
Natural Products Inhibitors of Monoamine Oxidases—Potential New Drug Leads for Neuroprotection, Neurological Disorders, and Neuroblastoma




Abstract
:1. Introduction

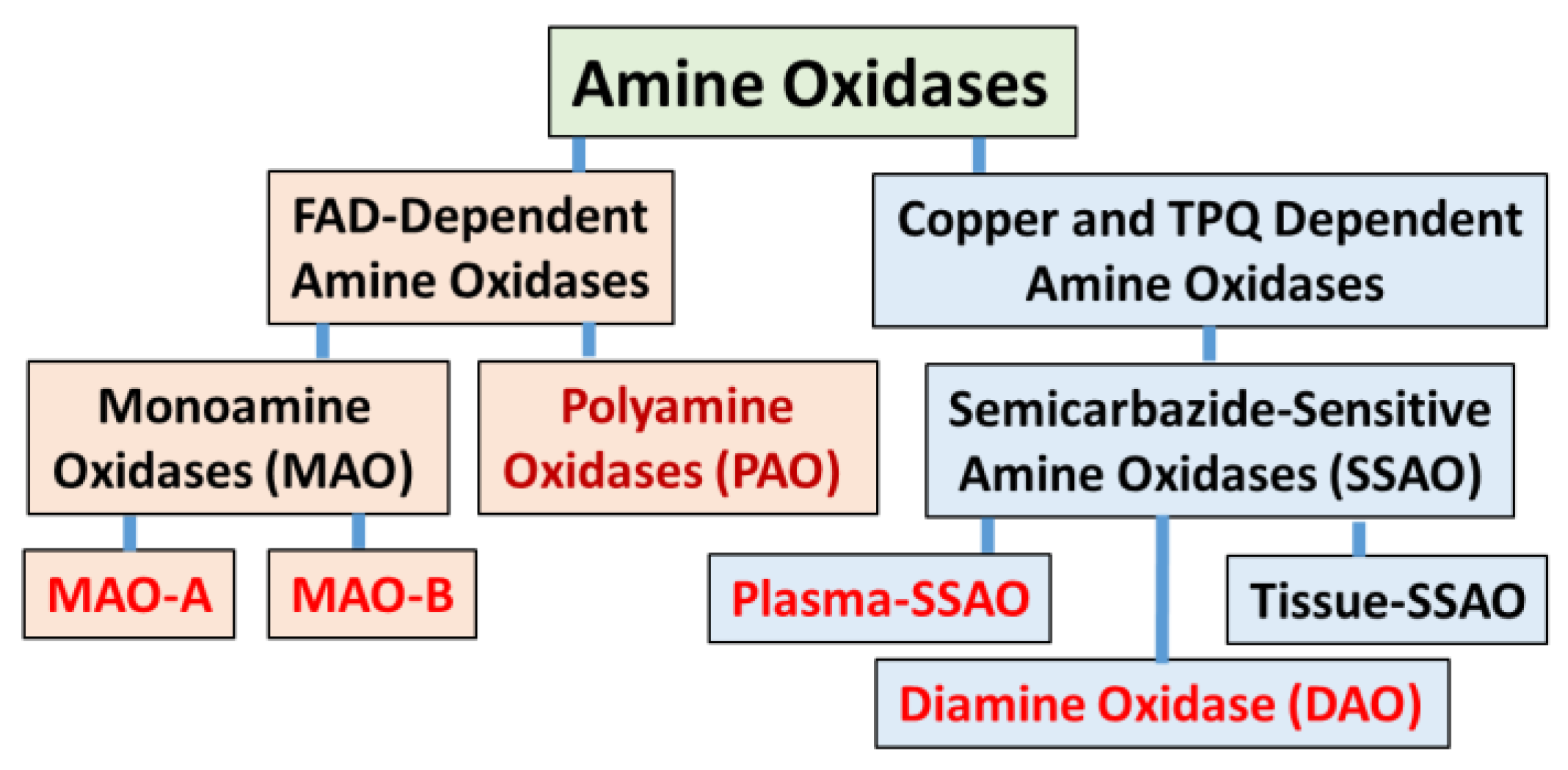
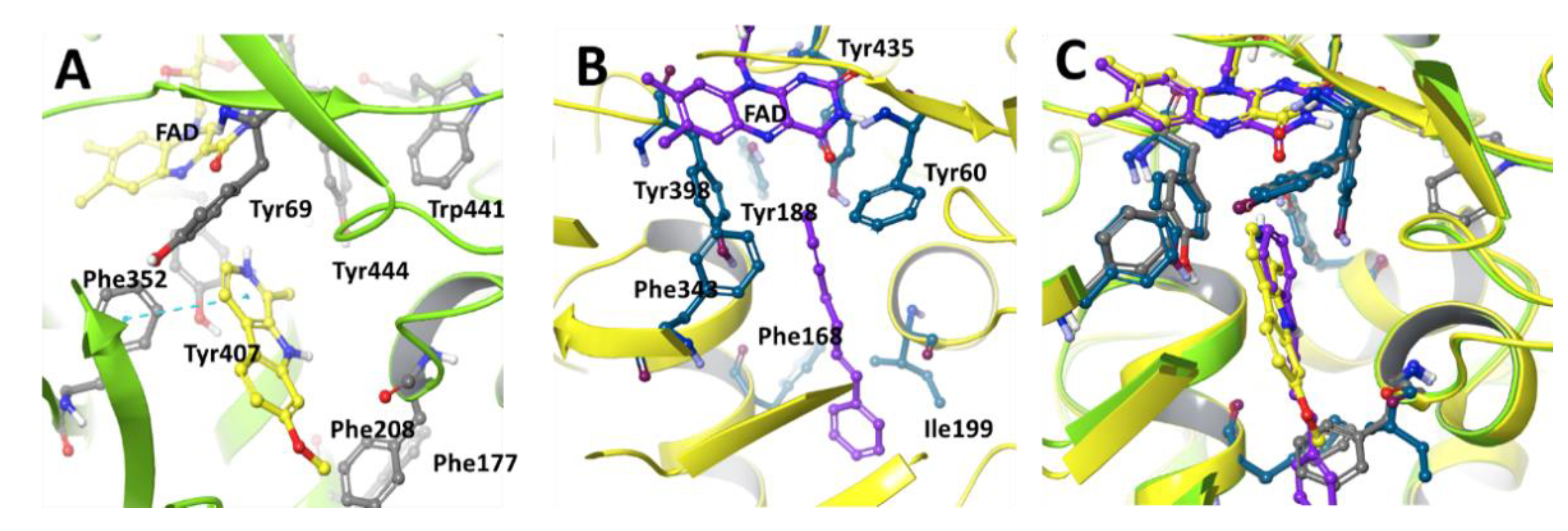
2. Structural Difference of MAO-A and MAO-B
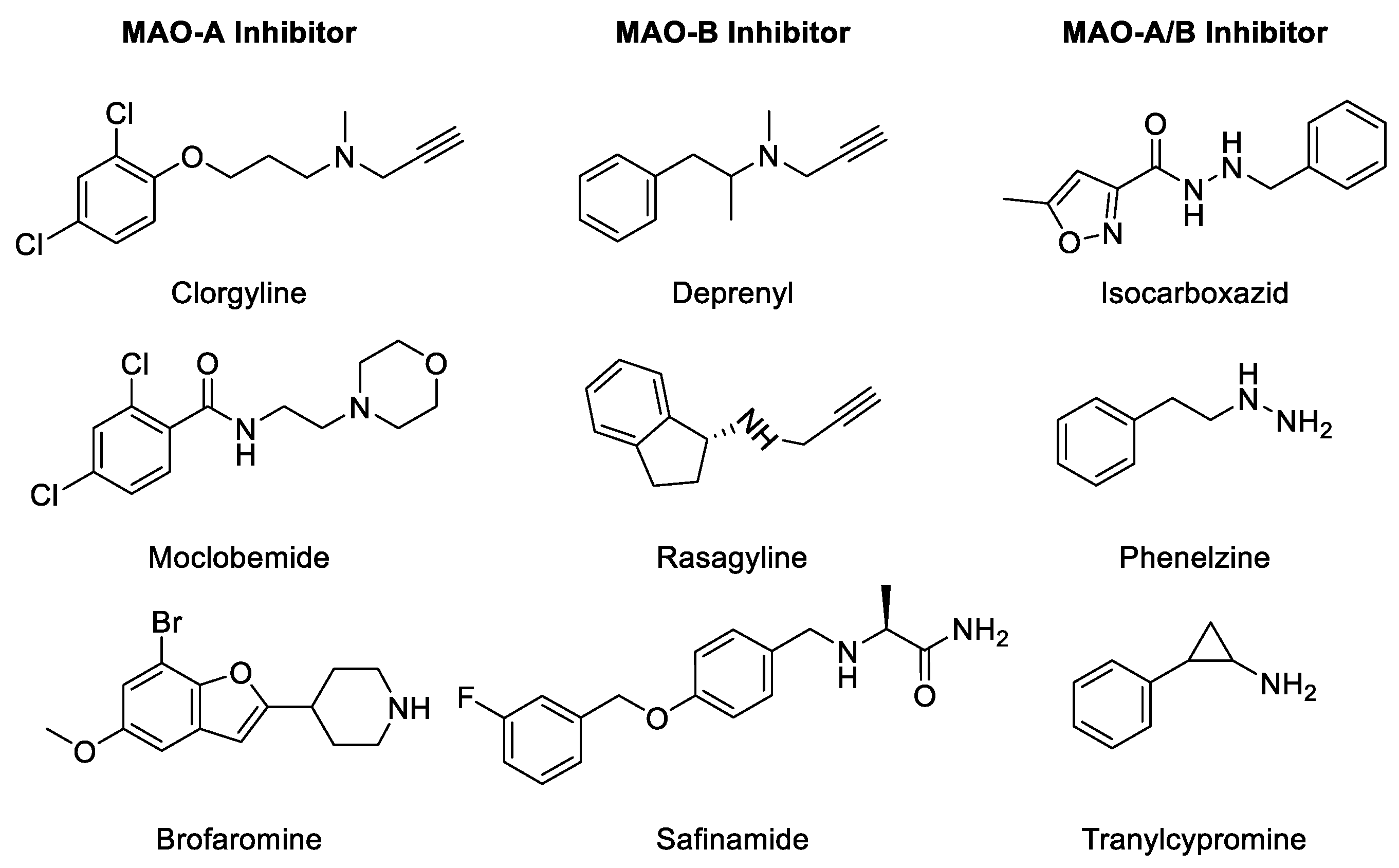
2.1. MAO Inhibitors (MAOIs)
2.2. Therapeutic Applications of MAO Inhibitors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor Type | MAO-A | MAO-B | Non-Selective |
|---|---|---|---|
| Irreversible inhibitors | Clorgyline; Lilly 51641 | (-) Deprenyl Lilly 54781 MDL 72145 AGN 1133 AFN 1135 Rasagiline Pargyline Selegiline | Phenelzine Tranylcypromine Isocarboxazid Nialamide Iproniazid Safrazine Metfendrazine |
| Reversible inhibitors | Harmaline Amiflamine Cimoxatone Moclobemide Brofaromine Ro 11-1163 Toloxatone MD 780515 FLA 336(+) | Safinamide |
3. MAO Inhibitors in Clinical Development
Limitations of Currently Approved MAOIs and Current Approach for Development of New MAOIs
4. Traditional and Psychoactive Medicinal Plants and Herbal Formulations for Treatment of Neurological Disorders
5. Different Classes of Natural Product MAOIs
5.1. Alkaloids
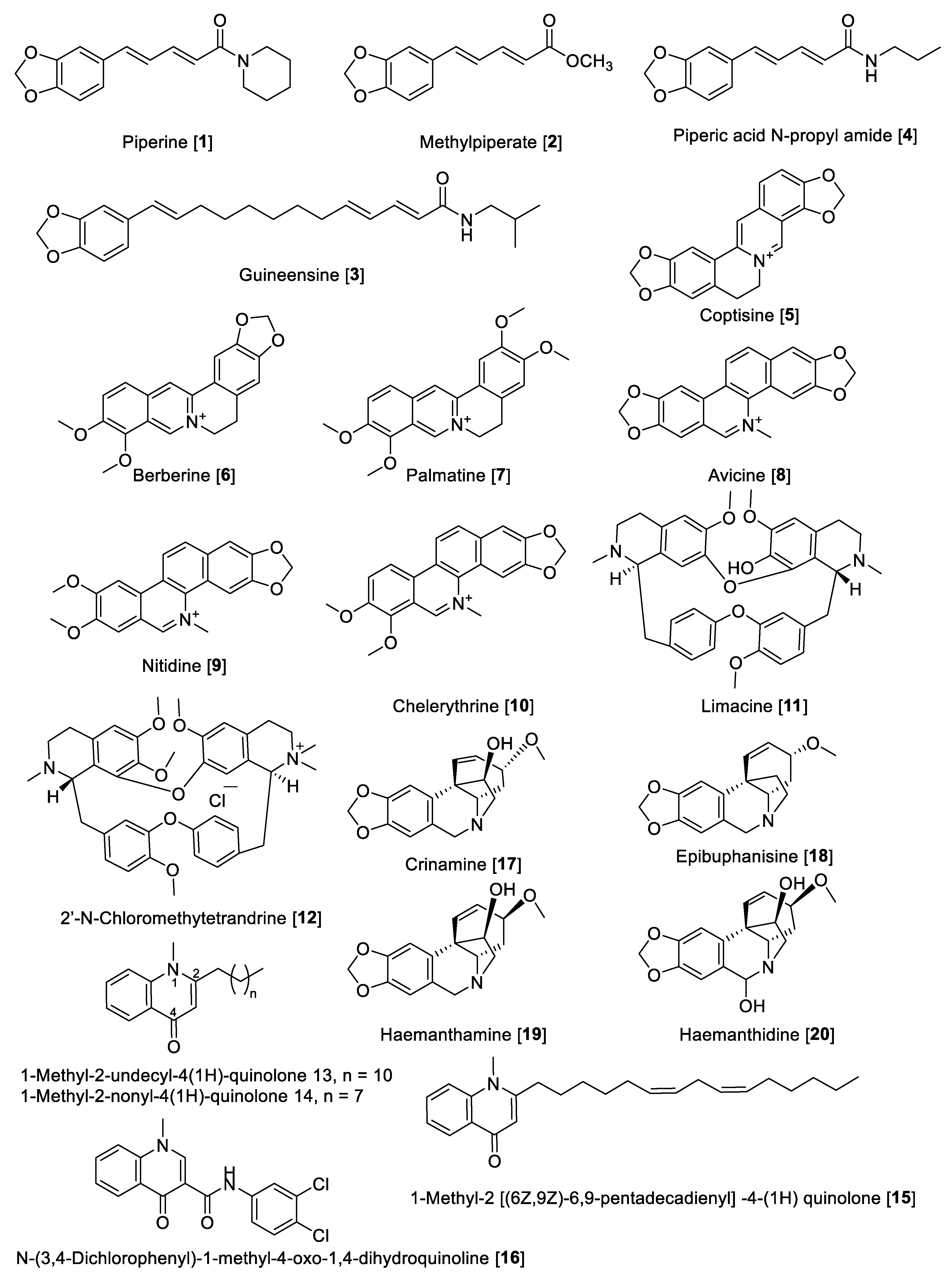
5.1.1. MAO Inhibitory Activity of Piperine, Quinolone, and Isoquinoline Alkaloids

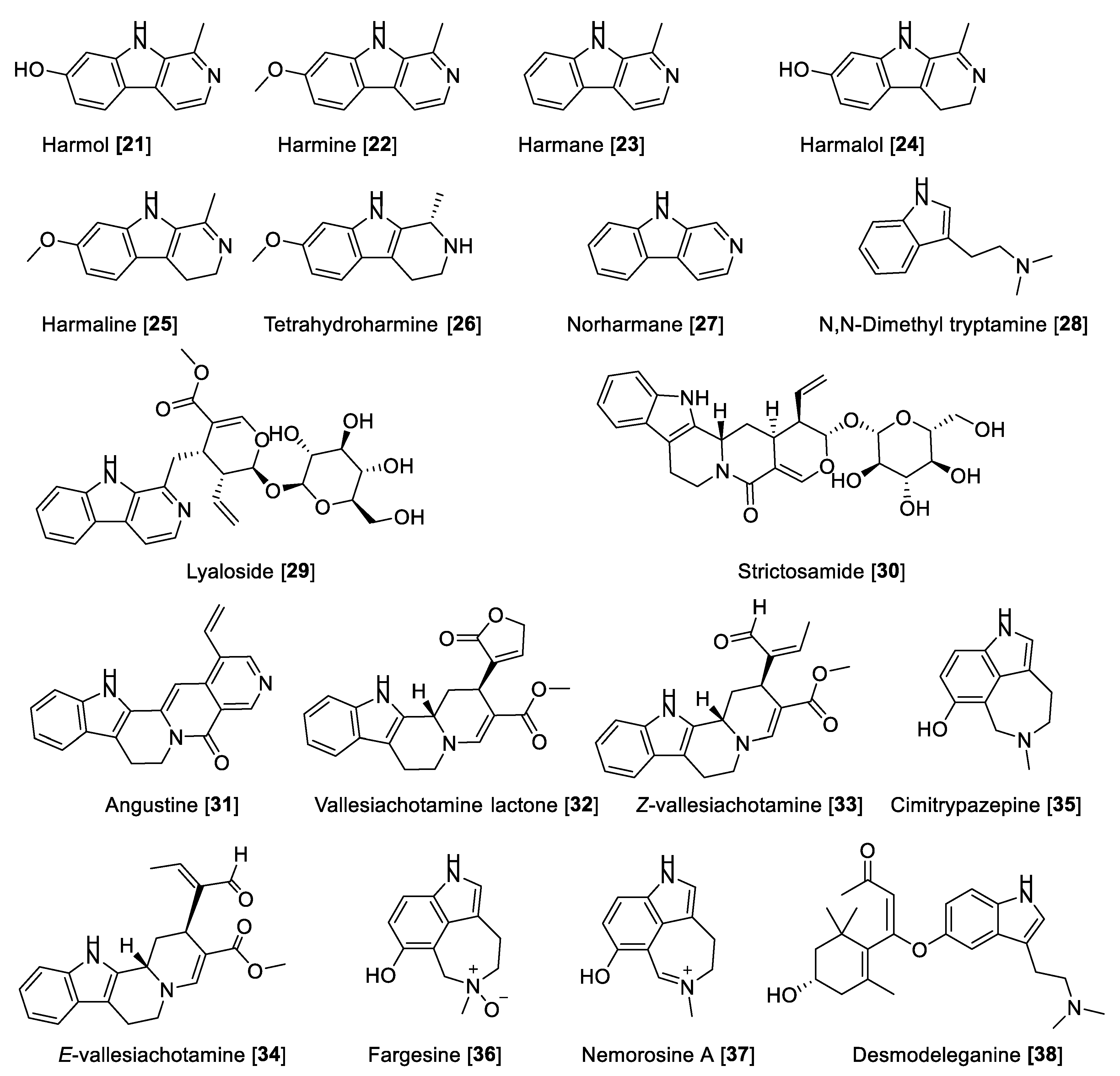
5.1.2. MAO Inhibitory Activity of Beta-Carbolines and Indole-Type Alkaloids

| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Piperine [1] | Piper longum | 49.3 | 35.8 | 91.3 | 79.9 | 0.54 | A | [68] |
| Piper longum | 20.9 | 19.0 | 7.0 | 3.19 | 2.98 | A | [69] | |
| Methylpiperate [2] | Piper longum | 27.1 | 23.5 | 1.6 | 1.3 | 16.93 | A | [70] |
| piperic acid N-propyl amide [4] | Piperine derivative | 3.66 | 0.045 | 81.3 | B | [71] | ||
| Coptisine [5] | Coptis japonica | 1.8 | 3.3 | A | [72] | |||
