
Identification and Evaluation of Traditional Chinese Medicine Natural Compounds as Potential Myostatin Inhibitors: An In Silico Approach

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Method and Materials
2.1. Preparation of Target Protein and Natural Compounds Library
2.2. Evaluation of Potential Leads and Drug-Ability
2.3. Visualization and Assessment of MSTN Protein
2.4. Molecular Dynamics (MD) Simulations
3. Result and Discussion
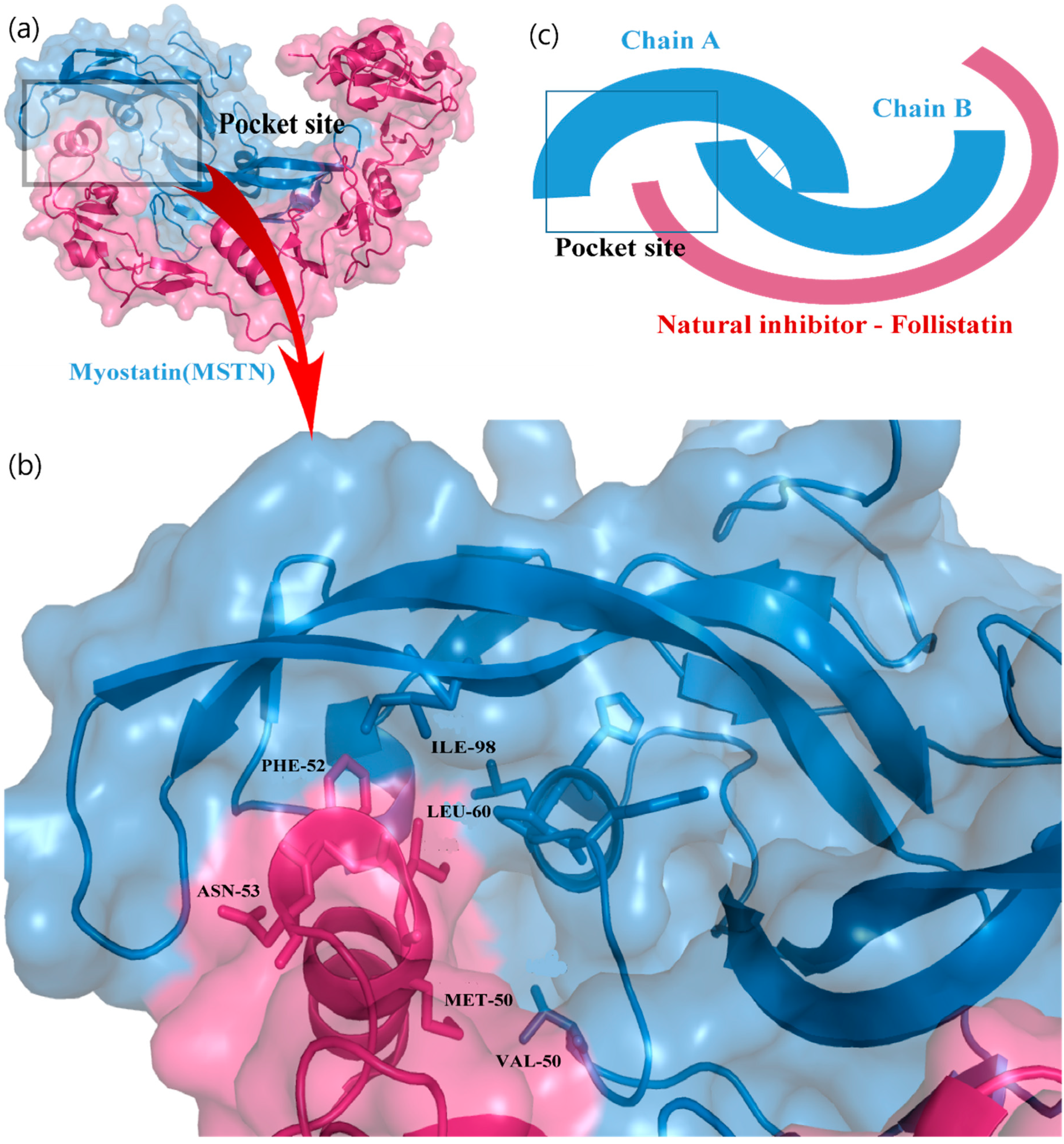
3.1. Active Pocket Analysis
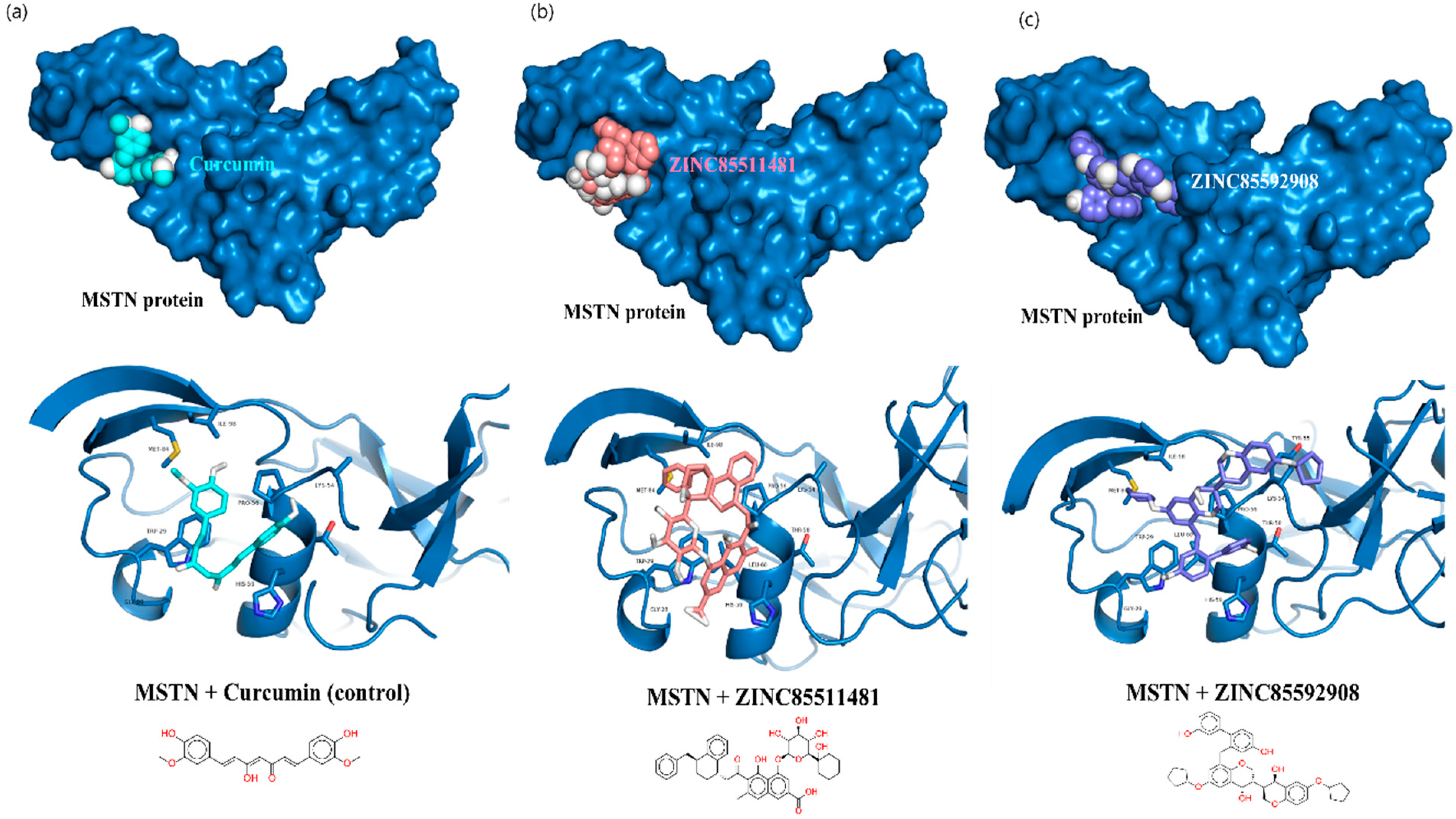
3.2. Molecular Docking, Hit Selection, and Drug-Ability Assessment
3.3. Interaction Analysis of MSTN Complexes
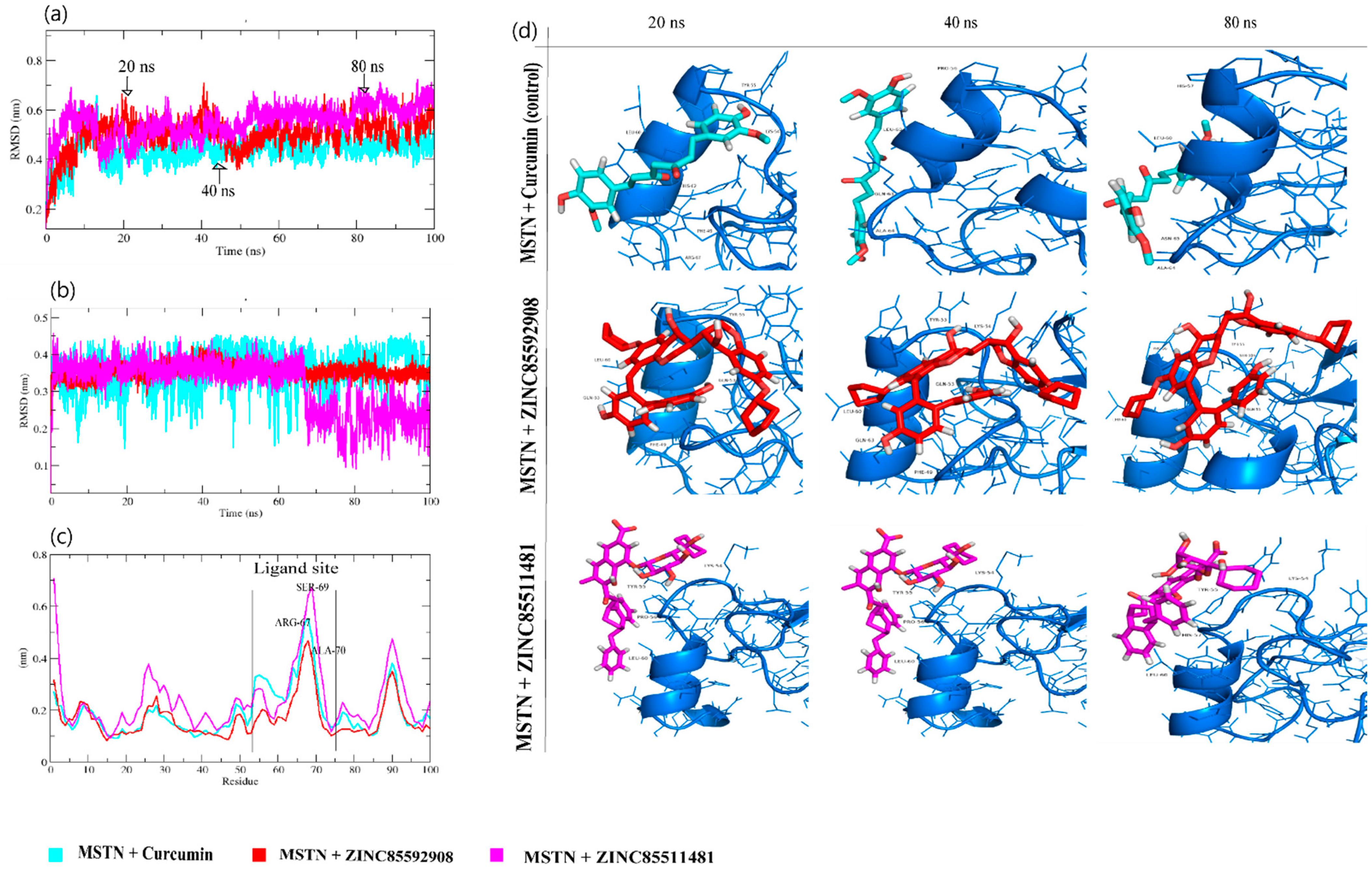
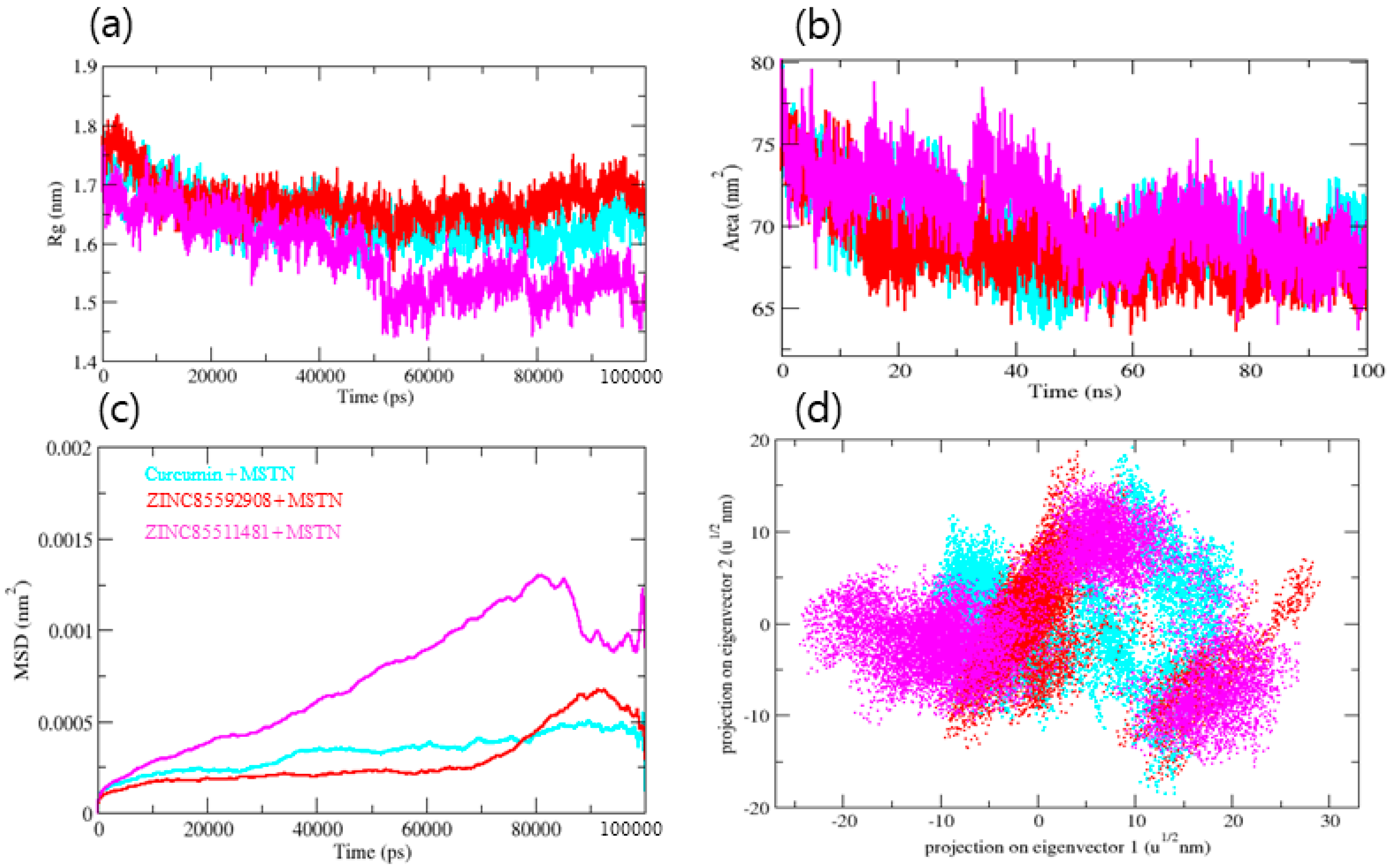
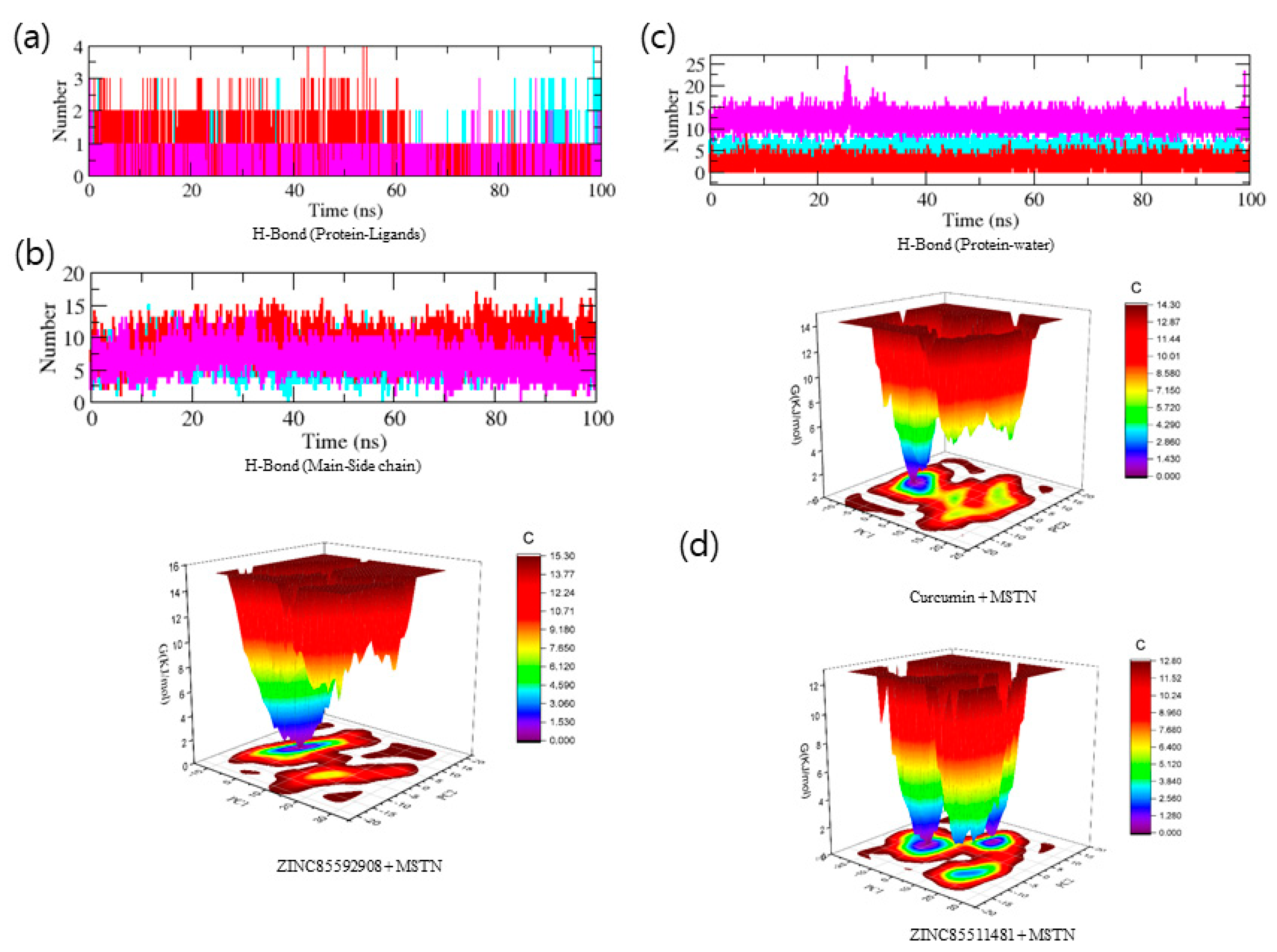
3.4. Molecular Dynamics Trajectory Analysis of MSTN-Ligand Complexes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thalacker-Mercer, A.; Riddle, E.; Barre, L. Protein and amino acids for skeletal muscle health in aging. Adv. Food Nutr. Res. 2019, 91, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Garry, G.A.; Antony, M.L.; Garry, D.J. Cardiotoxin Induced Injury and Skeletal Muscle Regeneration. Methods Mol. Biol. 2016, 1460, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Yablonka-Reuveni, Z.; Day, K.; Vine, A.; Shefer, G. Defining the transcriptional signature of skeletal muscle stem cells. J. Anim. Sci. 2008, 86, E207–E216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipina, C.; Hundal, H.S. Lipid modulation of skeletal muscle mass and function. J. Cachex. Sarcopenia Muscle 2016, 8, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Elkasrawy, M.N.; Hamrick, M.W. Myostatin (GDF-8) as a key factor linking muscle mass and bone structure. J. Musculoskelet. Neuronal Interact. 2010, 10, 56–63. [Google Scholar]
