
Acrylamide-Derived Ionome, Metabolic, and Cell Cycle Alterations Are Alleviated by Ascorbic Acid in the Fission Yeast

 ,
,
Abstract
:1. Introduction
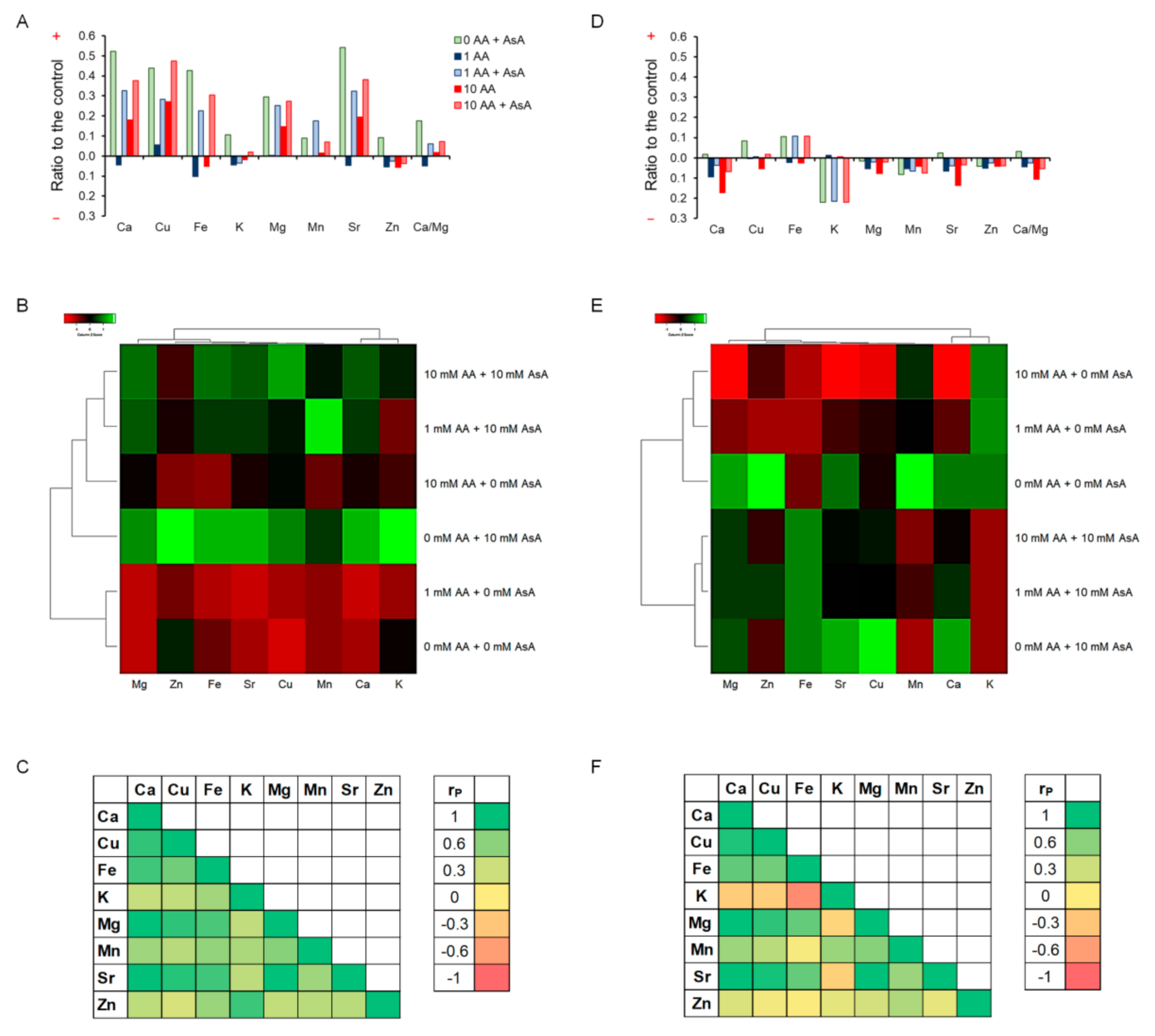
2. Results
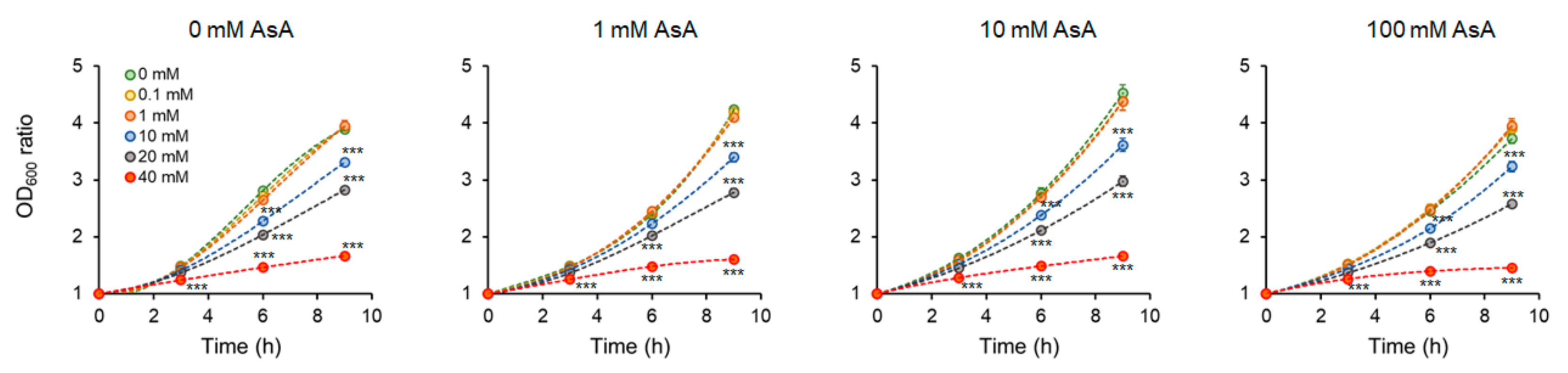
2.1. The time and Dose-Dependent Effect of Ascorbic Acid on the Cell Growth Intensity and the Impact of Acryl Amide Addition
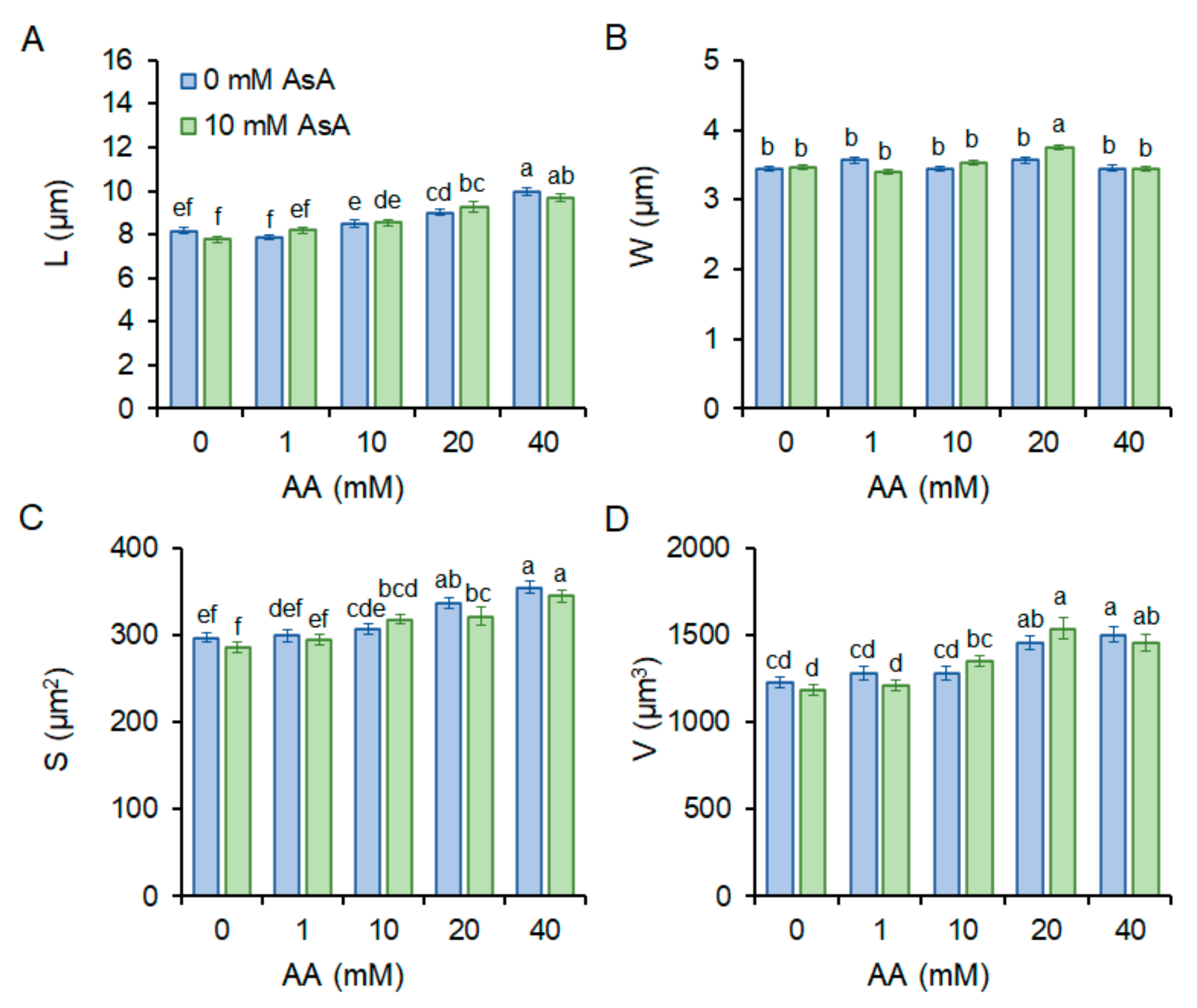
2.2. Cell Morphology
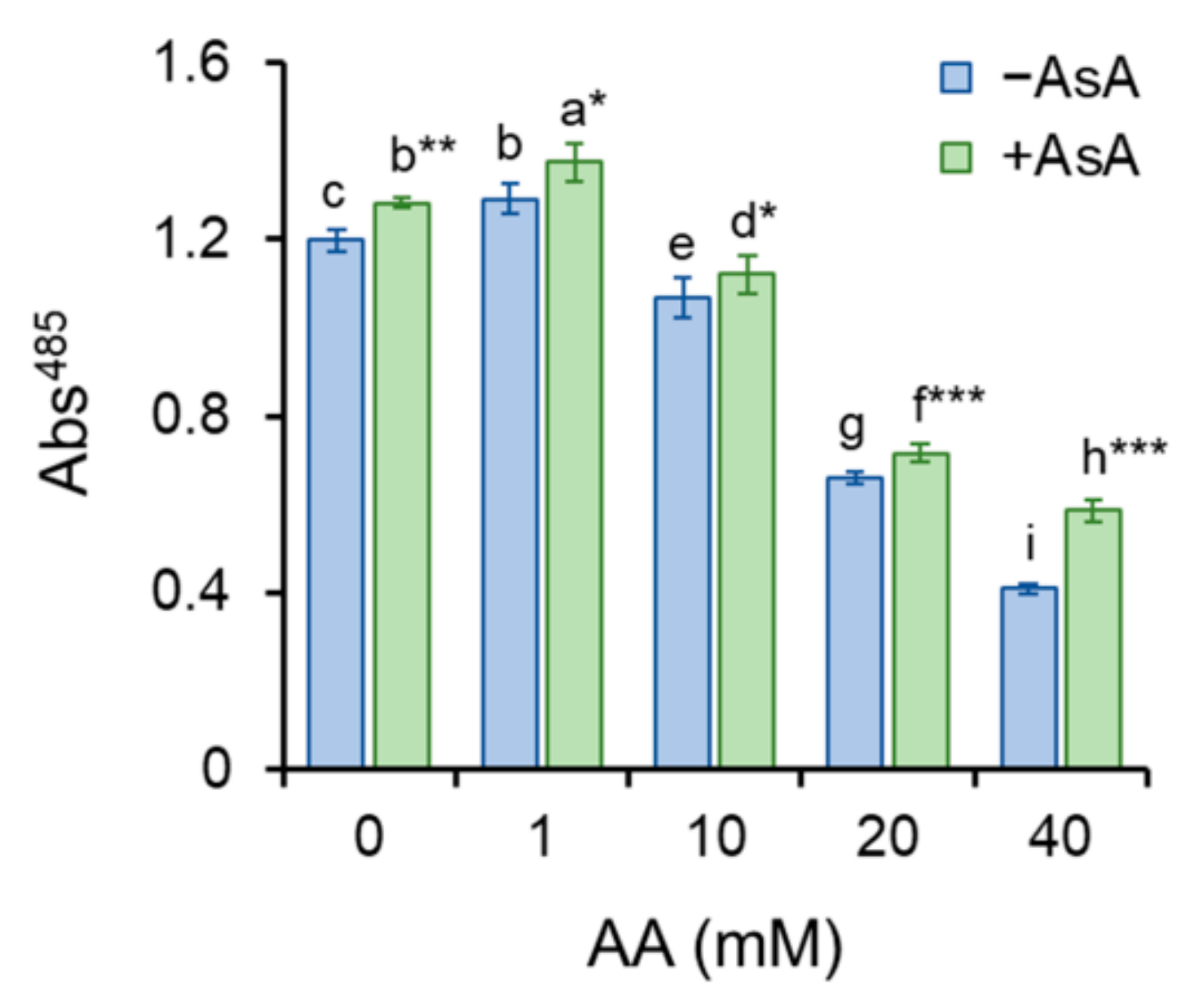
2.3. Cell Viability and Metabolic Activity
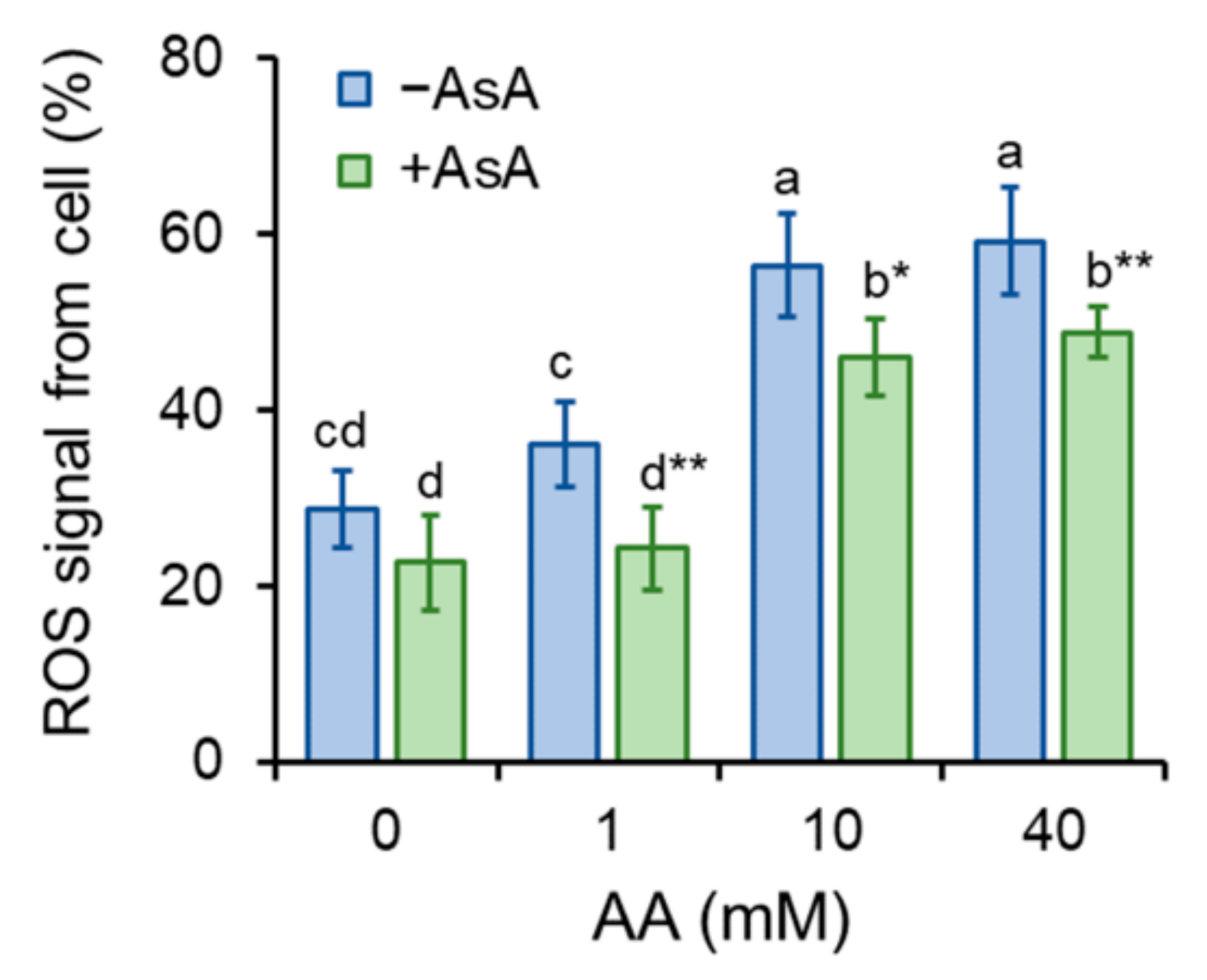
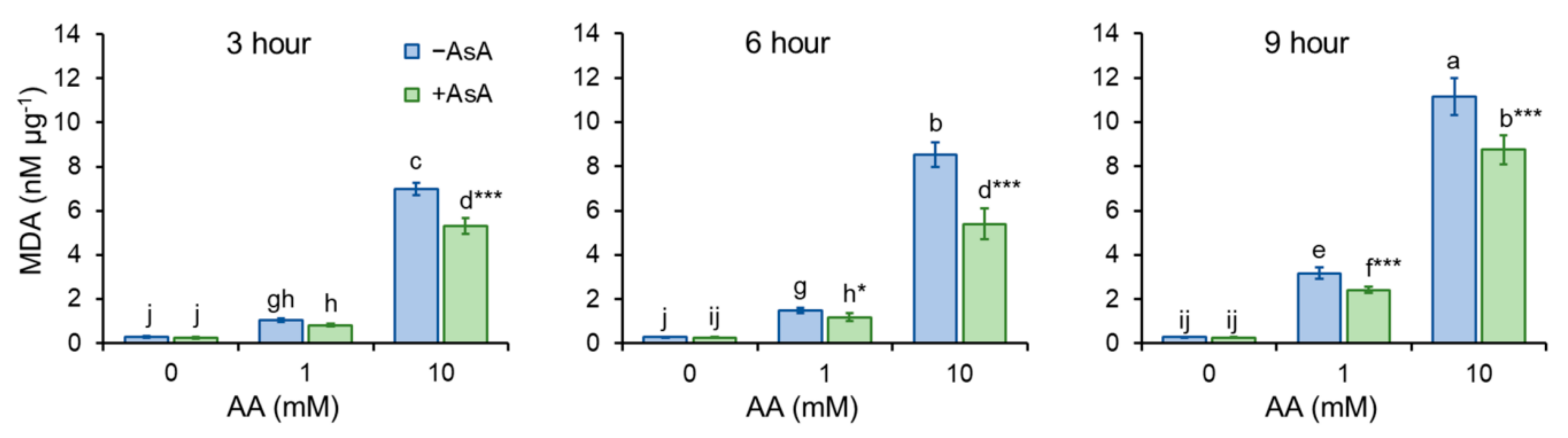
2.4. Ascorbic Acid Mitigates Aa-Induced Enhancement of ROS Production and Oxidative Stress
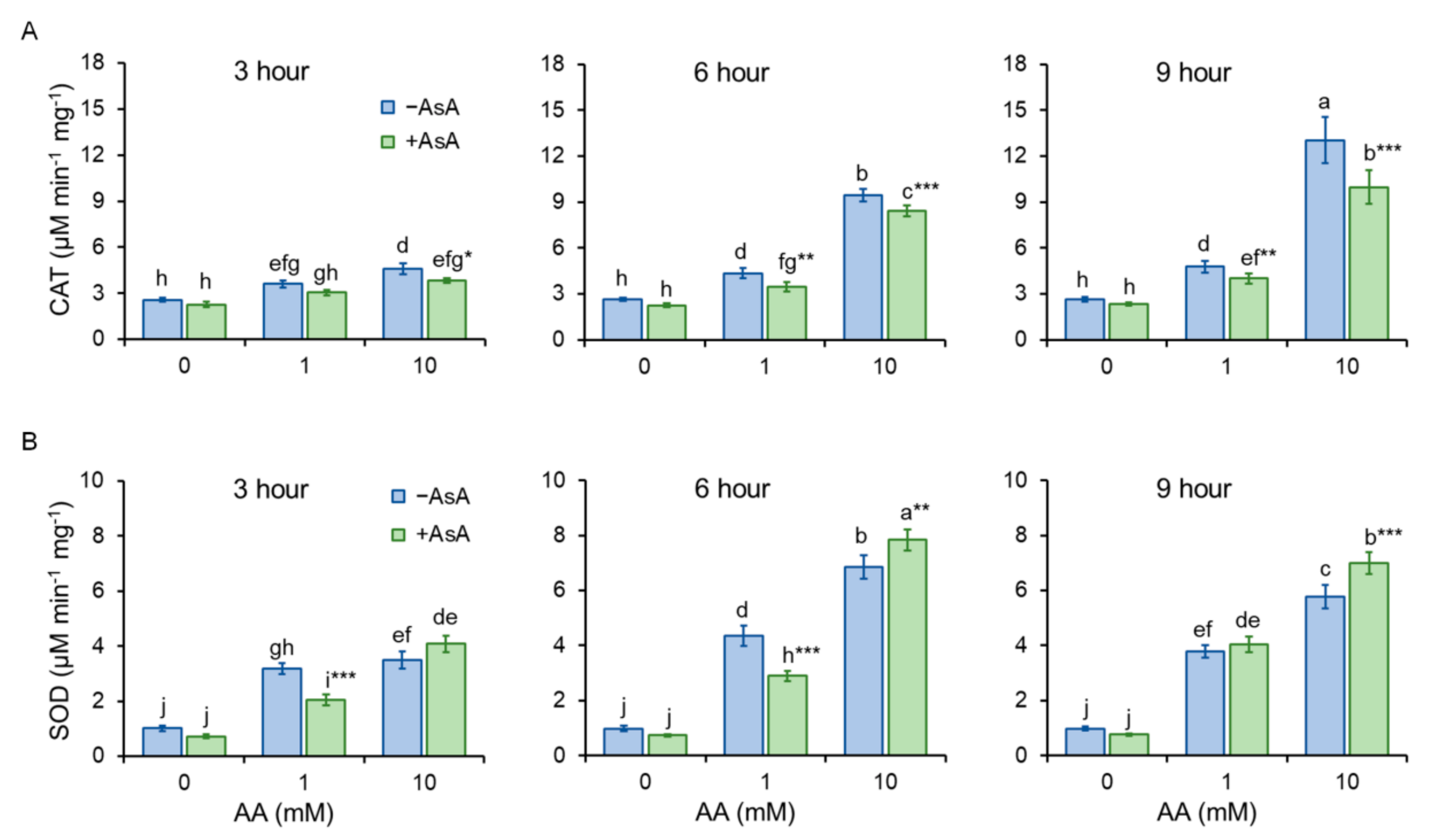
2.5. Acrylamide-Reduced Antioxidant Cell Capacity Is Significantly Restored by AsA
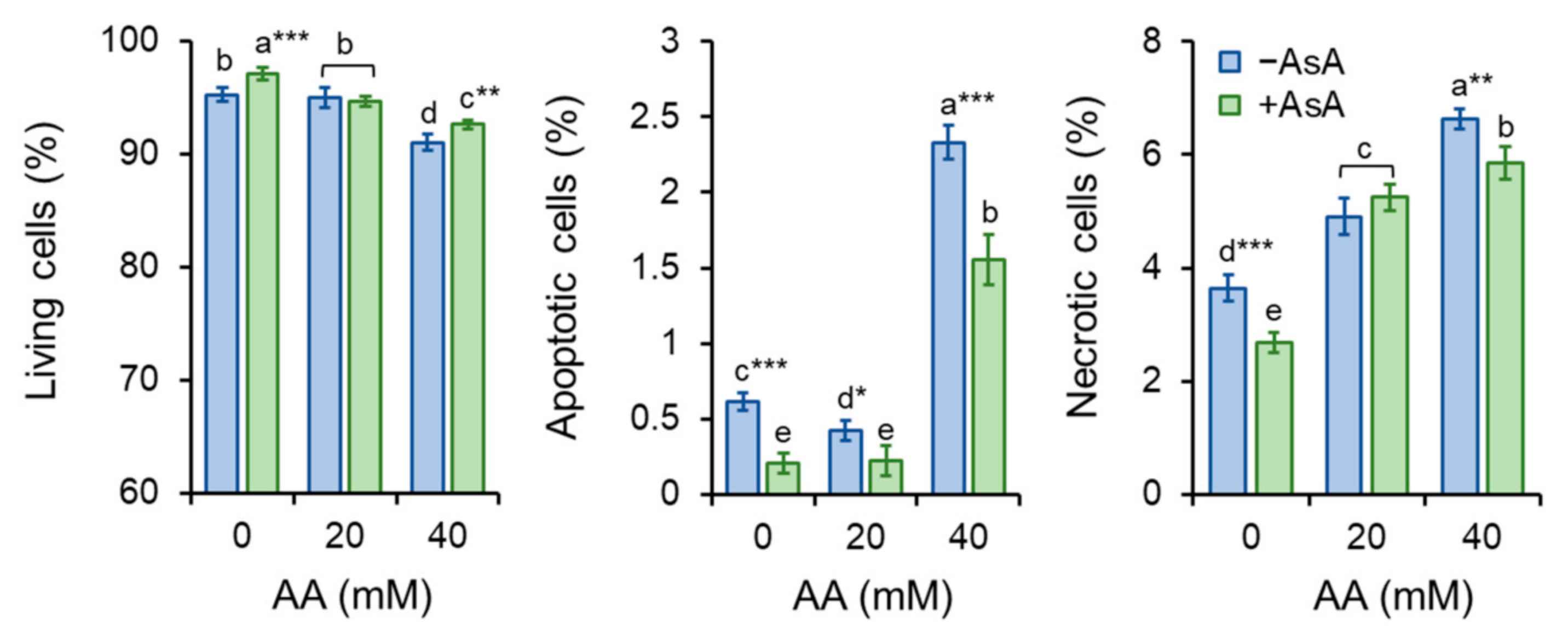
2.6. Acrylamide Toxicity Triggers Mild Apoptotic Events in S. pombe
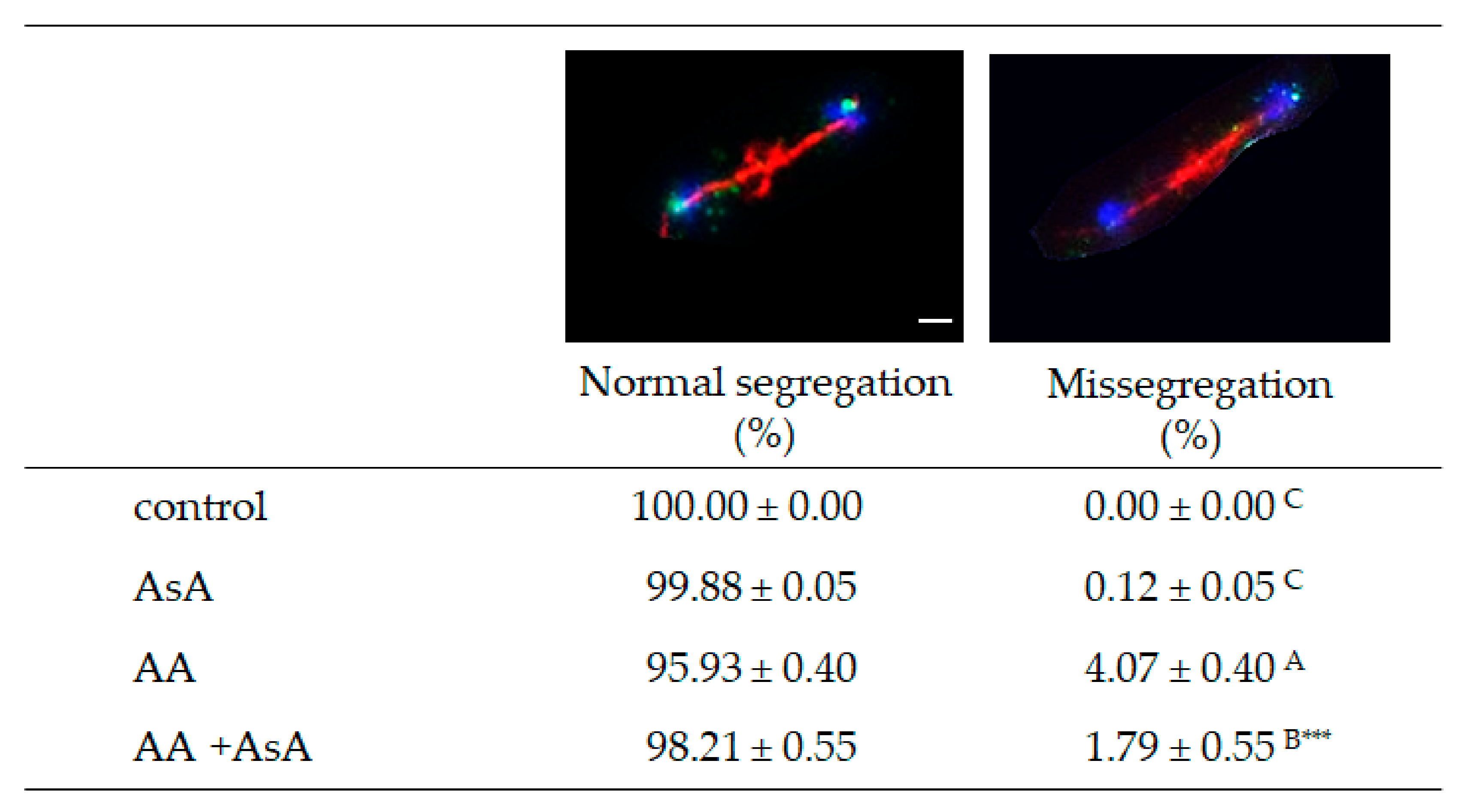
2.7. Acrylamide Triggers Errors in Chromosome Segregation during Cell Cycle
2.8. Ionome Balance
3. Discussion
4. Materials and Methods
4.1. Yeast Cultivation and Growth Conditions
4.2. Growth Intensity Determination
4.3. Relative Growth Rate (RGR) and Generation Time (gt)
4.4. IC50 Value
4.5. Characterization of the Yeast Morphology
4.6. Spot Test
4.7. Preparation of the Cell Extract for Biochemical Analyses
4.8. Metabolic Activity
4.9. Cell Viability
4.10. Biochemical Analysis
4.11. Determination of ROS Generation
4.12. Detection of Apoptosis and Necrosis
4.13. Determination of Ion Composition
4.14. Immunostaining and Fluorescence Microscopy
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jakobsen, L.S.; Granby, K.; Knudsen, V.K.; Nauta, M.; Pires, S.M.; Poulsen, M. Burden of disease of dietary exposure to acrylamide in Denmark. Food Chem. Toxicol. 2016, 90, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Parzefall, W. Minireview on the toxicity of dietary acrylamide. Food Chem. Toxicol. 2008, 46, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Chemistry, biochemistry, and safety of acrylamide. A review. J. Agric. Food Chem. 2003, 51, 4504–4526. [Google Scholar] [CrossRef] [PubMed]