| Avicine [8] | Zanthoxylum rigidum | 0.41 | >100 | >0.0041 | F | [76] | ||
| Nitidine [9] | Zanthoxylum rigidum | 1.89 | >300 | >0.0063 | F | [76] | ||
| Chelerythrin [10] | Zanthoxylum rigidum | 0.55 | >20 | >0.0275 | F | [76] | ||
| 1-Methyl-2-undecyl-4-(1H) quinolone [13] | Evodia rutaecarpa | 338.2 | 15.3 | 9.91 | 22.10 | A | [78] | |
| 1-Methyl-2-nonyl-4 (1H)-quinolone [14] | Evodia rutaecarpa | 240.2 | 2.3 | 104.4 | A | [80] | ||
| 1-Methyl-2 [(6Z,9Z)-6,9-pentadecadienyl] -4-(1H) quinolone [15] | Evodia rutaecarpa | >400 | 3.6 | 3.8 | 111.1 | A | [80] | |
| Quinoline derivative [16] | Quinolone derivative | >100 | 0.0053 | >18,867 | F | [81] | ||
| Crinamine [17] | Crossyne guttata | 0.014 | F | [82] | ||||
| Epibuphanisine [18] | Crossyne guttata | 0.039 | F | [82] | ||||
| Haemanthamine [19] | Scadoxus puniceus | 0.112 | F | [82] | ||||
| Haemanthidine [20] | Scadoxus puniceus | 0.017 | F | [82] | ||||
| Harmol [21] | Banisteriopsis caapi | 0.018 | F | [61] | ||||
| Banisteriopsis caapi | 0.5 | F | [84] | |||||
| Peganum harmala | 0.352 | F | [86] | |||||
| Harmine [22] | Banisteriopsis caapi | 0.002 | 20 | 0.0001 | F | [61] | ||
| Diverse vendors | 0.06 | NR | F | [84] | ||||
| Peganum harmala | 0.008 | NR | F | [86] | ||||
| Banisteriopsis caapi | 0.004 | >10 | >0.0004 | A | [89] | |||
| Harmane [23] | Banisteriopsis caapi | 0.64 | NR | F | [84] | |||
| Harmalol [24] | Banisteriopsis caapi | 0.66 | NR | F | [84] | |||
| Peganum harmala | 0.48 | NR | F | [86] | ||||
| Harmaline [25] | Banisteriopsis caapi | 0.002 | 25 | 0.00008 | F | [61] | ||
| Diverse vendors | 0.09 | NR | F | [84] | ||||
| Peganum harmala | 0.012 | NR | F | [86] | ||||
| Tetrahydro harmine [26] | Diverse vendors | 1.52 | NR | F | [84] | |||
| Norharmane [27] | Diverse vendors | 4.29 | NR | F | [84] | |||
| Lyaloside [29] | Psychotria. Laciniata | 182 | >100 | F | [93] | |||
