
Exceptional Properties of Lepidium sativum L. Extract and Its Impact on Cell Viability, Ros Production, Steroidogenesis, and Intracellular Communication in Mice Leydig Cells In Vitro

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material Collection and Processing
2.2. Biochemical Analysis of the Extract
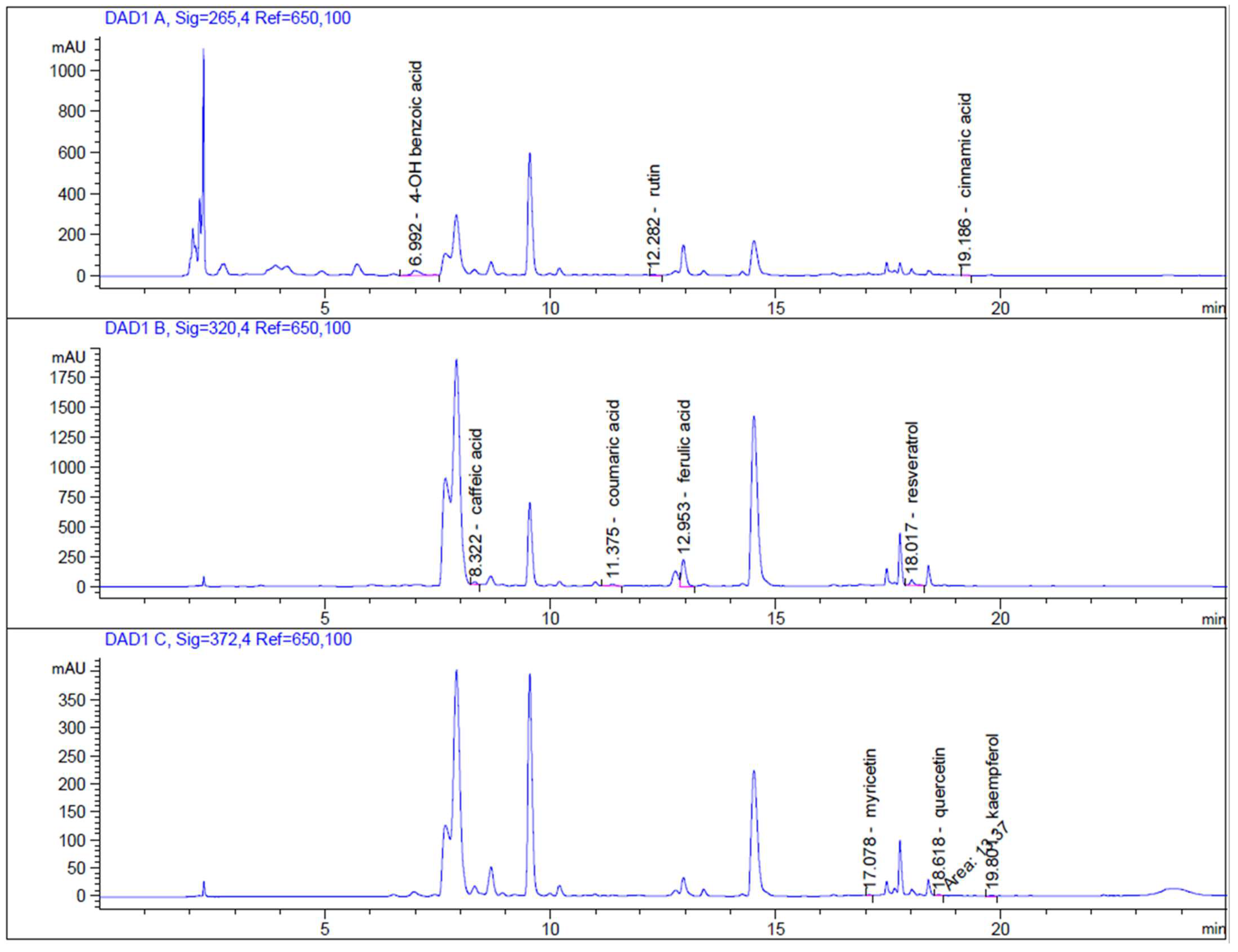
2.3. High-Performance Liquid Chromatography (HPLC-DAD) Analysis
2.4. Total Antioxidant Capacity
2.5. Leydig TM3 Cell Culture and Experimental Setups In Vitro
2.6. Cytotoxic Assays
2.6.1. Cell Viability
2.6.2. Cell Membrane Integrity
2.6.3. Lysosomal Activity
2.7. ROS Production
2.8. Steroid Hormone Secretion
2.9. Gap Junctional Intracellular Communication Assay
2.10. Statistical Analyses
3. Results
3.1. Biochemical Profile of Lepidium sativum L.
3.2. Biologically Active Compounds and Antioxidant Capacity of Lepidium sativum L.
3.3. Assessment of the Cytotoxic Effect of Lepidium sativum L.
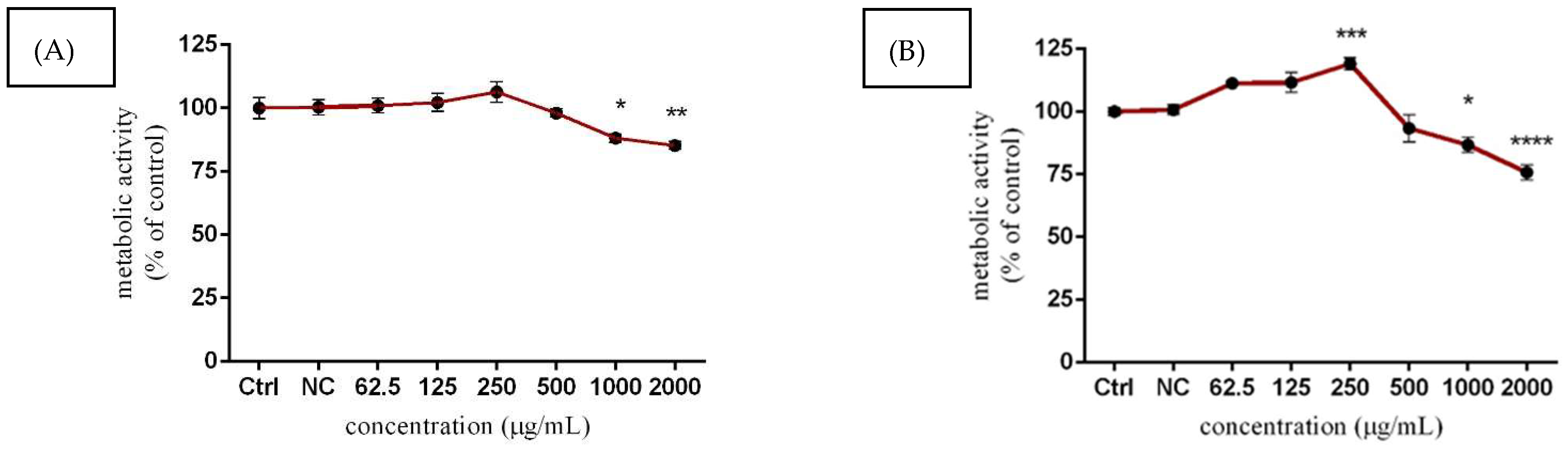
3.3.1. TM3 Metabolic Activity
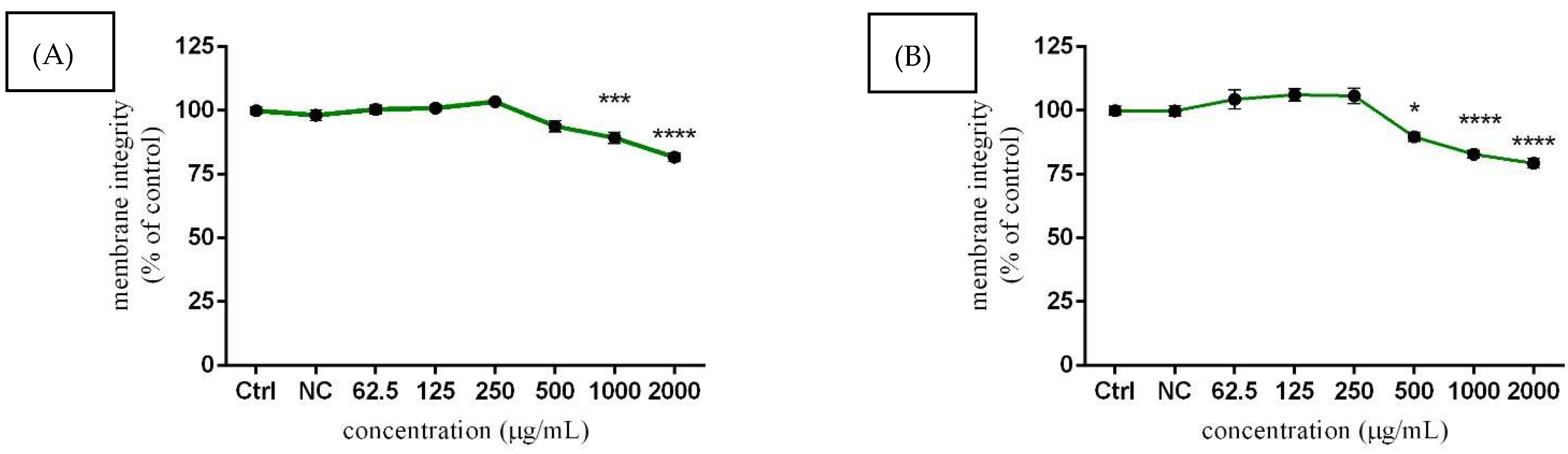
3.3.2. TM3 Cell Membrane Integrity
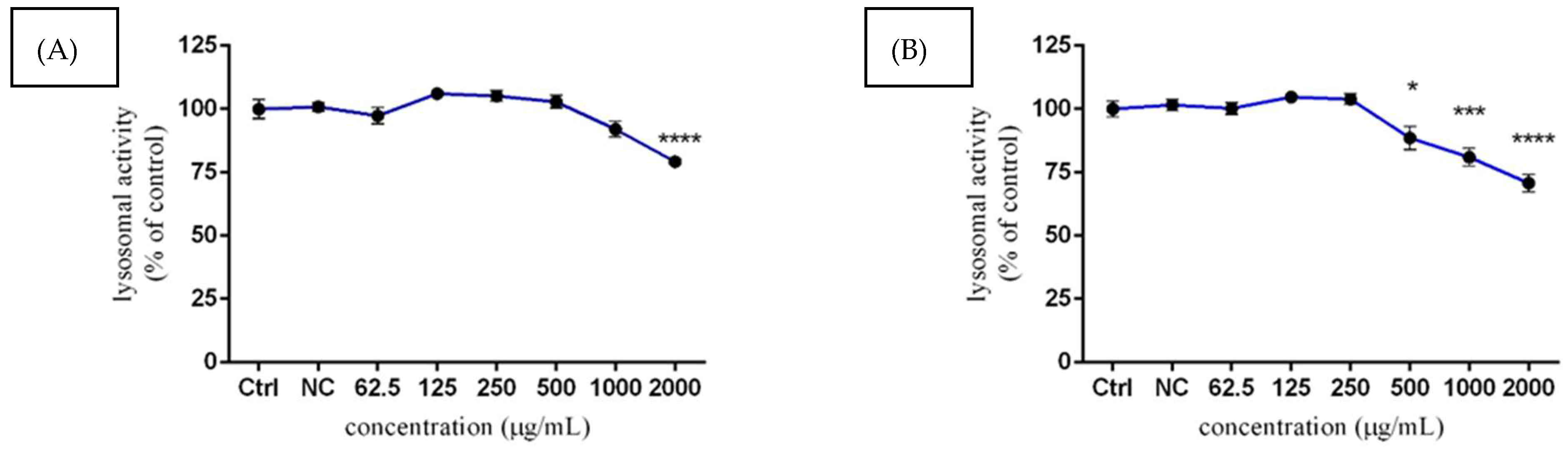
3.3.3. Lysosomal Activity of TM3 Cells
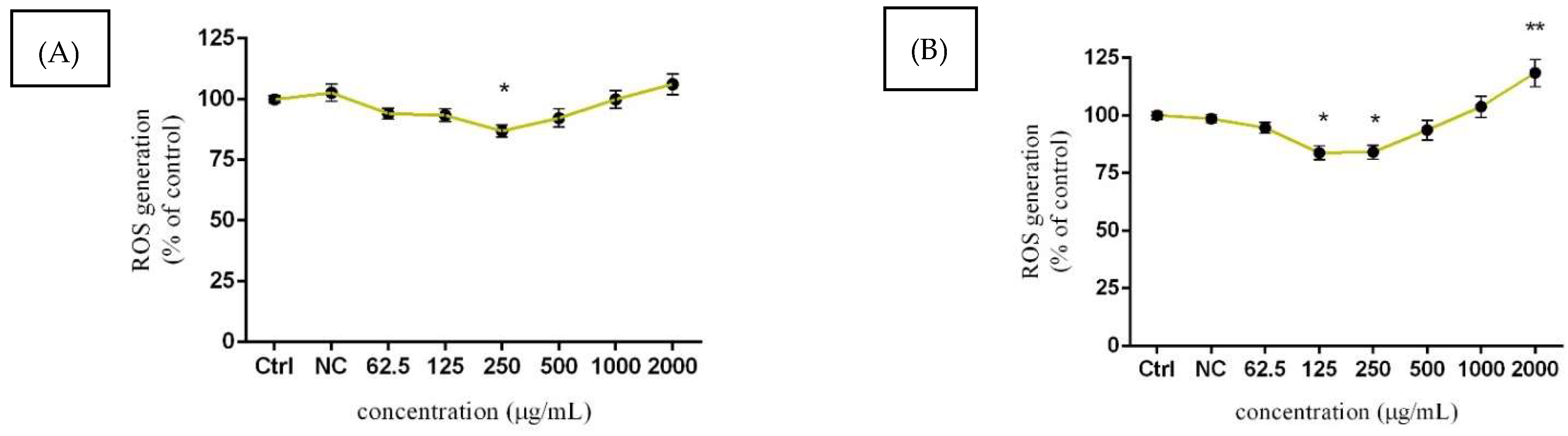
3.4. Assessment of ROS Production
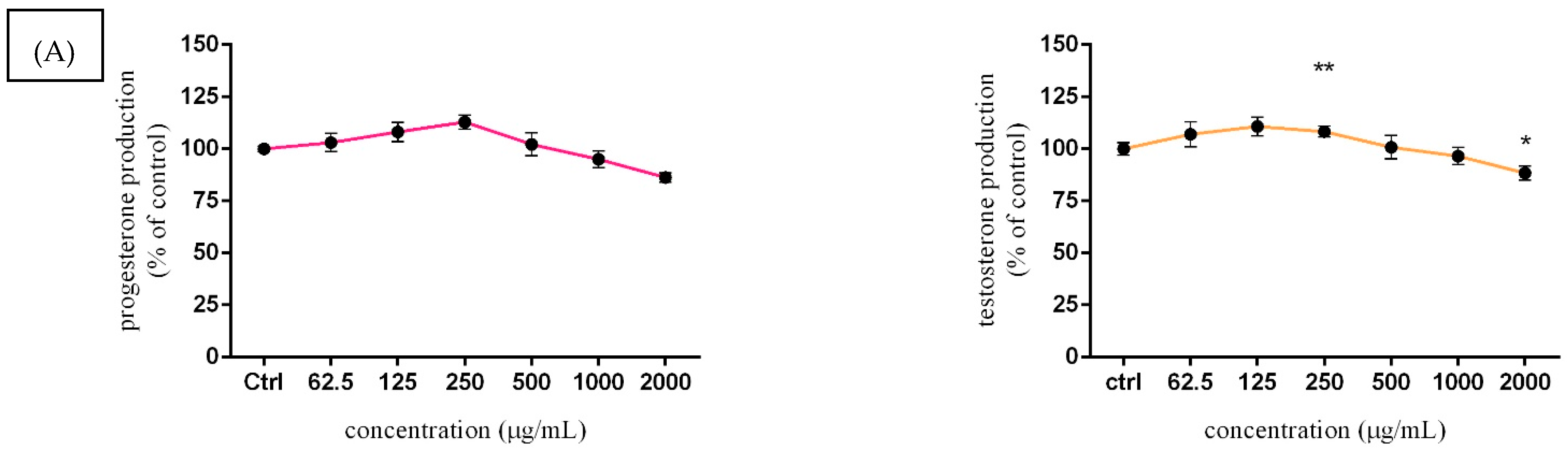
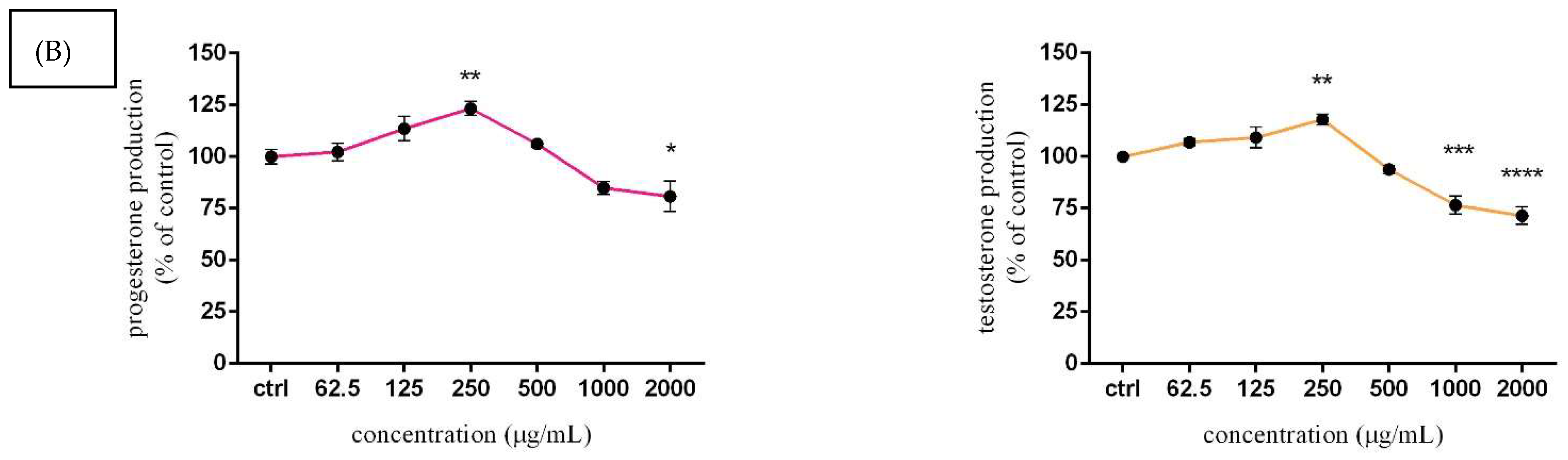
3.5. Assessment of Steroid Hormone Secretion
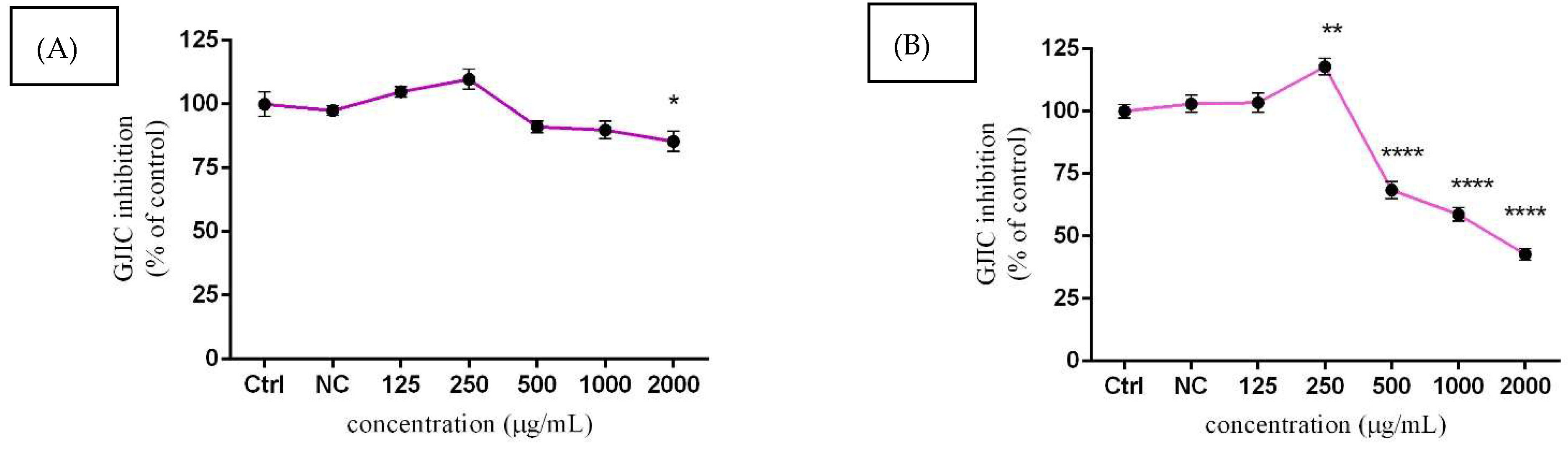
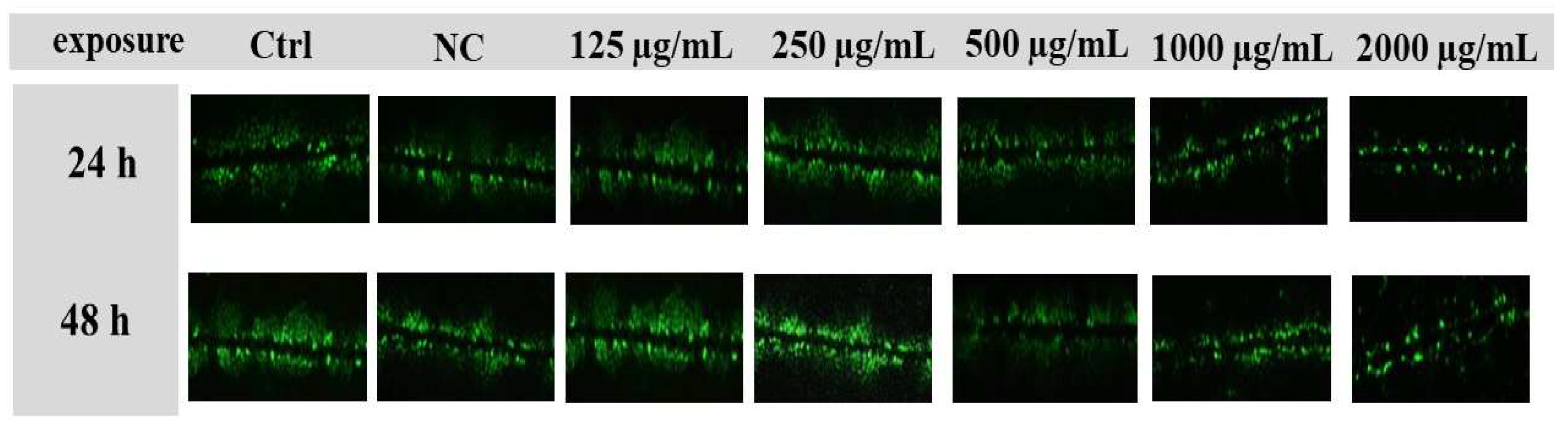
3.6. Assessment of GJIC Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, H.; Gong, T.T.; Jiang, Y.T.; Zhang, S.; Zhao, Y.H.; Wu, Q.J. Global, regional, and national prevalence and disability-adjusted life-years for infertility in 195 countries and territories, 1990–2017: Results from a global burden of disease study, 2017. Aging 2019, 11, 10952–10991. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Steiner, A.Z. Age-related infertility. Obstet. Gynecol. Clin. 2015, 42, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Belloc, S.; Hazout, A.; Zini, A.; Merviel, P.; Cabry, R.; Chahine, H.; Copin, H.; Benkhalifa, M. How to overcome male infertility after 40: Influence of paternal age on fertility. Maturitas 2014, 78, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo, M.; Mendiola, J.; Koch, H.M.; Moñino-García, M.; Noguera-Velasco, J.A.; Torres-Cantero, A.M. Associations between urinary organophosphate pesticide metabolite levels and reproductive parameters in men from an infertility clinic. Env. Res. 2015, 137, 292–298. [Google Scholar] [CrossRef]
