 | 90.96 |
| G43 | 188.76 ± 2.39 | 27.94 ± 1.35 | 0.63 | 67.45 |
| G78 | 177.69 ± 2.47 | 43.11 ± 1.39 | 0.79 | 79.02 |
| Sum of Squares | df | Mean Square | F Value | p-Value Prob > F | Sig. |
|---|---|---|---|---|---|
| 490.44 | 9 | 54.49 | 196.39 | <0.0001 | ** |
| 28.39 | 1 | 28.39 | 102.31 | <0.0001 | ** |
| 10.51 | 1 | 10.51 | 37.88 | 0.0005 | * |
| 154.88 | 1 | 154.88 | 558.18 | <0.0001 | ** |
| 0.060 | 1 | 0.060 | 0.22 | 0.6560 | |
| 0.64 | 1 | 0.64 | 2.31 | 0.1726 | |
| 0.56 | 1 | 0.56 | 2.03 | 0.1975 | |
| 29.24 | 1 | 29.24 | 105.38 | <0.0001 | ** |
| 58.90 | 1 | 58.90 | 212.28 | <0.0001 | ** |
| 181.35 | 1 | 181.35 | 653.56 | <0.0001 | ** |
| 1.94 | 7 | 0.28 | |||
| 1.03 | 3 | 0.34 | 1.49 | 0.3446 | - |
| 0.92 | 4 | 0.23 | |||
| 492.39 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Lv, W.; Fu, Y.; Li, Y.; Wang, J.; Chen, D.; Han, X.; Li, Z. Hepatoprotective Activity of Ethanol Extract of Rice Solid-State Fermentation of Ganoderma tsugae against CCl4-Induced Acute Liver Injury in Mice. Molecules 2022, 27, 5347. https://doi.org/10.3390/molecules27165347
Zhang X, Lv W, Fu Y, Li Y, Wang J, Chen D, Han X, Li Z. Hepatoprotective Activity of Ethanol Extract of Rice Solid-State Fermentation of Ganoderma tsugae against CCl4-Induced Acute Liver Injury in Mice. Molecules. 2022; 27(16):5347. https://doi.org/10.3390/molecules27165347
Chicago/Turabian StyleZhang, Xin, Wentao Lv, Yongping Fu, Yu Li, Jinhe Wang, Dongjie Chen, Xuerong Han, and Zhenhao Li. 2022. "Hepatoprotective Activity of Ethanol Extract of Rice Solid-State Fermentation of Ganoderma tsugae against CCl4-Induced Acute Liver Injury in Mice" Molecules 27, no. 16: 5347. https://doi.org/10.3390/molecules27165347