| Strictosamide [30] | Psychotria laciniata | 141 | >100 | F | [93] | |||
| Angustine [31] | Psychotria laciniata | 1.10 | 138 | 0.0079 | F | [93] | ||
| Vallesiachotamine lactone [32] | Psychotria laciniata | 0.87 | 34 | 0.025 | F | [93] | ||
| E-vallesiachotamine [33] | Psychotria laciniata | 2.14 | 120 | 0.017 | F | [93] | ||
| Z-vallesiachotamine [34] | Psychotria laciniata | 0.85 | 126 | 0.0067 | F | [93] | ||
| Cimitrypazepine [35] | Psychotria nemorosa | 22 | 1.4 | 15.71 | F | [94] | ||
| Fargesine [36] | Psychotria nemorosa | 27 | 1.4 | 19.28 | F | [94] | ||
| Nemorosine A [37] | Psychotria nemorosa | 31 | 0.9 | 34.4 | F | [94] | ||
| Desmodeleganine [38] | Desmonium elegans | 9.33 | 10.16 | 0.91 | F | [95] | ||
5.2. Flavonoids

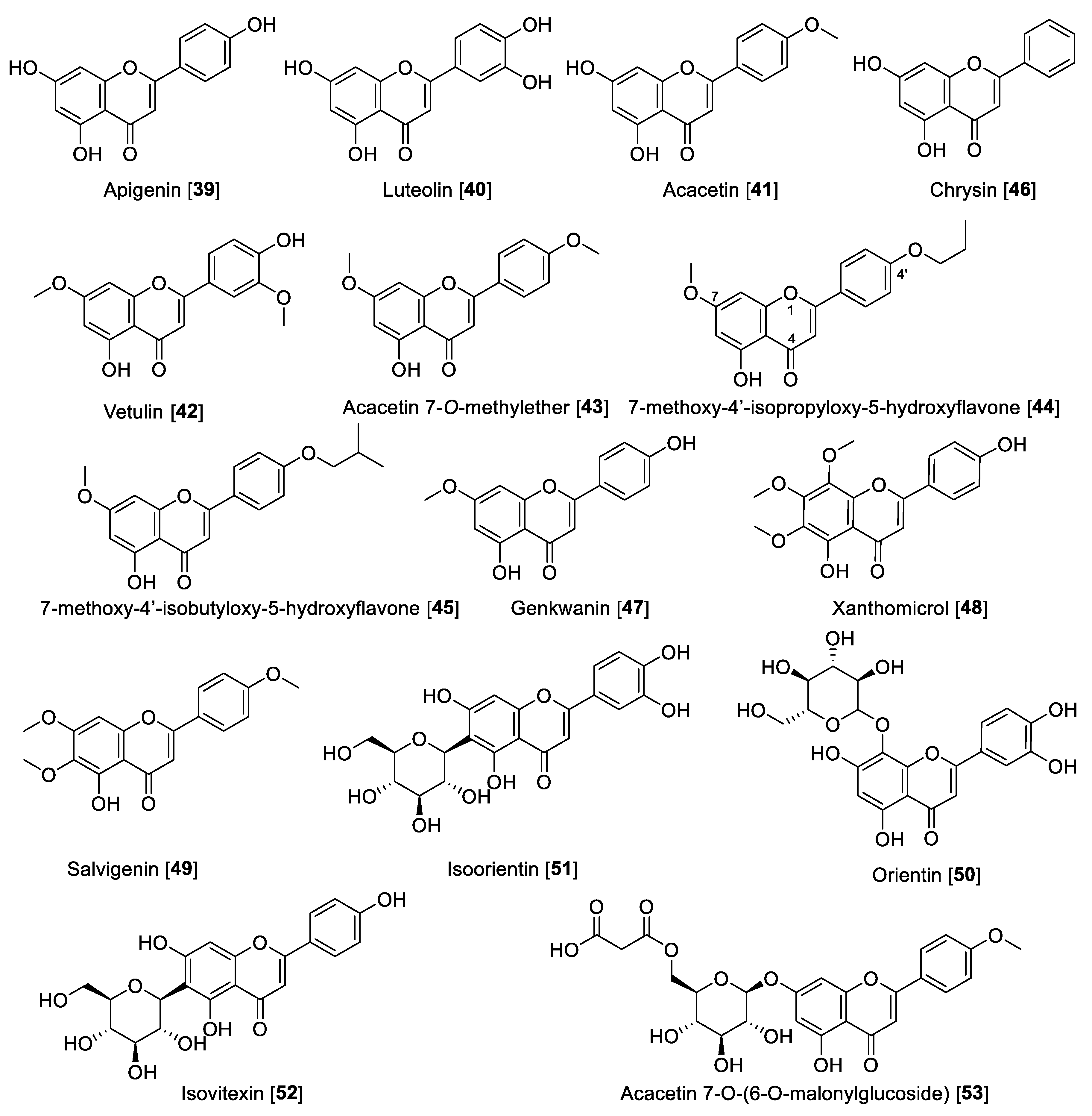
5.2.1. MAO Inhibitory Activity of Flavones
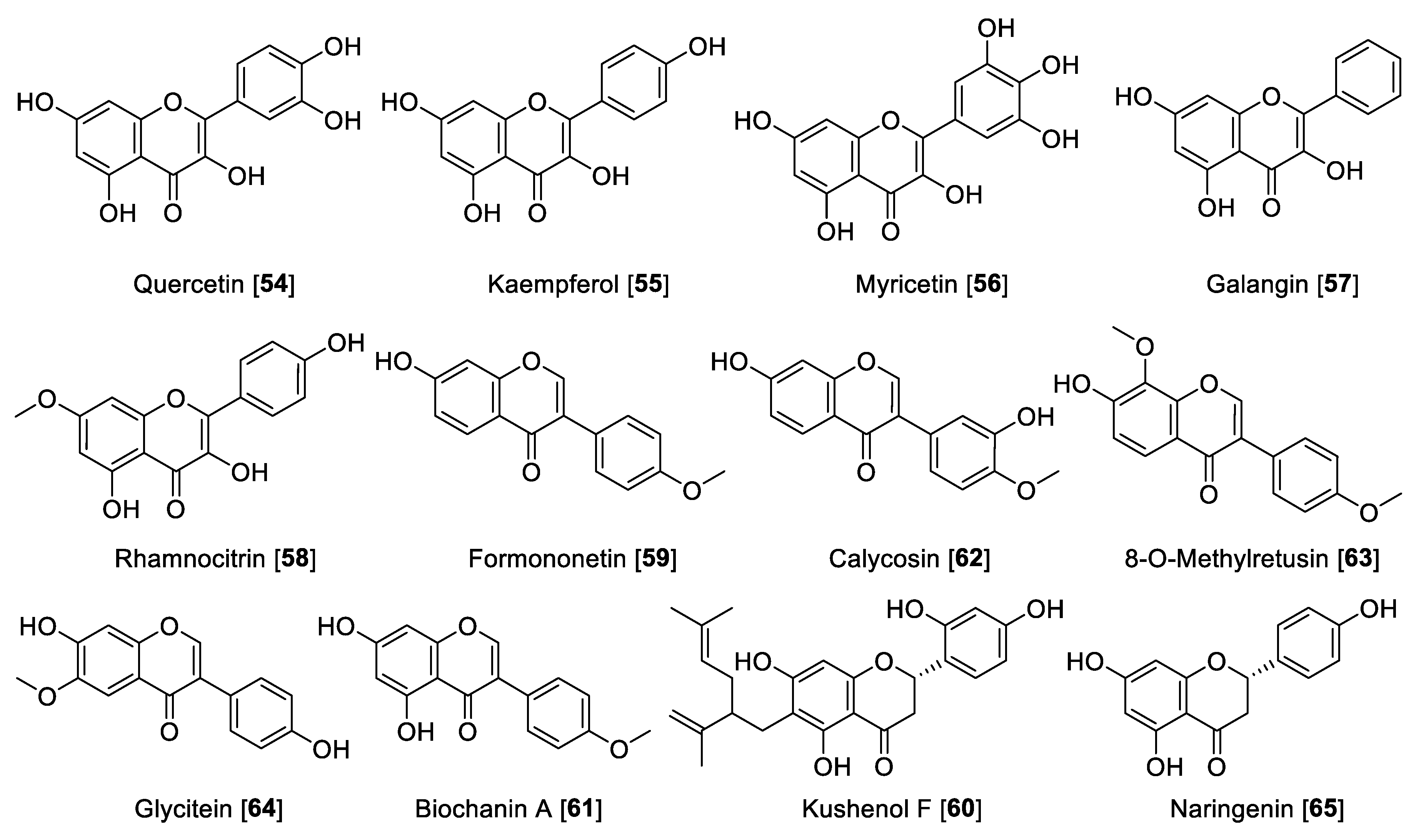
5.2.2. MAO Inhibitory Activity of Flavanols, Isoflavones, and Flavanones

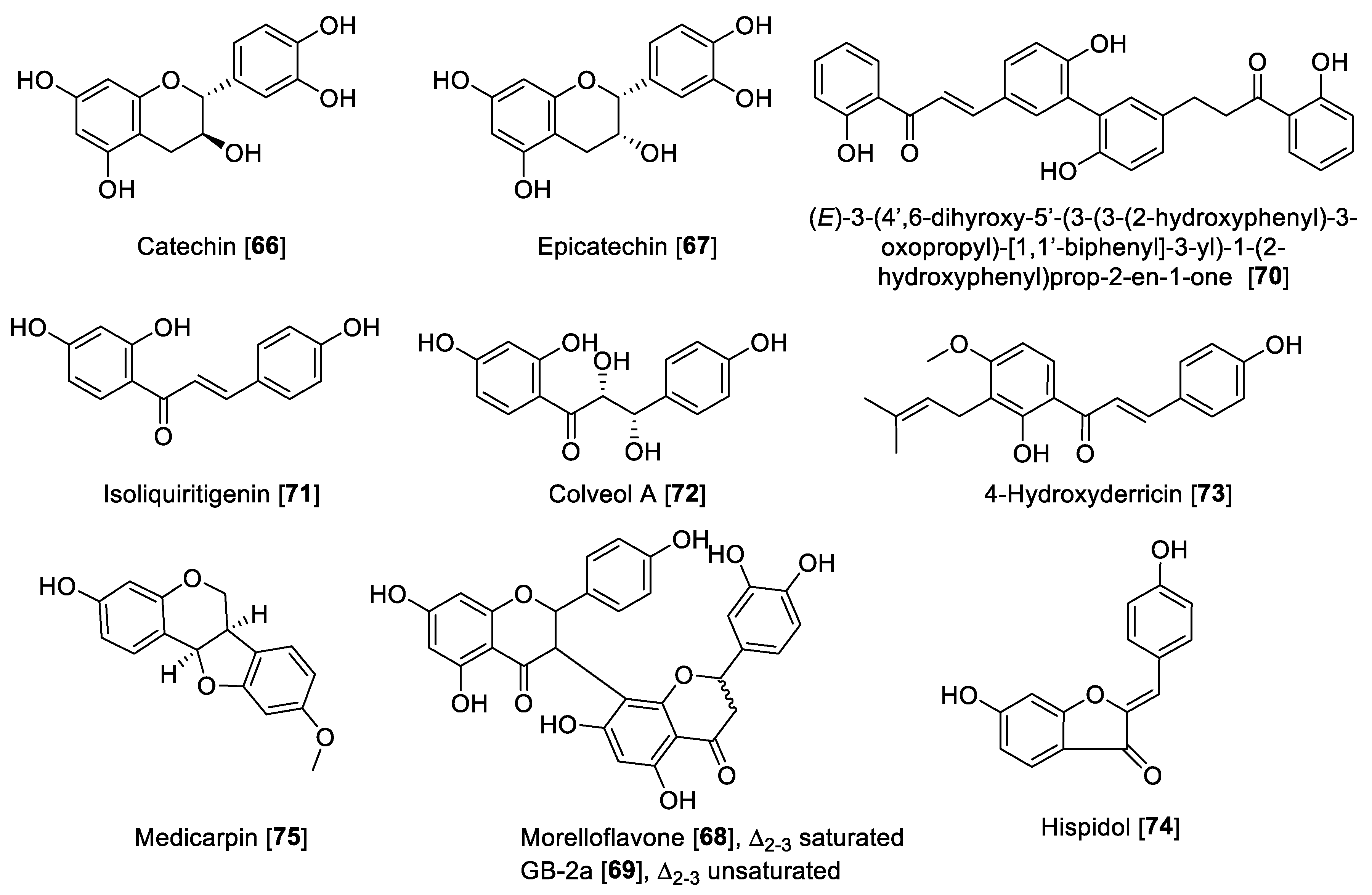
5.2.3. MAO Inhibitory Activity of Miscellaneous Flavonoids and Related Compounds
| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Apigenin [39] | Propolis | 0.64 | 0.125 | 1.12 | 0.238 | 0.57 | F | [104] |
| Luteolin [40] | Cirsium maacki | 8.57 | >100 | >0.0857 | F | [106] | ||
| Acacetin [41] | Calea urticifolia | 0.121 | 0.0592 | 0.049 | 0.049 | 2.46 | F | [107] |
| Vetulin [42] | Turnera diffusa | 18.79 | 0.447 | 42.03 | F | [108] | ||
| Acacetin 7-O-methyl ether [43] | Turnera diffusa | >100 | 0.198 | 0.045 | <505.05 | F | [108] | |
| 7-Methoxy-4′-isopropyloxy-5-hydroxyflavone [44] | Acacetin derivative | 30.74 | 0.016 | 0.052 | 1921.25 | F | [101] | |
| 7-Methoxy-4′-isobutyloxy-5-hydroxyflavone [45] | Acacetin derivative | >100 | 0.031 | 0.037 | >3225.8 | F | [101] | |
| Chrysin [46] | Cytisus villosus | 0.25 | 1.04 | NT | 0.24 | F | [109] | |
| Genkwanin [47] | Prunus padus | 0.14 | 0.097 | 0.35 | 0.12 | 0.4 | F | [109] |