- Akasheh, G.; Sirati, L.; Noshad Kamran, A.R.; Sepehrmanesh, Z. Comparison of the effect of sertraline with behavioral therapy on semen parameters in men with primary premature ejaculation. Urology 2014, 83, 800–804. [Google Scholar] [CrossRef]
- Gautam, R.; Priyadarshini, E.; Nirala, J.; Rajamani, P. Impact of nonionizing electromagnetic radiation on male infertility: An assessment of the mechanism and consequences. Int. J. Radiat. Biol. 2022, 98, 1063–1073. [Google Scholar] [CrossRef]
- Maresch, C.C.; Stute, D.C.; Alves, M.G.; Oliveira, P.F.; de Kretser, D.M.; Linn, T. Diabetes-induced hyperglycemia impairs male reproductive function: A systematic review. Hum. Reprod. Update 2018, 24, 86–105. [Google Scholar] [CrossRef]
- Khizroeva, J.; Nalli, C.; Bitsadze, V.; Lojacono, A.; Zatti, S.; Andreoli, L.; Tincani, A.; Shoenfeld, Y.; Makatsariya, A. Infertility in women with systemic autoimmune diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101369. [Google Scholar] [CrossRef]
- Rahman, R.; Saeed, M.; Zia, S.; Jan, F.A.U.Z.I.A.; Muzaffar, S.; Waheed, A. Thyroid disorders; Prevalence of thyroid disorders in primaty infertile women of reproductive age. Prof. Med. J. 2019, 26, 101–108. [Google Scholar] [CrossRef]
- Santana, V.P.; James, E.R.; Miranda-Furtado, C.L.; Souza, M.F.; Pompeu, C.P.; Esteves, S.C.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Reis, R.M.D. Differential DNA methylation pattern and sperm quality in men with varicocele. Fertil. Steril. 2020, 114, 770–778. [Google Scholar] [CrossRef]
- Benatta, M.; Kettache, R.; Buchholz, N.; Trinchieri, A. The impact of nutrition and lifestyle on male fertility. Arch. Ital. Urol. Androl. 2020, 92, 121–131. [Google Scholar] [CrossRef]
- Leaver, R.B. Male infertility: An overview of causes and treatment options. B. J. Nurs. 2016, 25, S35–S40. [Google Scholar] [CrossRef]
- Martins, R.V.; Silva, A.; Duarte, A.P.; Socorro, S.; Correia, S.; Maia, C.J. Natural Products as Protective Agents for Male Fertility. BioChem 2021, 1, 11. [Google Scholar] [CrossRef]