- Adani, G.; Filippini, T.; Wise, L.A.; Halldorsson, T.I.; Blaha, L.; Vinceti, M. Dietary Intake of Acrylamide and Risk of Breast, Endometrial, and Ovarian Cancers: A Systematic Review and Dose–Response Meta-analysis. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Z.; Minghua, W.; Bo, N.; Chaoyue, Y.; Haiyang, Y.; Haiqing, Y.; Chunyu, X.; Yan, Z.; Yuan, Y. Rosmarinic acid attenuates acrylamide induced apoptosis of BRL-3A cells by inhibiting oxidative stress and endoplasmic reticulum stress. Food Chem. Toxicol. 2021, 151, 112156. [Google Scholar] [CrossRef]
- IARC. Monographs on the evaluation of carcinogenic risks to humans. Int. Agency Res. Cancer IARC 1994, 60, 389–433. Available online: https://publications.iarc.fr/Book-And-Report-Series/Iarc-Monographs-On-The-Identification-Of-Carcinogenic-Hazards-To-Humans/Some-Industrial-Chemicals-1994 (accessed on 2 May 2022).
- Shipp, A.; Lawrence, G.; Gentry, R.; McDonald, T.; Bartow, H.; Bounds, J.; Macdonald, N.; Clewell, H.; Allen, B.; Van Landingham, C. Acrylamide: Review of toxicity data and dose-response analyses for cancer and noncancer effects. Crit. Rev. Toxicol. 2006, 36, 481–608. [Google Scholar] [CrossRef]
- Kacar, S.; Sahinturk, V.; Kutlu, H.M. Effect of acrylamide on BEAS-2B normal human lung cells: Cytotoxic, oxidative, apoptotic and morphometric analysis. Acta Histochem. 2019, 121, 595–603. [Google Scholar] [CrossRef]
- Song, D.; Xu, C.; Holck, A.L.; Liu, R. Acrylamide inhibits autophagy, induces apoptosis and alters cellular metabolic profiles. Ecotoxicol. Environ. Saf. 2021, 208, 111543. [Google Scholar] [CrossRef]
- Fuhr, U.; Boettcher, M.I.; Kinzig-Schippers, M.; Weyer, A.; Jetter, A.; Lazar, A.; Taubert, D.; Tomalik-Scharte, D.; Pournara, P.; Jakob, V.; et al. Toxicokinetics of Acrylamide in Humans after Ingestion of a Defined Dose in a Test Meal to Improve Risk Assessment for Acrylamide Carcinogenicity. Cancer Epidemiol. Biomark. Prev. 2006, 15, 266–271. [Google Scholar] [CrossRef] [Green Version]