- Consitt, L.; Clark, B. The vicious cycle of myostatin signaling in sarcopenic obesity: Myostatin role in skeletal muscle growth, insulin signaling and implications for clinical trials. J. Frailty Aging 2017, 7, 21–27. [Google Scholar] [CrossRef]
- Allendorph, G.P.; Isaacs, M.J.; Kawakami, Y.; Belmonte, J.C.I.; Choe, S. BMP-3 and BMP-6 Structures Illuminate the Nature of Binding Specificity with Receptors. Biochemistry 2007, 46, 12238–12247. [Google Scholar] [CrossRef]
- Lee, S.-J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Kishioka, Y.; Wakamatsu, J.-I.; Hattori, A.; Nishimura, T. Interaction between myostatin and extracellular matrix components. Anim. Sci. J. 2010, 81, 102–107. [Google Scholar] [CrossRef]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ashraf, J.M.; Nahm, S.; Kim, Y.; Park, S.; Choi, I. Fibromodulin: A master regulator of myostatin controlling progression of satellite cells through a myogenic program. FASEB J. 2016, 30, 2708–2719. [Google Scholar] [CrossRef]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ahmad, K.; Malik, A.; Rabbani, G.; Kim, T.; Lee, I.; Lee, Y.H.; Park, S.; et al. Fibromodulin and regulation of the intricate balance between myoblast differentiation to myocytes or adipocyte-like cells. FASEB J. 2018, 32, 768–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Jan, A.T.; Baig, M.H.; Lee, E.J.; Choi, I. Matrix gla protein: An extracellular matrix protein regulates myostatin expression in the muscle developmental program. Life Sci. 2017, 172, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Ahmad, K.; Shaikh, S.; Jan, A.T.; Seo, M.