- Painuli, S.; Quispe, C.; Herrera-Bravo, J.; Semwal, P.; Martorell, M.; Almarhoon, Z.M.; Taheri, Y.; Calina, D.; Cho, W.C. Nutraceutical profiling, bioactive composition, and biological applications of Lepidium sativum L. Oxid Med. Cell Longev. 2021, 2022, 2910411. [Google Scholar] [CrossRef]
- Ibraheem, S.R.; Ibraheem, M.R.; Hashim, S.S. Effect of Lepidium sativum aqueous crude extract in some fertility parameters in mice. Int. J. Sci. Res. 2017, 6, 260–266. [Google Scholar] [CrossRef]
- Imade, O.V.; Smith, O.F.; Gazal, O.; Adekunle, E.O.; Beshel, J.A. Effects of Lepidium sativum seed on reproductive characteristics in rabbit bucks. J. Phytopharm. 2020, 9, 89–95. [Google Scholar] [CrossRef]
- Aoki, Y.; Tsujimura, A.; Nagashima, Y.; Hiramatsu, I.; Uesaka, Y.; Nozaki, T.; Ogishima, T.; Shirai, M.; Shoyama, Y.; Tanaka, H.; et al. Effect of Lepidium meyenii on in vitro fertilization via improvement in acrosome reaction and motility of mouse and human sperm. Reprod. Med. Biol. 2018, 18, 57–64. [Google Scholar] [CrossRef]
- Ohta, Y.; Yoshida, K.; Kamiya, S.; Kawate, N.; Takahashi, M.; Inaba, T.; Hatoya, S.; Morii, H.; Takahashi, K.; Ito, M.; et al. Feeding hydroalcoholic extract powder of Lepidium meyenii (maca) increases serum testosterone concentration and enhances steroidogenic ability of Leydig cells in male rats. Andrologia 2016, 48, 347–354. [Google Scholar] [CrossRef]
- Getahun, T.; Sharma, V.; Gupta, N. Chemical composition, antibacterial and antioxidant activities of oils obtained by different extraction methods from Lepidium sativum L. seeds. Ind. Crops Prod. 2020, 156, 112876. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; Mahmoud, A.Z.; Fashori, N.N.; Alraraj, R.; Alsarra, I.A. Chemical composition and antimicrobial, antioxidant, and anti-inflammatory activities of Lepidium sativum seed oil. Saudi J. Biol. Sci. 2019, 26, 1089–1192. [Google Scholar] [CrossRef]
- Jahani, S.; Heidari, Z.; Azami, M.; Moudi, B. Comparison of anticancer effects of hydroalcoholic extracts of Camellia sinensis and Lepidium sativum L. on hela cell line. Int. J. Cancer Manag. 2020, 13, e98913. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Cruz-Martins, N.; López-Jornet, P.; Lopez, E.P.; Harun, N.; Yeskalieyeva, B.; Beyatli, A.; Sytar, O.; Calina, D.; Cho, W.C. Natural coumarins: Exploring the pharmacological complexity and underlying molecular mechanisms. Oxid Med. Cell Longev. 2021, 2021, 6492346. [Google Scholar] [CrossRef]