| Xanthomicrol [48] | Sideritis spp | 0.76 | 99.54 | 0.0076 | F | [111] | ||
| Salvigenin [49] | Sideritis spp. | 0.54 | 6.27 | 0.086 | F | [111] | ||
| Orientin [50] | Vitex grandiflora | >100 | 11.04 | >9.05 | F | [113] | ||
| Isoorientin [51] | Vitex grandiflora | >100 | 11.08 | >9.02 | F | [113] | ||
| Isovitexin [52] | Vitex grandiflora | >100 | 21.3 | >4.69 | F | [113] | ||
| Acacetin 7-O-(6-O-malonylglucoside) [53] | Agastache rugosa | 2.34 | 1.06 | 1.87 | 0.38 | 1.25 | F | [114] |
| Quercetin [54] | Hypericum hircinum | 0.010 | 20 | 0.0005 | I | [116] | ||
| Hypericum afrum | 1.52 | 0.29 | 28.39 | 0.053 | F | [109] | ||
| Vitis vinifera | 3.98 | >100 | >0.039 | F | [117] | |||
| Kaempferol [55] | Vitis vinifera | 0.525 | >100 | >0.00525 | F | [117] | ||
| Myricetin [56] | Hypericum afrum | 9.93 | 2.24 | 59.34 | 0.167 | F | [109] | |
| Galangin [57] | Propolis | 0.13 | 0.029 | 3.65 | 1.998 | 0.035 | F | [104] |
| Rhamnocitrin [58] | Prunus padus | 0.051 | 0.097 | 2.97 | 0.12 | 0.017 | F | [110] |
| Formononetin [59] | Sophora flavescens | 21.2 | 11.0 | 1.92 | A | [119] | ||
| Maackia amurensis | 4.82 | 0.19 | 25.36 | F | [121] | |||
| Kushenol F [60] | Sophora flavescens | 103.7 | 63.1 | 1.64 | A | [119] | ||
| Biochanin [61] | Psoralea corylifolia | 3.43 | 0.099 | 0.09 | 0.0038 | 38.11 | F | [120] |
| Calycosin [62] | Maackia amurensis | 70.5 | 0.24 | 293.75 | F | [122] | ||
| 8-O-Methylretusin [63] | Maackia amurensis | 18.7 | 0.23 | 81.30 | F | [122] | ||
| Glycitein [64] | Pueraria lobata | 8.3 | 24.9 | 0.33 | F | [123] | ||
| Naringenin [65] | Colvillea racemosa | 8.64 | 0.272 | 31.76 | F | [124] | ||
| Catechin [66] | Uncaria rhynchophylla | 88.6 | 74 | B | [127] | |||
| Epicatechin [67] | Uncaria rhynchophylla | 58.9 | 21 | B | [127] | |||
| Morelloflavone [68] | Garcinia gardneriana | 5.05 | 66.2 | 0.076 | F | [127] | ||