-G.; Lee, E.J.; Choi, I. Dermatopontin in Skeletal Muscle Extracellular Matrix Regulates Myogenesis. Cells 2019, 8, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Ahmad, S.S.; Lim, J.H.; Ahmad, K.; Shaikh, S.; Lee, Y.-S.; Park, S.J.; Jin, J.O.; Lee, Y.-H.; Choi, I. Interaction of Fibromodulin and Myostatin to Regulate Skeletal Muscle Aging: An Opposite Regulation in Muscle Aging, Diabetes, and Intracellular Lipid Accumulation. Cells 2021, 10, 2083. [Google Scholar] [CrossRef]
- Bradley, L.; Yaworsky, P.J.; Walsh, F.S. Myostatin as a therapeutic target for musculoskeletal disease. Cell. Mol. Life Sci. 2008, 65, 2119–2124. [Google Scholar] [CrossRef]
- Baig, M.H.; Jan, A.T.; Rabbani, G.; Ahmad, K.; Ashraf, J.M.; Kim, T.; Min, H.S.; Lee, Y.H.; Cho, W.-K.; Ma, J.Y.; et al. Methylglyoxal and Advanced Glycation End products: Insight of the regulatory machinery affecting the myogenic program and of its modulation by natural compounds. Sci. Rep. 2017, 7, 5916. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- DeLano, W.L. The PyMOL Molecular Graphics System; Delano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Biovia, D.S. Discovery Studio Modeling Environment; Dassault Systèmes: San Diego, CA, USA, 2015. [Google Scholar]
- Irwin, J.J.; Shoichet, B.K. ZINC—A Free Database of Commercially Available Compounds for Virtual Screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ai, H.; Chen, W.; Yin, Z.; Hu, H.; Zhu, J.; Zhao, J.; Zhao, Q.; Liu, H. CarcinoPred-EL: Novel models for predicting the carcinogenicity of chemicals using molecular fingerprints and ensemble learning methods. Sci. Rep. 2017, 7, 2118. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.B.; Holloway, G.A. New Substructure Filters for Removal of Pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterling, T.; Irwin, J.J. ZINC 15–ligand discovery for everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Pol-Fachin, L.; Fernandes, C.L.; Verli, H. GROMOS96 43a1 performance on the characterization of glycoprotein conformational ensembles through molecular dynamics simulations. Carbohydr. Res. 2009, 344, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Schuttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. Sect. Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.-M.; Yang, Z.; Liu, C.-W.; Wang, R.; Tien, P.; Dale, R.; Sun, L.-Q. Myostatin antisense RNA-mediated muscle growth in normal and cancer cachexia mice. Gene Ther. 2007, 15, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Kasamatsu, S.; Shinozaki, S.; Yasuhara, S.; Kaneki, M. Myostatin deficiency not only prevents muscle wasting but also improves survival in septic mice. Am. J. Physiol. Metab. 2021, 320, E150–E159. [Google Scholar] [CrossRef]
- Wallner, C.; Huber, J.; Drysch, M.; Schmidt, S.V.; Wagner, J.M.; Dadras, M.; Lehnhardt, M.; Behr, B. Myostatin Upregulation in Patients in the Chronic Phase of Severe Burn Injury Leads to Muscle Cell Catabolism. Eur. Surg. Res. 2019, 60, 86–96. [Google Scholar] [CrossRef]
- Tsuchida, K. Targeting myostatin for therapies against muscle-wasting disorders. Curr. Opin. Drug Discov. Dev. 2008, 11, 487–494. [Google Scholar]
- Cash, J.N.; A Rejon, C.; McPherron, A.C.; Bernard, D.J.; Thompson, T.B. The structure of myostatin:follistatin 288: Insights into receptor utilization and heparin binding. EMBO J. 2009, 28, 2662–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benet, L.Z.; Hosey, C.M.; Ursu, O.; Oprea, T.I. BDDCS, the Rule of 5 and drugability. Adv. Drug Deliv. Rev. 2016, 101, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Molecule Name | rBonds | MW (D) | LogP | LogS | H-Acceptors | H-Donors | Druglikeness | DrugScore |