- Ibrahim, M.; Kebede, A. In vitro antibacterial activities of methanol and aqueous leave extracts of selected medicinal plants against human pathogenic bacteria. Saudi J. Biol. Sci. 2020, 27, 2261–2268. [Google Scholar] [CrossRef]
- Ivanisova, E.; Menhartova, K.; Terentjeva, M.; Harangozo, L.; Kantor, A.; Kacaniova, M. The evaluation of chemical, antioxidant, antimicrobial and sensory properties of kombucha tea beverage. J. Food Sci. Technol. 2020, 57, 1840–1846. [Google Scholar] [CrossRef]
- Luksic, L.; Arvay, J.; Vollmannova, A.; Toth, T.; Skrabanja, V.; Trcek, J.; Germ, M.; Kreft, I. Hydrothermal treatment of Tartary buckwheat grain hinders the transformation of rutin to quercetin. J. Cereal Sci. 2016, 72, 131–134. [Google Scholar] [CrossRef]
- Jambor, T.; Arvay, J.; Ivanisova, E.; Tvrda, E.; Kovacik, A.; Greifova, H.; Lukac, N. Investigation of the properties and effects of Salvia Officinalis L. on the viability, steroidogenesis and reactive oxygen species production in TM3 Leydig cells in vitro. Physiol. Res. 2020, 69, 661–673. [Google Scholar] [CrossRef]
- Jambor, T.; Arvay, J.; Tvrda, E.; Kovacik, A.; Greifova, H.; Lukac, N. The effect of Apium Graveolens L., Levisticum Officinale and Calendula Officinalis L. on cell viability, membrane integrity, steroidogenesis, and itracellular communication in mice Leydig cells in vitro. Physiol. Res. 2021, 70, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Shymanska, O.; Vergun, O.; Rakhmetov, D.; Brindza, J.; Ivanisova, E. A total content of phenolic compounds in the ethanol extracts of Galega officinalis L. and G. orientalis lam. Agrobiodiversity Improv. Nutr. Health Life Qual. 2018, 2, 140–145. [Google Scholar] [CrossRef]
- Willett, C.W. Balancing life-style and genomics research for disease prevention. Science 2002, 196, 695–698. [Google Scholar] [CrossRef]
- Ivanisova, E.; Grygorieva, O.; Abraamova, V.; Shubertova, Y.; Tarentjeva, M.; Brindya, J. Characterization of morphological parameters and biological activity of jujube fruit (Ziziphus jujuba Mill). J. Berr. Res. 2017, 7, 249–260. [Google Scholar] [CrossRef]
- Farmakope Polska. The Polish Pharmaceutical Society, 5th ed.; PTFarm: Warszawa, Poland, 1999; pp. 880–881. [Google Scholar]