| GB-2a [69] | Garcinia gardneriana | 5.47 | 56.7 | 0.20 | F | [127] | ||
| Bichalcone-derivative [70] | Gentiana lutea | 12.5 | 6.2 | 1.2 | 2.01 | B | [127] | |
| Isoliquiritigenin [71] | Colvillea racemosa | 22.66 | 0.51 | 44.43 | F | [124] | ||
| Colveol A [72] | Colvillea racemosa | 0.62 | 29.90 | 0.020 | F | [124] | ||
| 4-Hydroxyderricin [73] | Angelica keiskei | >3000 | 3.43 | >874.63 | F | [131] | ||
| Hispidol [74] | Glycine max | 0.26 | 0.10 | 2.45 | 0.51 | 0.10 | F | [132] |
| Medicarpin [75] | Maackia amurensis | 10.2 | 0.30 | 34.0 | F | [122] | ||
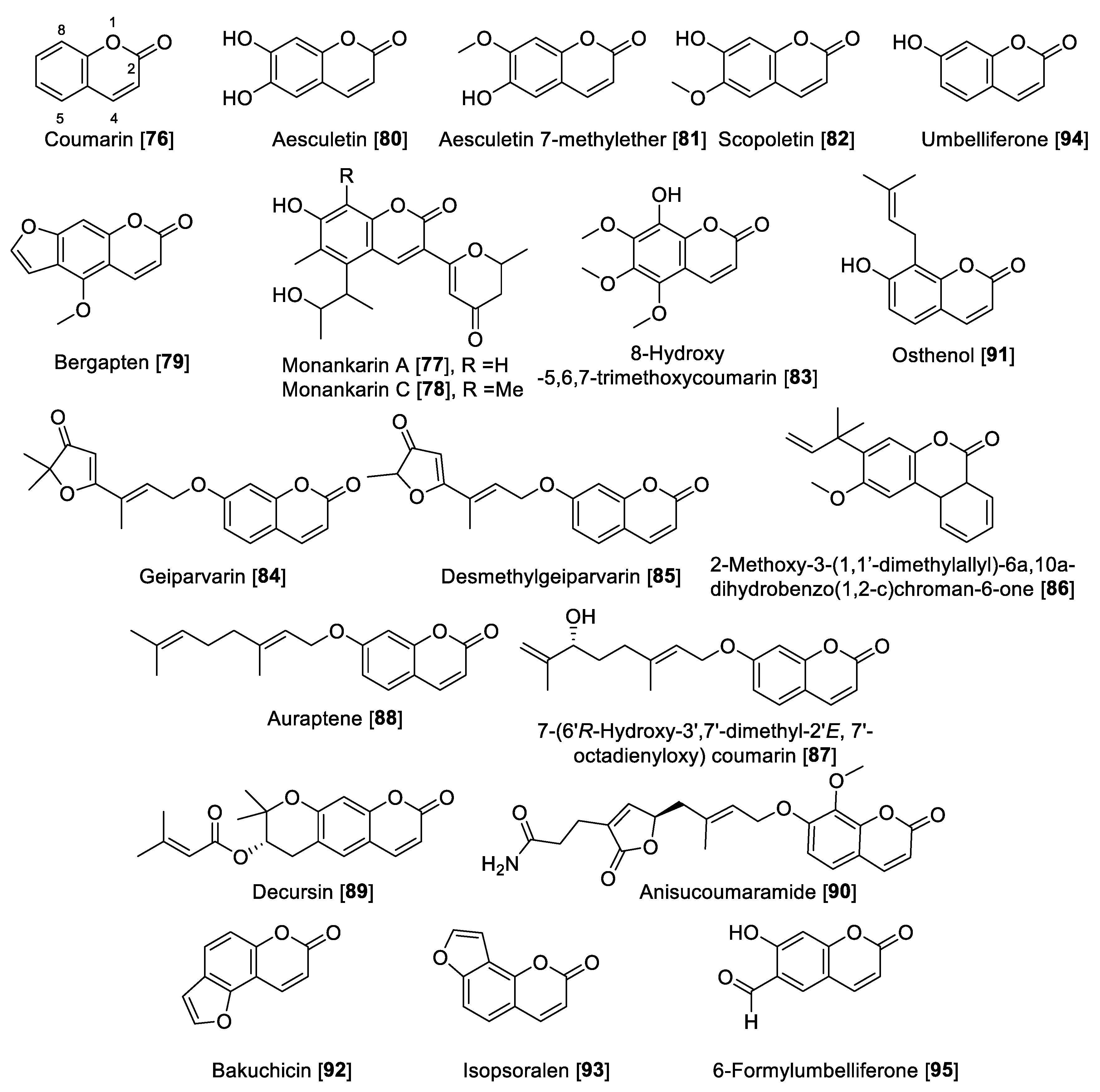
5.3. MAO Inhibitory Activity of Coumarins

| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Geiparvarin [84] | Geijera parviflora | 27 | 0.144 | 187.5 | B | [142] | ||
| Desmethylgeiparvarin [85] | Geijera parviflora | 24 | 0.028 | 857.1 | B | [142] | ||
| 2-Methoxy-3-(1,10-dimethylallyl)-6a,10a-dihydrobenzo(1,2-c)chroman-6-one [86] | Gentina lutea | >100 | -- | 2.9 | 1.1 | >34.5 | B | [129] |
| 7-(6′R-Hydroxy-3′,7′-dimethyl-2′E, 7′-octadienyloxy) coumarin [87] | Dictamnus albus | 1.3 | -- | 0.5 | 0.46 | 2.6 | A | [141] |
| Auraptene [88] | Dictamnus albus | 34.6 | -- | 0.6 | 0.83 | 57.6 | A | [141] |
| Decursin [89] | Angelica gigas | 0.6 | 70.5 | 0.0085 | F | [142] | ||
| Anisucoumaramide [90] | Clausena anisum-olens | >100 | 0.143 | 699.3 | F | [143] | ||
| Osthenol [91] | Angelica pubescens | 0.74 | 0.26 | >60 | -- | >0.012 | F | [144] |
| Bakuchicin [92] | Angelica pubescens | 1.78 | 5.44 | 0.32 | F | [144] | ||
| Isopsoralen [93] | Angelica pubescens | 0.88 | 0.46 | 2.73 | 0.32 | F | [144] | |
| Umbelliferone [94] | Angelica decursiva | 39.16 | 147.37 | 0.26 | F | [145] | ||
| 6-Formylumbelliferone [95] | Angelica decursiva | 3.23 | 3.05 | 15.31 | 6.81 | 0.21 | F | [145] |
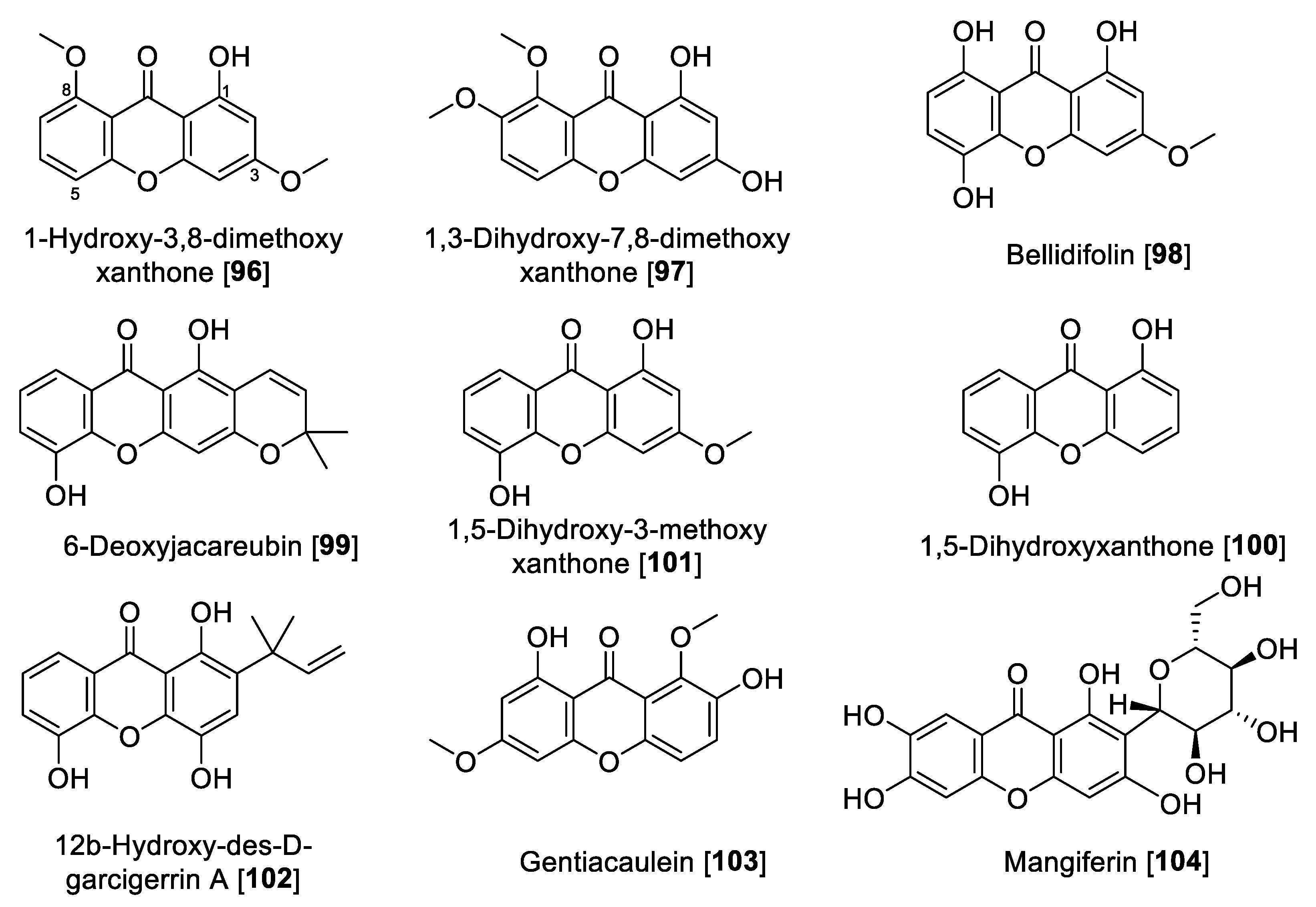
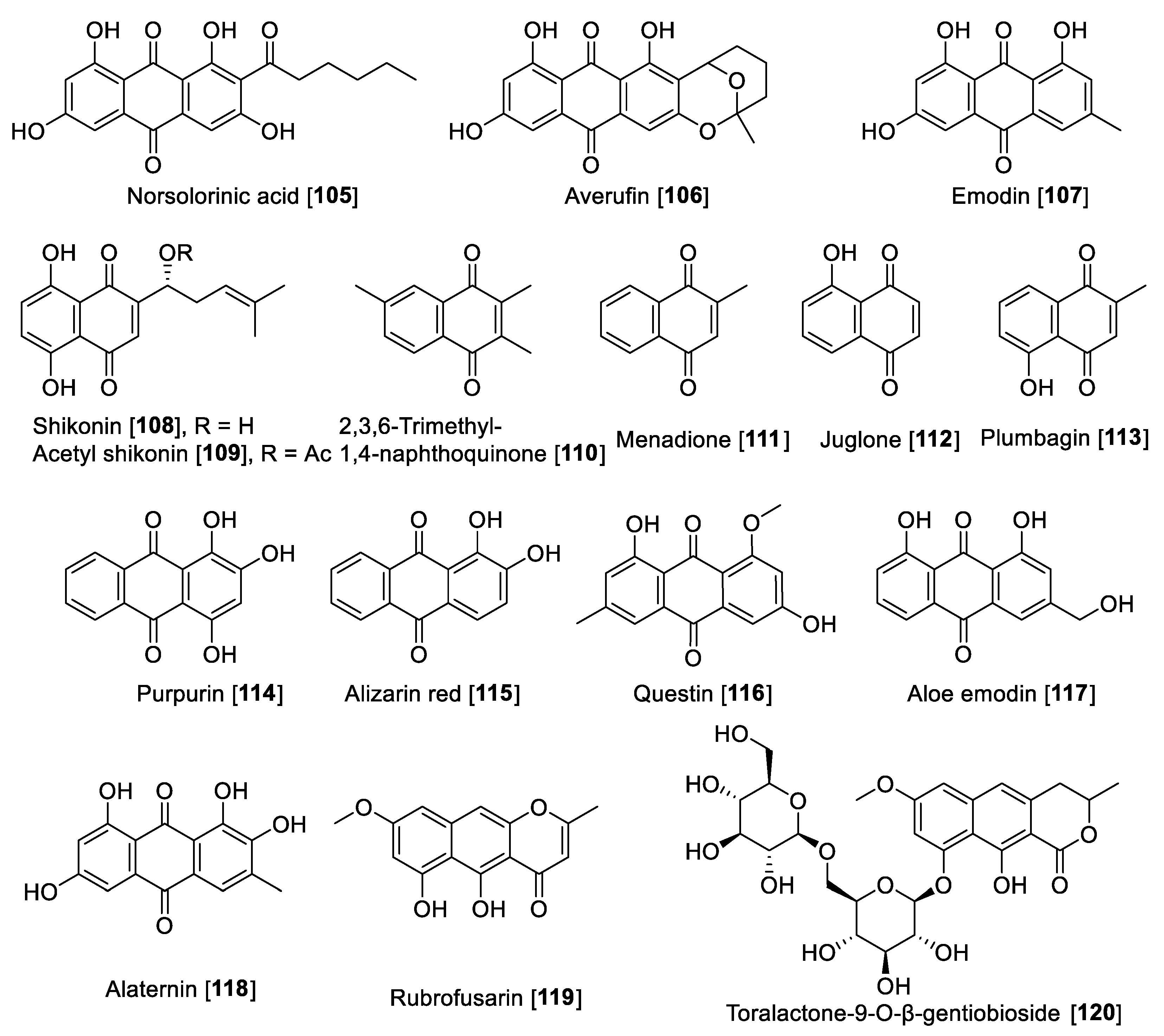