- Kwolek-Mirek, M.; Zadrąg-Tęcza, R.; Bednarska, S.; Bartosz, G. Yeast Saccharomyces cerevisiae devoid of Cu,Zn-superoxide dismutase as a cellular model to study acrylamide toxicity. Toxicol. Vitr. 2011, 25, 573–579. [Google Scholar] [CrossRef]
- Albalawi, A.; Alhasani, R.H.A.; Biswas, L.; Reilly, J.; Shu, X. Protective effect of carnosic acid against acrylamide-induced toxicity in RPE cells. Food Chem. Toxicol. 2017, 108, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Kommuguri, U.N.; Pallem, P.V.S.; Bodiga, S.; Bodiga, V.L. Effect of dietary antioxidants on the cytostatic effect of acrylamide during copper-deficiency in Saccharomyces cerevisiae. Food Funct. 2014, 5, 705–715. [Google Scholar] [CrossRef]
- Ahmad Bainmahfouz, F.R.; Ali, S.S.; Al-Shali, R.A.; El-Shitany, N.A.E.-A. Vitamin E and 5-amino salicylic acid ameliorates acrylamide-induced peripheral neuropathy by inhibiting caspase-3 and inducible nitric oxide synthase immunoexpression. J. Chem. Neuroanat. 2021, 113, 101935. [Google Scholar] [CrossRef]
- Chen, H.; Gu, Z. Effect of Ascorbic Acid on the Properties of Ammonia Caramel Colorant Additives and Acrylamide Formation. J. Food Sci. 2014, 79, C1678–C1682. [Google Scholar] [CrossRef]
- Zheng, L.; Zhu, H.-Z.; Wang, B.-T.; Zhao, Q.-H.; Du, X.-B.; Zheng, Y.; Jiang, L.; Ni, J.-Z.; Zhang, Y.; Liu, Q. Sodium selenate regulates the brain ionome in a transgenic mouse model of Alzheimer’s disease. Sci. Rep. 2016, 6, 39290. [Google Scholar] [CrossRef] [Green Version]
- López-Orenes, A.; Bueso, M.C.; Conesa, H.; Calderón, A.A.; Ferrer, M.A. Seasonal ionomic and metabolic changes in Aleppo pines growing on mine tailings under Mediterranean semi-arid climate. Sci. Total Environ. 2018, 637–638, 625–635. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Wood, V.; Fantes, P.A. An Ancient Yeast for Young Geneticists: A Primer on the Schizosaccharomyces pombe Model System. Genetics 2015, 201, 403–423. [Google Scholar] [CrossRef] [Green Version]