- Grygorieva, O.; Vergun, O.; Klymenko, S.; Zhurba, M.; Horcinova, V.; Ivanisova, E.; Brindza, J. Estimation of phenolic compounds content and antioxidant activitz of leaves extracts of some selected non-traditional plants. Potravin. Slov. J. Food Sci. 2020, 14, 501–509. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Larrauri, J.A.; Saura-calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1999, 76, 270–276. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Yawer, A.; Sychrova, E.; Raska, J.; Babica, P.; Sovadinova, I. Endocrine-disrupting chemicals affect Sertoli TM4 cell functionality through dysregulation of gap junctional intercellular communication in vitro. Food Chem. Toxicol. 2022, 164, 113004. [Google Scholar] [CrossRef]
- Hamid, R.; Rotshtezyn, Y.; Rabadi, L.; Parikh, R.; Bullock, P. Comparison of alamar blue and MTT assays for high through-put screening. Toxicol. Vitr. 2004, 18, 703–710. [Google Scholar] [CrossRef]
- Schreer, A.; Tinson, C.; Sherry, J.P.; Schirmer, K. Application of Alamar blue/5-carboxyfluorescein diacetate acetoxymethyl ester as a noninvasive cell viability assay in primary hepatocytes from rainbow trout. Anal. Biochem. 2005, 344, 76–85. [Google Scholar] [CrossRef]
- Repetto, G.; Del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, J.W.; Cha, Y.N.; Kim, C. A quantitative nitroblue tetrazolium assay for determining intracellular superoxide anion production in phagocytic cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Babica, P.; Sovadinová, I.; Upham, B.L. Scrape Loading/Dye Transfer Assay. In Gap Junction Protocols; Vinken, M., Johnstone, S., Eds.; Humana Press: New York, NY, USA, 2016; Volume 1437, Methods in Molecular Biology. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hosseinzade, A.; Sadeghi, O.; Biregani, A.N.; Soukhtehzari, S.; Brandt, G.S.; Esmaillzadeh, A. Immunomodulatory effects of flavonoids: Possible induction of T CD4+ regulatory cells through suppression of mTOR pathway signaling activity. Front. Immunol. 2019, 10, 51. [Google Scholar] [CrossRef]
- Ghoora, M.D.; Haldipur, A.C.; Srividya, N. Comparative evaluation of phytochemical content, antioxidant capacities and overall antioxidant potential of select culinary microgreens. J. Agric. Food Res. 2020, 2, 100046. [Google Scholar] [CrossRef]