|---|---|---|---|---|---|---|---|---|---|
| 1. | ZINC85542646 | 6 | 743.105 | 9.8211 | −11.256 | 4 | 4 | 2.1393 | 0.0482794 |
| 2. | ZINC85569060 | 14 | 742.93 | 7.2932 | −9.232 | 8 | 7 | 1.1649 | 0.02769731 |
| 3. | ZINC85542795 | 8 | 735.126 | 9.3453 | −9.439 | 4 | 4 | 3.5148 | 0.1333178 |
| 4. | ZINC85625736 | 13 | 732.871 | 8.4908 | −9.179 | 10 | 6 | −3.2413 | 0.025806 |
| 5. | ZINC85542639 | 6 | 715.051 | 9.1759 | −10.588 | 4 | 4 | 2.7851 | 0.05082912 |
| 6. | ZINC85542627 | 6 | 717.067 | 9.5135 | −10.833 | 4 | 4 | 2.1393 | 0.04927217 |
| 7. | ZINC85531289 | 6 | 720.856 | 8.5678 | −8.717 | 9 | 1 | −5.2949 | 0.02028264 |
| 8. | ZINC85542877 | 10 | 723.115 | 9.6422 | −9.433 | 4 | 4 | 1.6451 | 0.1248953 |
| 9. | ZINC85542671 | 6 | 720.071 | 7.3836 | −8.911 | 5 | 5 | 4.507 | 0.1466092 |
| 10. | ZINC85569094 | 14 | 720.924 | 7.7686 | −8.987 | 8 | 7 | −0.80112 | 0.03387373 |
| 11. | ZINC85532197 | 3 | 743.063 | 8.2324 | −9.457 | 6 | 2 | 1.6002 | 0.1263831 |
| 12. | ZINC85532197_01 | 3 | 742.055 | 8.2324 | −9.457 | 6 | 1 | 1.6002 | 0.1264543 |
| 13. | ZINC85511481 | 14 | 710.817 | 6.4144 | −8.805 | 10 | 6 | −3.0206 | 0.05147698 |
| 14. | ZINC85569082 | 14 | 692.87 | 7.0778 | −8.648 | 8 | 7 | −0.63016 | 0.03765389 |
| 15. | ZINC85542801 | 6 | 699.093 | 8.6257 | −9.107 | 4 | 4 | 1.8161 | 0.1313846 |
| 16. | ZINC85542734 | 7 | 693.045 | 8.4862 | −9.062 | 4 | 4 | 2.1393 | 0.1351283 |
| 17. | ZINC85596043 | 8 | 686.87 | 8.4597 | −9.448 | 8 | 4 | −5.2779 | 0.0430603 |
| 18. | ZINC85531399 | 6 | 678.819 | 8.8366 | −7.533 | 8 | 1 | −4.778 | 0.02747577 |
| 19. | ZINC85542810 | 6 | 685.066 | 8.2215 | −8.82 | 4 | 4 | 1.5833 | 0.1331636 |