- Hayles, J.; Nurse, P. Introduction to Fission Yeast as a Model System. Cold Spring Harb. Protoc. 2018, 2018, pdb.top079749. [Google Scholar] [CrossRef] [Green Version]
- Villagran, M.; Ferreira, J.; Martorell, M.; Mardones, L. The Role of Vitamin C in Cancer Prevention and Therapy: A Literature Review. Antioxidants 2021, 10, 1894. [Google Scholar] [CrossRef]
- Vo, T.V.; Das, J.; Meyer, M.J.; Cordero, N.A.; Akturk, N.; Wei, X.; Fair, B.J.; Degatano, A.G.; Fragoza, R.; Liu, L.G.; et al. A Proteome-wide Fission Yeast Interactome Reveals Network Evolution Principles from Yeasts to Human. Cell 2016, 164, 310–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, A.; Freitas, A.V.; Ralston, Z.A.; Tang, Z. Fission Yeast Schizosaccharomyces pombe: A Unicellular “Micromammal” Model Organism. Curr. Protoc. 2021, 1, e151. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.A.; Russell, P. Cell cycle regulation in Schizosaccharomyces pombe. Curr. Opin. Microbiol. 2000, 3, 631–636. [Google Scholar] [CrossRef]
- Boonstra, J.; Post, J.A. Molecular events associated with reactive oxygen species and cell cycle progression in mammalian cells. Gene 2004, 337, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, G.; Palchaudhuri, S.; Dixit, S.; Chattopadhyay, D. MAPK mediated cell cycle regulation is associated with Cdc25 turnover in S. pombe after exposure to genotoxic stress. Cell Cycle 2008, 7, 365–372. [Google Scholar] [CrossRef]
- Yang, L.; Dong, L.; Zhang, L.; Bai, J.; Chen, F.; Luo, Y. Acrylamide Induces Abnormal mtDNA Expression by Causing Mitochondrial ROS Accumulation, Biogenesis, and Dynamics Disorders. J. Agric. Food Chem. 2021, 69, 7765–7776. [Google Scholar] [CrossRef]
- Agus, H.H.; Kok, G.; Derinoz, E.; Oncel, D.; Yilmaz, S. Involvement of Pca1 in ROS-mediated apoptotic cell death induced by alpha-thujone in the fission yeast (Schizosaccharomyces pombe). FEMS Yeast Res. 2020, 20, foaa022. [Google Scholar] [CrossRef]