- Keutgen, N.; Hausknecht, M.; Tomasyewska-Sowa, M.; Keutgen, A.J. Nutritional and sensory quality of two tzpes of Cress microgrens depending on the mineral nutrition. Agronomy 2021, 11, 1110. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nkhel, C.; Pannico, A.; Grayiani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Pouphael, Y. Genotype-specific modulatory effects of select spectral bandwidths on the nutritive and phytochemical composition of microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in macronutrient content, phytochemical constitution and in vitro antioxidant capacity of green and red butterhead lettuce dictated by different developmental stages of harvest maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Gaspari, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Nutrient supplementation configures the bioactive profile and production characteristics of three Brassica L. microgreens species grown in peat-based media. Agronomy 2021, 11, 346. [Google Scholar] [CrossRef]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Drouet, S.; Hano, C.; Abbasi, B.H. Effect of ultraviolet-c radiation and melatonin stress on biosynthesis of antioxidant and antidiabetic metabolites produced in in vitro callus culture of Lepidium sativum L. Int. J. Mol. Sci. 2019, 20, 1787. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Chemical constituents and pharmacological effects of Lepidium sativum: A review. Int. J. Curr. Pharm. Res. 2019, 11, 1–10. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Kumar, N.V.A.; Yucca, P.; Varoni, E.M.; Dini, L.; Martins, N.; Martorell, M.; Docea, A.O.; Setyer, N.W.; Calinca, D.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Phzsiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Elspeiy, M.; Adbella, M.; Khalifah, A. Effects of oral administration of Lepidium sativum, moringa oleifera oils and aqueous extract of Vitex agnnus cactus on reproductive performance and blood biochemical of doe rabbits. Egyp. J. Rabb. Sci. 2021, 31, 1–24. [Google Scholar] [CrossRef]
- Asl, R.F.; Khosravi, M.; Hajikhani, R.; Solati, J.; Fahimi, H. Complementary effects of coenzyme Q10 and Lepidium sativum supplementation on the reproductive function of mice: An experimental study. Int. J. Reprod. Biomed. 2021, 19, 607–618. [Google Scholar] [CrossRef]
- Alshebbi, E.S.; Farshori, N.N.; Al-Oqail, M.M.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. Protective effect of Lepidium sativum seed extract against hydrogen peroxide-induced cytotoxicity and oxidative stress in human liver cells (HepG2). Pharm. Biol. 2016, 54, 314–321. [Google Scholar] [CrossRef]