| 20. | ZINC85592913 | 2 | 678.819 | 7.9773 | −8.865 | 8 | 4 | −4.767 | 0.07450064 |
| 21. | ZINC85531346 | 3 | 676.803 | 7.8245 | −7.602 | 8 | 1 | −7.5213 | 0.02813334 |
| 22. | ZINC85542876 | 6 | 673.055 | 8.3219 | −8.832 | 4 | 4 | 1.8161 | 0.1365408 |
| 23. | ZINC95911591 | 1 | 656.816 | 9.9572 | −11.412 | 6 | 1 | −3.9286 | 0.0727493 |
| 24. | ZINC85542793 | 6 | 671.039 | 8.0144 | −8.677 | 4 | 4 | 1.5283 | 0.1360255 |
| 25. | ZINC85542803 | 6 | 671.039 | 7.9522 | −8.66 | 4 | 4 | 1.3666 | 0.134584 |
| 26. | ZINC85592908 | 12 | 664.792 | 7.6353 | −8.595 | 8 | 4 | −2.4411 | 0.08293768 |
| 27. | ZINC85531409 | 5 | 650.765 | 7.8019 | −6.708 | 8 | 1 | −5.4649 | 0.03143037 |
| 28. | ZINC85530919 | 7 | 644.718 | 8.5529 | −7.639 | 8 | 4 | −1.1148 | 0.0982419 |
| 29. | ZINC85542903 | 6 | 645.001 | 7.7106 | −8.402 | 4 | 4 | 1.5283 | 0.1435007 |
| 30. | ZINC85542935 | 6 | 645.001 | 7.6484 | −8.385 | 4 | 4 | 1.3666 | 0.1421086 |
| 31. | ZINC85542917 | 6 | 645.001 | 7.6484 | −8.385 | 4 | 4 | 1.5283 | 0.14408 |
| 32. | ZINC85592903 | 10 | 636.739 | 7.0594 | −7.91 | 8 | 4 | −5.2216 | 0.08549763 |
| 33. | ZINC85542926 | 5 | 616.947 | 7.2631 | −7.795 | 4 | 4 | 3.5148 | 0.1706919 |
| 34. | ZINC85531359 | 7 | 620.693 | 5.5821 | −5.48 | 10 | 2 | −9.9673 | 0.04023146 |
| 35. | ZINC85543487 | 1 | 629.007 | 9.673 | −9.675 | 2 | 2 | −0.43731 | 0.1041854 |
| 36. | ZINC85949541 | 2 | 592.69 | 6.3122 | −6.34 | 8 | 0 | 1.9873 | 0.1715774 |
| 37. | ZINC70454202_01 | 4 | 593.742 | 6.3699 | −6.12 | 7 | 3 | 4.6261 | 0.2320921 |
| 38. | ZINC70454202 | 4 | 594.749 | 6.3699 | −6.12 | 7 | 4 | 4.6261 | 0.2316766 |
| 39. | ZINC85543478 | 1 | 616.996 | 9.3076 | −9.498 | 2 | 2 | −0.52971 | 0.1050597 |
| 40. | ZINC85541065 | 3 | 576.691 | 7.2035 | −8.002 | 7 | 1 | 4.6261 | 0.147236 |