- Huang, Y.-S.; Hsieh, T.-J.; Lu, C.-Y. Simple analytical strategy for MALDI-TOF-MS and nanoUPLC—MS/MS: Quantitating curcumin in food condiments and dietary supplements and screening of acrylamide-induced ROS protein indicators reduced by curcumin. Food Chem. 2015, 174, 571–576. [Google Scholar] [CrossRef]
- Hung, C.H.; Lin, Y.C.; Tsai, Y.G.; Lin, Y.C.; Kuo, C.H.; Tsai, M.L.; Kuo, C.H.; Liao, W.T. Acrylamide Induces Mitophagy and Alters Macrophage Phenotype via Reactive Oxygen Species Generation. Int. J. Mol. Sci. 2021, 22, 1683. [Google Scholar] [CrossRef]
- Nowak, A.; Zakłos-Szyda, M.; Żyżelewicz, D.; Koszucka, A.; Motyl, I. Acrylamide Decreases Cell Viability, and Provides Oxidative Stress, DNA Damage, and Apoptosis in Human Colon Adenocarcinoma Cell Line Caco-2. Molecules 2020, 25, 368. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Liu, C.; Jiang, M.; Feng, L.; Chen, Y.; Han, J. After In Vitro Digestion, Jackfruit Flake Affords Protection against Acrylamide-Induced Oxidative Damage. Molecules 2019, 24, 3322. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Feng, L.; Shen, Y.; Su, H.; Li, Y.; Zhuang, J.; Zhang, L.; Zheng, X. Myricitrin Inhibits Acrylamide-Mediated Cytotoxicity in Human Caco-2 Cells by Preventing Oxidative Stress. BioMed Res. Int. 2013, 2013, e724183. [Google Scholar] [CrossRef]
- Cao, J.; Liu, Y.; Jia, L.; Jiang, L.-P.; Geng, C.-Y.; Yao, X.-F.; Kong, Y.; Jiang, B.-N.; Zhong, L.-F. Curcumin Attenuates Acrylamide-Induced Cytotoxicity and Genotoxicity in HepG2 Cells by ROS Scavenging. J. Agric. Food Chem. 2008, 56, 12059–12063. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.; Li, Y.; Bao, T.; Gowd, V.; Chen, W. Protective property of mulberry digest against oxidative stress—A potential approach to ameliorate dietary acrylamide-induced cytotoxicity. Food Chem. 2017, 230, 306–315. [Google Scholar] [CrossRef]