- Aslani, E.; Naghsh, N.; Ranjbar, M. Cytotoxic effects of hydro-alcoholic extracts of cress (Lepidium Sativum)-made from different stages of the plant-on k562 Leukemia cell line. Hormozgan Med. J. 2014, 18, 411–419. [Google Scholar]
- Selek, S.; Koyuncu, I.; Caglar, H.G.; Bektas, I.; Yilmaz, M.A.; Gonel, A.; Akyuz, E. The evaluation of antioxidant and anticancer effects of Lepidium Sativum Subsp Spinescens L. methanol extract on cancer cells. Cell. Mol. Biol. 2018, 64, 72–80. [Google Scholar] [CrossRef]
- Greco, A.; Del Prete, C.; De Biase, D.; Palumbo, V.; Albanese, S.; Mancini, M.; Paciello, O.; Cocchia, N. Effects of oral administration of Lepidium Meyenii on morphology of mice testis and motility of epididymal sperm calls after tetrahydrocannabinol exposure. Front. Vet. Sci. 2021, 8, 692874. [Google Scholar] [CrossRef]
- Balgoon, M.J. Assessment of the protective effect of Lepidium sativum against aluminium/induced liver and kidney effects in albino rat. Biomed. Res. Int. 2019, 2019, 4516730. [Google Scholar] [CrossRef]
- Totland, M.Z.; Rasmussen, N.L.; Knudsen, L.M.; Leithe, E. Regulation of gap junction intercellular communication by connexin ubiquitination: Physiological and pathophysiological implications. Cell Mol. Life Sci. 2019, 77, 573–591. [Google Scholar] [CrossRef]
- Gilleron, J. Connexins as potential therapeutic targets for testis pathologies. Cell Mol. Med. 2015, 1, 1–3. [Google Scholar] [CrossRef]
- Dolati, P.; Khodabandeh, Y.; Yamiri, M.J.; Jamhiri, I.; Mehrabani, D. The effect of lead acetate and quercetin on the tight and gap junctions in the mouse testis. Biol. Trace Elem. Res. 2020, 198, 535–543. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Wu, X.; Li, C.; Huang, Y.; Zhou, H.; Cui, Y. Resveratrol sensitive colorectal cancer cells to cetuximab bz connexin 43 uprregulation-induced Akt inhibition. Front. Oncol. 2020, 10, 383. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, X.; Wu, Y.; Yhao, Q.; Liu, X.; Yhang, G.; Yhao, Y.; Ning, Y.; Wang, K.; Tan, Y.; et al. Synergistic effect of resveratrol and HSV-TK/GCV therapy on murine hepatoma cells. Cancer Biol. Ther. 2019, 20, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Chang, M.Y.; Chang, W.W.; Wang, W.K.; Liu, C.F.; Lin, S.T.; Lee, C.H. Resveratrol enhances chemosensitivity in mouse melanoma model through connexin 43 upregulation. Environ. Toxicol. 2015, 30, 877–886. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormone | Intra-Assay Variability (%) | Inter-Assay Variability (%) | Sensitivity |