| 41. | ZINC85531053 | 3 | 576.647 | 6.7914 | −10.044 | 8 | 1 | 1.741 | 0.1367792 |
| 42. | ZINC42802834 | 2 | 562.664 | 7.0099 | −8.195 | 7 | 1 | 4.6261 | 0.1527083 |
| 43. | ZINC85541288 | 2 | 562.664 | 7.0327 | −8.871 | 7 | 1 | 4.8552 | 0.1497575 |
| 44. | ZINC95910145 | 2 | 548.637 | 6.757 | −8.557 | 7 | 2 | 4.8369 | 0.1594868 |
| 45. | ZINC44086846 | 2 | 546.621 | 6.8386 | −9.35 | 7 | 1 | 4.9691 | 0.1565135 |
| 46. | ZINC85991498_01 | 1 | 548.593 | 5.5895 | −6.867 | 8 | 2 | −1.7726 | 0.1196459 |
| 47. | ZINC85991498 | 2 | 548.593 | 5.5895 | −6.867 | 8 | 2 | −1.7726 | 0.1196459 |
| 48. | ZINC03780340 | 6 | 504.449 | 5.9594 | −10.586 | 8 | 6 | −1.1275 | 0.07115073 |
| 49. | ZINC14680812 | 6 | 512.513 | 2.823 | −5.007 | 8 | 7 | 0.51052 | 0.2703446 |
| 50. | ZINC85596478 | 3 | 482.618 | 7.2172 | −7.679 | 4 | 1 | −6.4955 | 0.04087312 |
| 51. | ZINC85947357_01 | 3 | 525.814 | 8.162 | −9.5 | 4 | 4 | −2.6575 | 0.04786098 |
| 52. | ZINC04098631 | 3 | 440.494 | 6.1569 | −7.512 | 5 | 3 | −3.1957 | 0.1446293 |
| 53. | Curcumin | 8 | 368.384 | 2.039 | −3.622 | 6 | 2 | −4.7745 | 0.391063 |
| S. No. | Molecule | BBB Permeant | PAINS | WLOGP | TPSA | Log S | Skin Permeability | CYP2D6 Inhibitor | Carcinogenicity |
|---|---|---|---|---|---|---|---|---|---|
| 1. | ZINC85542795 | No | 0 | 8.79 | 72.72 | −12.48 | −2.98 | No | None |
| 2. | ZINC85531289 | No | 0 | 7.4 | 121.5 | −10.37 | −5.04 | No | None |
| 3. | ZINC85542877 | No | 0 | 8.63 | 72.72 | −12.42 | −2.95 | No | None |
| 4. | ZINC85542671 | No | 0 | 7.73 | 84.75 | −10.87 | −4.17 | No | None |
| 5. | ZINC85541288 | No | 0 | 5.5 | 61.42 | −6.92 | −5.57 | No | None |
| 6. | ZINC85532197 | No | 0 | 8.31 | 127.62 | −10.4 | −5.24 | No | None |