- Orta Yilmaz, B.; Yildizbayrak, N.; Aydin, Y. Vitamin C inhibits glycidamide-induced genotoxicity and apoptosis in Sertoli cells. J. Biochem. Mol. Toxicol. 2020, 34, e22545. [Google Scholar] [CrossRef]
- Catalgol, B.; Ozhan, G.; Alpertunga, B. Acrylamide-induced oxidative stress in human erythrocytes. Hum. Exp. Toxicol. 2009, 28, 611–617. [Google Scholar] [CrossRef]
- Celik, F.S.; Cora, T.; Yigin, A.K. Investigation of Genotoxic and Cytotoxic Effects of Acrylamide in HEK293 Cell Line. J. Cancer Prev. Curr. Res. 2018, 9, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Gül, M.; Kayhan Kuştepe, E.; Erdemli, M.E.; Altınöz, E.; Gözükara Bağ, H.G.; Gül, S.; Göktürk, N. Protective effects of crocin on acrylamide-induced testis damage. Andrologia 2021, 53, e14176. [Google Scholar] [CrossRef]
- Reshmitha, T.R.; Nisha, P. Lycopene mitigates acrylamide and glycidamide induced cellular toxicity via oxidative stress modulation in HepG2 cells. J. Funct. Foods 2021, 80, 104390. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.M.; Elsawy, H.; Alzahrani, A.M.; Ali, A.; Mahmoud, O. Silymarin Ameliorates Acrylamide-Induced Hyperlipidemic Cardiomyopathy in Male Rats. BioMed Res. Int. 2019, 2019, e4825075. [Google Scholar] [CrossRef]
- Požgajová, M.; Navrátilová, A.; Šebová, E.; Kovár, M.; Kačániová, M. Cadmium-Induced Cell Homeostasis Impairment is Suppressed by the Tor1 Deficiency in Fission Yeast. Int. J. Mol. Sci. 2020, 21, 7847. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, F.; Wang, C.; Wu, M.; Zhang, X.; Wang, Q.; Yao, X.; Fu, C.; Zhang, X.; Zang, J. Phosphorylation of CENP-C by Aurora B facilitates kinetochore attachment error correction in mitosis. Proc. Natl. Acad. Sci. USA 2017, 114, E10667–E10676. [Google Scholar] [CrossRef] [Green Version]
- Hálová, L.; Petersen, J. Aurora promotes cell division during recovery from TOR-mediated cell cycle arrest by driving spindle pole body recruitment of Polo. J. Cell Sci. 2011, 124, 3441–3449. [Google Scholar] [CrossRef] [Green Version]