5.4. MAO Inhibitory Activity of Xanthones, Anthraquinones, and Naphthoquinones
| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Bellidolin [98] | Gentiana lactea. | 0.66 | >100 | >0.0066 | C | [157] | ||
| 6-Deoxyjacareubin [99] | Hypericum brasiliense | 12.0 | 47.3 | 0.25 | B | [156] | ||
| 1,5-Dihidroxyxanthone [100] | Hypericum brasiliense | 0.73 | 76.3 | 0.0095 | B | [156] | ||
| 1,5-dihydroxy-3-methoxy xanthone [101] | Chironia krebsii | 0.04 | 33.0 | 0.0012 | C | [157] | ||
| 12b-hydroxy-des-D-garcigerrin A [102] | Garcinia gerrardii | 3.3 | >100 | 0.033 | C | [157] | ||
| Gentiacaulein [103] | Gentiana kochiana | 0.22 | 96.0 | 0.0022 | B | [161] | ||
| Mangiferin [104] | Hypericum aucheri | 410 | >1000 | 0.41 | C | [170] | ||
| Emodin [107] | Polygonaceae Fam. | 35.4 | 15.1 | B | [68] | |||
| Shikonin [108] | Lithospermum erythrorhizon | 16.4 | 12.8 | 13.6 | 13.0 | 1.20 | A | [167] |
| Acetyl shikonin [109] | Lithospermum erythrorhizon | 16.9 | 10.5 | 10.1 | 6.3 | 1.67 | A | [167] |
| 2,3,6-Trimethyl-1,4-naphthoquinone [110] | Nicotiana tabacum | 3.0 | 6.0 | 0.5 | E | [168] | ||
| 1.14 | 7.14 | 1.59 | F | [169] | ||||
| Menadione [111] | Vit K derivative | 26.0 | 0.4 | 65 | F | [169] | ||
| 10.2 | 3.02 | 3.37 | F | [170] | ||||
| Juglone [112] | Juglans spp. | 1.71 | 4.36 | 0.39 | F | [170] | ||
| Plumbagin [113] | Plumbago spp. | 4.91 | 1.09 | 4.50 | F | [170] | ||
| Purpurin [114] | Rubia tinctorum | 2.50 | 0.422 | >40 | >0.062 | F | [172] | |
| Alizarin red [115] | Rubia tinctorum | 30.1 | >60 | >0.50 | F | [173] | ||
| Questin [116] | Cassia obtusifolia | 0.17 | 4.14 | 10.58 | 0.016 | F | [173] | |
| Aloe emodin [117] | Cassia obtusifolia | 2.47 | 0.50 | >400 | >0.0061 | F | [173] | |
| Alaternin [118] | Cassia obtusifolia | 5.35 | 3.97 | 4.55 | 1.17 | F | [173] | |
| Rubrofusarin [119] | Cassia obtusifolia | 5.90 | 4.38 | 91.40 | 0.064 | F | [173] | |