| 7. | ZINC85511481 | No | 0 | 5.91 | 173.98 | −9.86 | −6.08 | No | None |
| 8. | ZINC85592908 | No | 0 | 7.13 | 117.84 | −9.54 | −5.21 | No | None |
| 9. | ZINC14680812 | No | 0 | 3.69 | 158.68 | −4.19 | −8.53 | No | None |
| 10. | ZINC85592903 | No | 0 | 6.14 | 117.84 | −8.63 | −5.67 | No | None |
| 11. | ZINC85531359 | No | 0 | 4.29 | 141.73 | −6.83 | −7.14 | No | None |
| 12. | ZINC70454202 | No | 0 | 4.59 | 92.21 | −8.13 | −5.38 | No | None |
| 13. | ZINC95910145 | No | 0 | 5.2 | 72.42 | −6.82 | −5.71 | No | None |
| 14. | ZINC44086846 | No | 0 | 5.68 | 72.75 | −6.74 | −5.76 | No | None |
| 15. | ZINC04098631 | No | 0 | 6.27 | 79.15 | −7.88 | −4.43 | No | None |
| 16. | ZINC85991498 | No | 0 | 5.39 | 109.94 | −6.7 | −6.34 | No | None |
| 17. | Curcumin | No | 0 | 3.15 | 93.06 | −4.83 | −6.28 | No | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.; Ahmad, K.; Shaikh, S.; Lim, J.H.; Chun, H.J.; Ahmad, S.S.; Lee, E.J.; Choi, I. Identification and Evaluation of Traditional Chinese Medicine Natural Compounds as Potential Myostatin Inhibitors: An In Silico Approach. Molecules 2022, 27, 4303. https://doi.org/10.3390/molecules27134303
Ali S, Ahmad K, Shaikh S, Lim JH, Chun HJ, Ahmad SS, Lee EJ, Choi I. Identification and Evaluation of Traditional Chinese Medicine Natural Compounds as Potential Myostatin Inhibitors: An In Silico Approach. Molecules. 2022; 27(13):4303. https://doi.org/10.3390/molecules27134303
Chicago/Turabian StyleAli, Shahid, Khurshid Ahmad, Sibhghatulla Shaikh, Jeong Ho Lim, Hee Jin Chun, Syed Sayeed Ahmad, Eun Ju Lee, and Inho Choi. 2022. "Identification and Evaluation of Traditional Chinese Medicine Natural Compounds as Potential Myostatin Inhibitors: An In Silico Approach" Molecules 27, no. 13: 4303. https://doi.org/10.3390/molecules27134303