- Sickles, D.W.; Sperry, A.O.; Testino, A.; Friedman, M. Acrylamide effects on kinesin-related proteins of the mitotic/meiotic spindle. Toxicol. Appl. Pharmacol. 2007, 222, 111–121. [Google Scholar] [CrossRef]
- Adler, I.-D.; Zouh, R.; Schmid, E. Perturbation of cell division by acrylamide in vitro and in vivo. Mutat. Res. Lett. 1993, 301, 249–254. [Google Scholar] [CrossRef]
- Chen, J.-H.; Tsou, T.-C.; Chiu, I.-M.; Chou, C.-C. Proliferation Inhibition, DNA Damage, and Cell-Cycle Arrest of Human Astrocytoma Cells after Acrylamide Exposure. Chem. Res. Toxicol. 2010, 23, 1449–1458. [Google Scholar] [CrossRef]
- Hassan, H.A.; EL-Kholy, W.M.; EL-Sawi, M.R.F.; Galal, N.A.; Ramadan, M.F. Myrtle (Myrtus communis) leaf extract suppresses hepatotoxicity induced by monosodium glutamate and acrylamide through obstructing apoptosis, DNA fragmentation, and cell cycle arrest. Environ. Sci. Pollut. Res. 2020, 27, 23188–23198. [Google Scholar] [CrossRef]
- Pozgajova, M.; Navratilova, A.; Arvay, J.; Duranova, H.; Trakovicka, A. Impact of cadmium and nickel on ion homeostasis in the yeast Schizosaccharomyces pombe. J. Environ. Sci. Health B 2020, 55, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Popa, C.V.; Dumitru, I.; Ruta, L.L.; Danet, A.F.; Farcasanu, I.C. Exogenous Oxidative Stress Induces Ca2+ Release in the Yeast Saccharomyces Cerevisiae. FEBS J. 2010, 277, 4027–4038. [Google Scholar] [CrossRef] [PubMed]
- Navrátilová, A.; Kovár, M.; Požgajová, M. Ascorbic acid mitigates cadmium-induced stress, and contributes to ionome stabilization in fission yeast. Environ. Sci. Pollut. Res. 2021, 28, 15380–15393. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Danku, J.; Baxter, I.; Kim, S.; Vatamaniuk, O.K.; Vitek, O.; Ouzzani, M.; Salt, D.E. High-resolution genome-wide scan of genes, gene-networks and cellular systems impacting the yeast ionome. BMC Genom. 2012, 13, 623. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.; Russell, P. Growth and the Environment of Schizosaccharomyces pombe. Cold Spring Harb. Protoc. 2016, 2016, pdb.top079764. [Google Scholar] [CrossRef] [Green Version]