| Toralactone-9-O-β-gentiobioside [120] | Cassia obtusifolia | 7.36 | 4.30 | >400 | >0.0184 | F | [173] | |
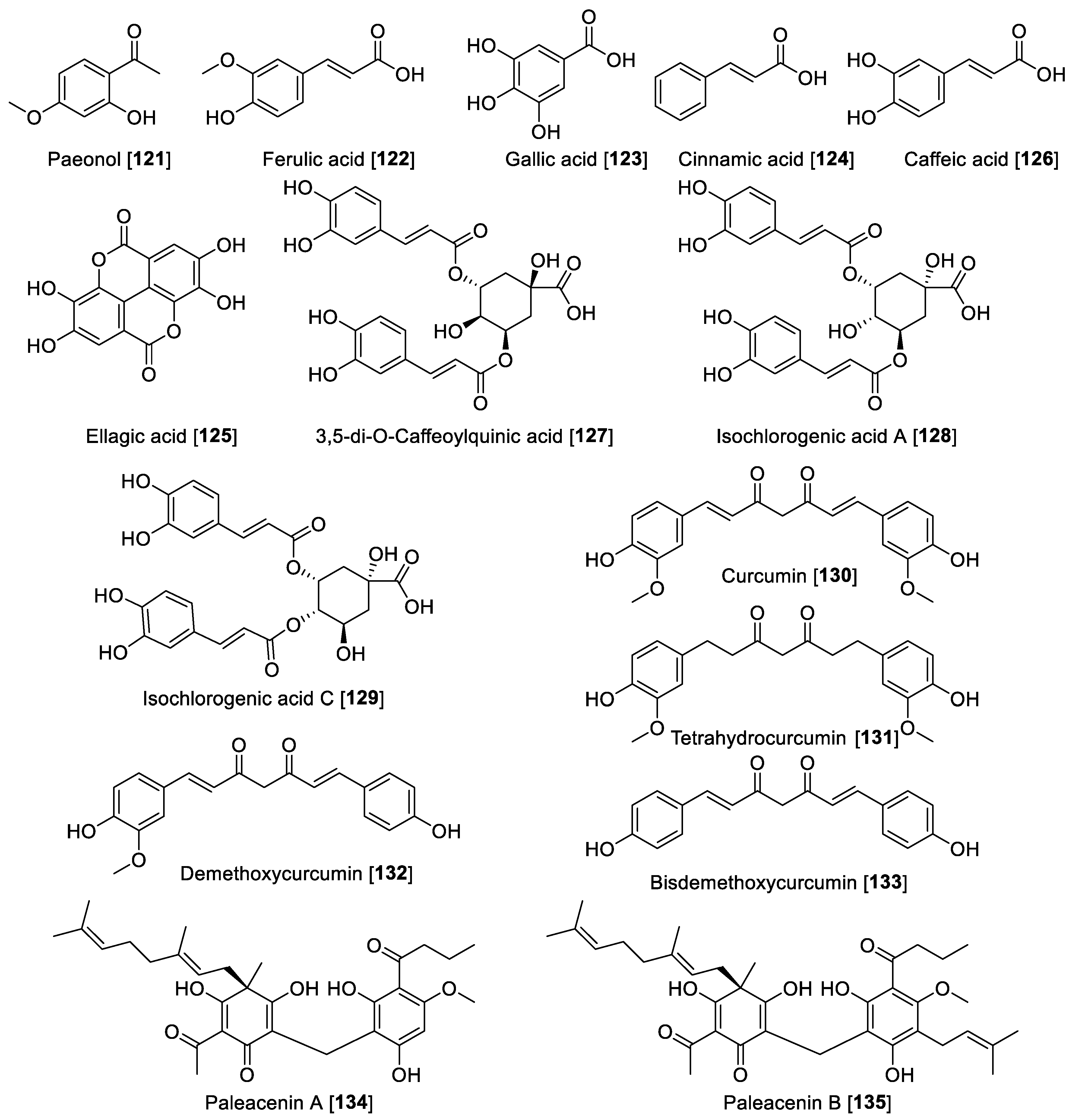
5.5. MAO Inhibitory Phenols and Polyphenolic Compounds
| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Paeonol [121] | Paeonia spp. | 54.6 | 51.1 | 42.5 | 1.28 | B | [68] | |
| Ferulic acid [122] | 7.55 | 24.0 | 0.31 | F | [175] | |||
| Gallic acid [123] | 9.49 | NR | F | [175] | ||||
| t-Cinnamic acid [124] | 6.47 | 1.21 | 5.34 | A | [175] | |||
| Ellagic acid [125] | 0.40 | B | [175] | |||||
| Caffeic acid [126] | 11.7 | 22.9 | 0.51 | F | [175] | |||
| 3,5-di-O-caffeoylquinic acid [127] | Lonicera macranthoides | 20.04 | B | [176] | ||||
| Isochlorogenic acid A [128] | Lonicera japonica | 29.05 | 9.55 | J | [177] | |||
| Isochlorogenic acid C [129] | Lonicera japonica | 29.77 | 9.53 | J | [177] | |||
| Curcumin [130] | Curcuma longa | 3.64 | 3.36 | 1.08 | F | [180] | ||
| Demethoxycurcumin [132] | Curcuma longa | 3.09 | 0.91 | 2.59 | 0.86 | 1.19 | F | [180] |
| bis-Demethoxycurcumin [133] | Curcuma longa | 3.24 | 1.40 | 2.45 | 0.80 | 1.32 | F | [180] |
| Paleacenins A [134] | Elaphoglossum paleaceum | 31.0 | 4.7 | 6.59 | B | [181] | ||
| Paleacenins C [135] | Elaphoglossum paleaceum | 1.3 | 4.4 | 0.29 | B | [181] | ||
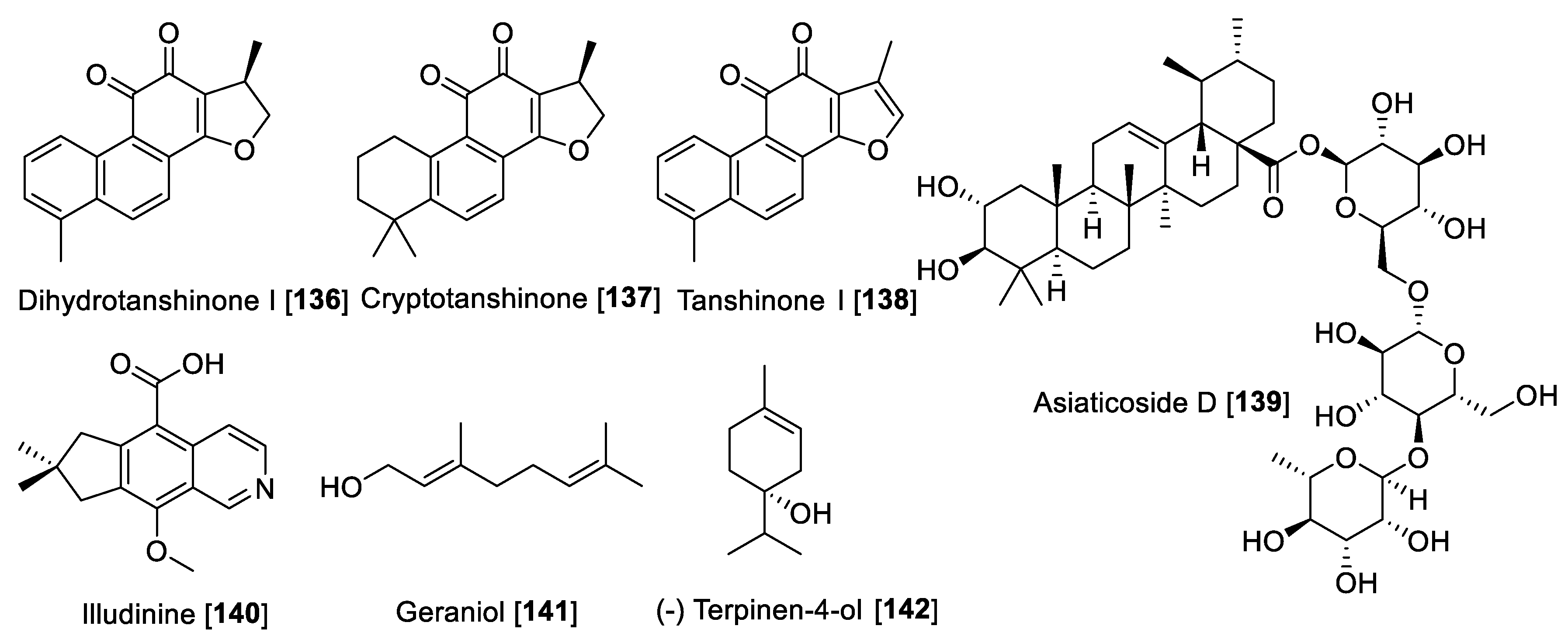
5.6. MAO Inhibitory Terpenes and Terpenoids
5.7. MAO Inhibitors from Marine Sources
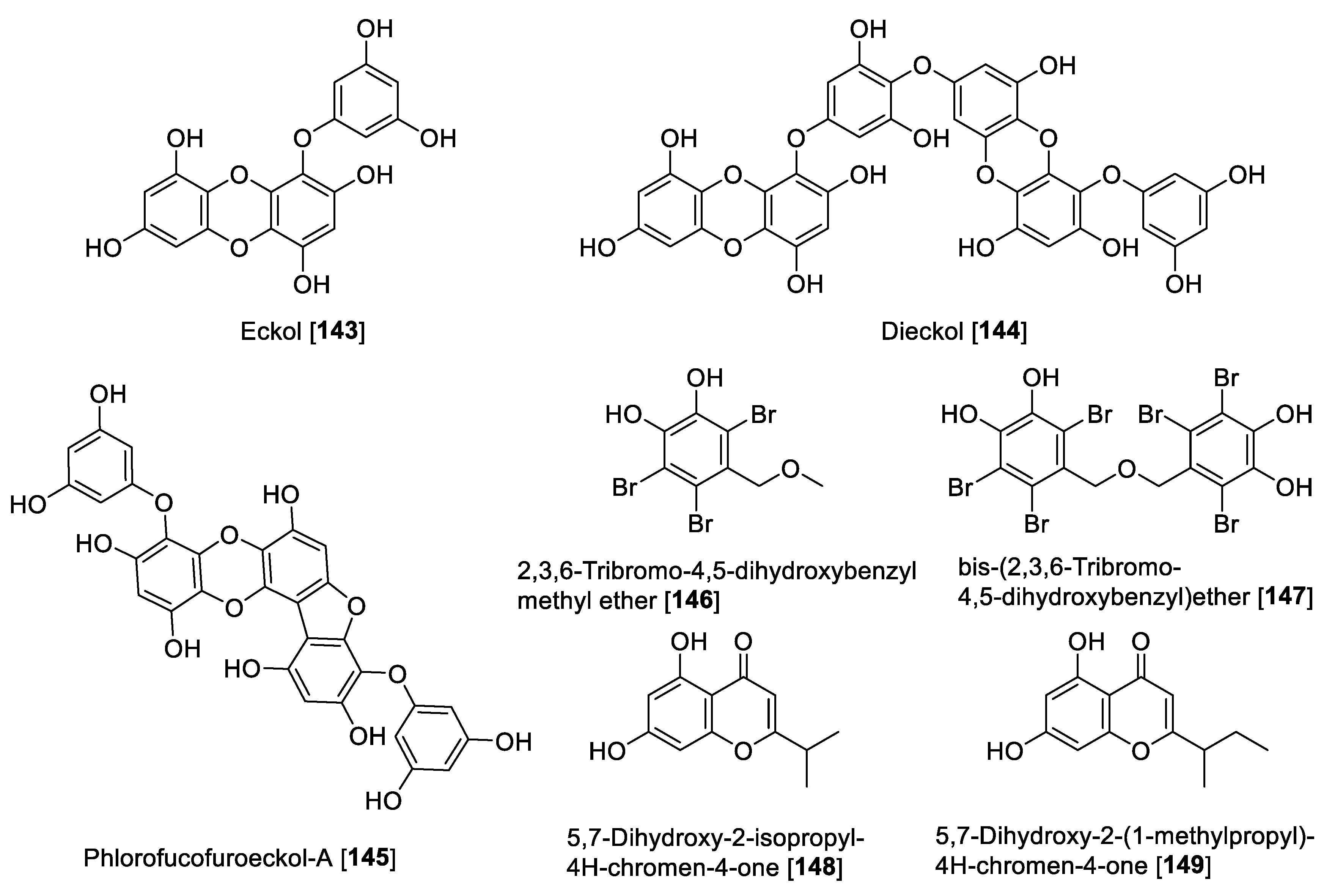
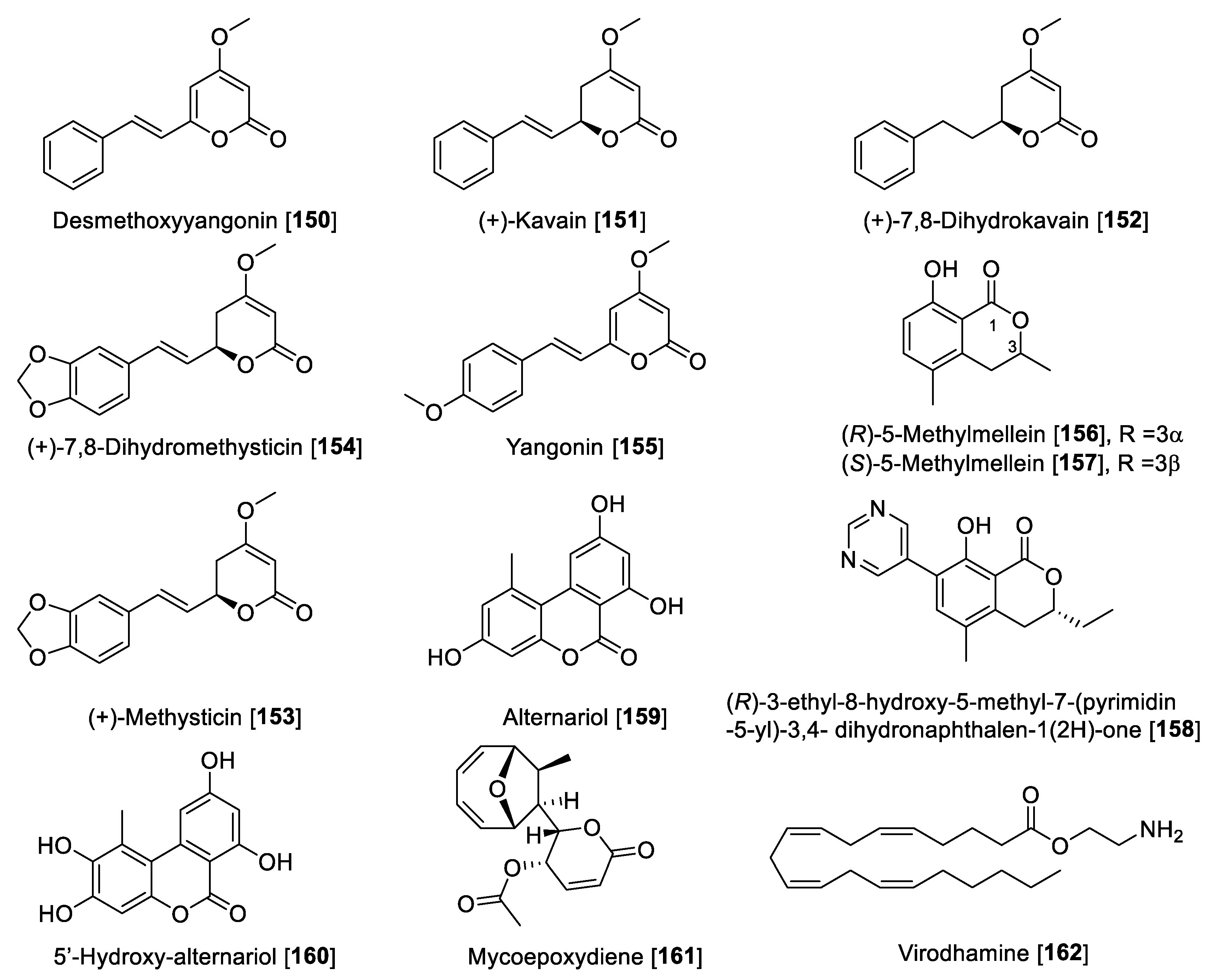
5.8. MAO Inhibitors from Miscellaneous Classes of Natural Products
| Compounds | Source | MAO-A | MAO-B | SI | Enzyme Source | References | ||
|---|---|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | IC50 (μM) | Ki (μM) | MAOA/B | ||||
| Dihydrotanshinone I [136] | Salvia miltiorrhiza | 23 | F | [183] | ||||
| Cryptotanshinone [137] | Salvia miltiorrhiza | 80 | F | [183] | ||||