|---|---|---|---|
| Progesterone | ≤4.0 | ≤9.3 | 0.05 ng/mL |
| Testosterone | ≤7.0 | ≤8.3 | 0.10 ng/mL |
| Parameter | Concentration |
|---|---|
| The total polyphenols content | 94.10 ± 6.72 mg GAE/g d.w. |
| The total flavonoids content | 139.05 ± 7.19 mg QE/g d.w. |
| The total phenolic acids content | 70.89 ± 3.10 mg CAE/g d.w. |
| Phenolic Compounds | Concentration (mg/kg d.w.) |
|---|---|
| 4 -OH benzoic acid | 74.64 (± 0.62) |
| caffeic acid | 28.69 (± 0.18) |
| trans p-coumaric acid | 22.67 (± 0.66) |
| rutin | 23.31 (± 2.08) |
| ferulic acid | 333.66 (± 0.64) |
| myricetin | 2.59 (± 0.10) |
| resveratrol | 43.04 (± 0.11) |
| quercetin | 3.32 (± 0.06) |
| cinnamic acid | 1.56 (± 0.34) |
| kaempferol | 2.38 (± 0.03) |
| Parameter | Value |
|---|---|
| DPPH assay | 9.43 ± 0.01 mg TEAC/g |
| ABTS assay | 105.95 ± 0.01mg TEAC/g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jambor, T.; Zajickova, T.; Arvay, J.; Ivanisova, E.; Tirdilova, I.; Knizatova, N.; Greifova, H.; Kovacik, A.; Galova, E.; Lukac, N. Exceptional Properties of Lepidium sativum L. Extract and Its Impact on Cell Viability, Ros Production, Steroidogenesis, and Intracellular Communication in Mice Leydig Cells In Vitro. Molecules 2022, 27, 5127. https://doi.org/10.3390/molecules27165127
Jambor T, Zajickova T, Arvay J, Ivanisova E, Tirdilova I, Knizatova N, Greifova H, Kovacik A, Galova E, Lukac N. Exceptional Properties of Lepidium sativum L. Extract and Its Impact on Cell Viability, Ros Production, Steroidogenesis, and Intracellular Communication in Mice Leydig Cells In Vitro. Molecules. 2022; 27(16):5127. https://doi.org/10.3390/molecules27165127
Chicago/Turabian StyleJambor, Tomas, Terezia Zajickova, Julius Arvay, Eva Ivanisova, Ivana Tirdilova, Nikola Knizatova, Hana Greifova, Anton Kovacik, Eliska Galova, and Norbert Lukac. 2022. "Exceptional Properties of Lepidium sativum L. Extract and Its Impact on Cell Viability, Ros Production, Steroidogenesis, and Intracellular Communication in Mice Leydig Cells In Vitro" Molecules 27, no. 16: 5127. https://doi.org/10.3390/molecules27165127