- Bayliak, M.M.; Burdylyuk, N.I.; Lushchak, V.I. Quercetin increases stress resistance in the yeast Saccharomyces cerevisiae not only as an antioxidant. Ann. Microbiol. 2016, 66, 569–576. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ďurovcová, I.; Goffa, E.; Šestáková, Z.; Mániková, D.; Gaplovská-Kyselá, K.; Chovanec, M.; Ševčovičová, A. Acute Exposure to Bisphenol A Causes Oxidative Stress Induction with Mitochondrial Origin in Saccharomyces cerevisiae Cells. J. Fungi 2021, 7, 543. [Google Scholar] [CrossRef]
- Sousa, C.A.; Soares, H.M.V.M.; Soares, E.V. Nickel Oxide Nanoparticles Trigger Caspase- and Mitochondria-Dependent Apoptosis in the Yeast Saccharomyces cerevisiae. Chem. Res. Toxicol. 2019, 32, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Pozgajova, M.; Cipak, L.; Trakovicka, A. Prp4 kinase is required for proper segregation of chromosomes during meiosis in Schizosaccharomyces pombe. Acta Biochim. Pol. 2013, 60, 871–873. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AsA | IC50 (mM of AA) | R2 |
|---|---|---|
| 0 mM | 30.8337 | 0.932 |
| 10 mM | 37.8238 | 0.916 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovár, M.; Navrátilová, A.; Kolláthová, R.; Trakovická, A.; Požgajová, M. Acrylamide-Derived Ionome, Metabolic, and Cell Cycle Alterations Are Alleviated by Ascorbic Acid in the Fission Yeast. Molecules 2022, 27, 4307. https://doi.org/10.3390/molecules27134307
Kovár M, Navrátilová A, Kolláthová R, Trakovická A, Požgajová M. Acrylamide-Derived Ionome, Metabolic, and Cell Cycle Alterations Are Alleviated by Ascorbic Acid in the Fission Yeast. Molecules. 2022; 27(13):4307. https://doi.org/10.3390/molecules27134307
Chicago/Turabian StyleKovár, Marek, Alica Navrátilová, Renata Kolláthová, Anna Trakovická, and Miroslava Požgajová. 2022. "Acrylamide-Derived Ionome, Metabolic, and Cell Cycle Alterations Are Alleviated by Ascorbic Acid in the Fission Yeast" Molecules 27, no. 13: 4307. https://doi.org/10.3390/molecules27134307