| Tanshinone I [138] | Salvia miltiorrhiza | 84 | F | [183] | ||||
| Illudinine [140] | Clitocybe illudens | 18.3 | F | [185] | ||||
| Eckol [143] | Eisenia bicyclis | 7.20 | 20.26 | 83.44 | 162.8 | 0.086 | F | [185] |
| Dieckol [144] | Eisenia bicyclis | 11.43 | 20.28 | 43.42 | 18.50 | 0.26 | F | [190] |
| Phlorofucofuroeckol-A [145] | Ecklonia stolonifera | 9.22 | 5.18 | 4.89 | 2.69 | 1.88 | F | [190] |
| 2,3,6-Tribromo-4,5-dihydroxybenzyl methyl ether [146] | Symphyocladia latiuscula | 63.16 | 25.4 | 105.13 | 40.7 | 0.60 | F | [191] |
| bis-(2,3,6-Tribromo-4,5-dihydroxybenzyl) ether [147] | Symphyocladia latiuscula | 89.31 | 22.8 | 102.53 | 35.5 | 0.87 | F | [191] |
| 5,7-Dihydroxy-2-isopropyl-4H-chromen-4-one [148] | Streptomyces spp. | 2.70 | 27.0 | 0.10 | F | [192] | ||
| 5,7-Dihydroxy-2-(1-methylpropyl)-4H-chromen-4-one [149] | Streptomyces spp. | 6.92 | 3.42 | 2.02 | F | [192] | ||
| Desmethoxyyangonin [150] | Renealmia alpinia | 1.85 | 0.922 | 0.12 | 0.031 | 15.41 | F | [22] |
| (+)-Kavain [151] | Piper methysticum | 19.0 | 5.34 | 3.55 | F | [193] | ||
| (+)-7,8-Dihydrokavain [152] | Piper methysticum | >100 | 8.23 | 12.15 | F | [193] | ||
| (+)-Methysticin [153] | Piper methysticum | 8.12 | 0.42 | 19.33 | F | [193] | ||
| (+)-7,8-Dihydromethysticin [154] | Piper methysticum | 23.2 | 0.855 | 27.13 | F | [193] | ||
| Yangonin [155] | Piper methysticum | 1.29 | 1.12 | 0.085 | 0.226 | 15.17 | F | [193] |
| (S)-5-Methymellein [156] | Rosellinia corticium | 5.31 | 2.45 | 9.15 | 0.58 | F | [194] | |
| (R)-5-Methylmellein [157] | Xylaria nigripes | 4.6 | 38.5 | 0.11 | F | [195] | ||
| (R)-3-Ethyl-8-hydroxy-5-methyl-7-(pyrimidin-5-yl)-3,4- dihydronaphthalen-1(2H)-one [158] | Methylmellein derivative | 0.06 | >50 | >0.012 | F | [195] | ||
| Alternariol [159] | Diaporthe mahothocarpus | 0.020 | 0.0075 | 20.7 | 0.00096 | F | [196] | |
| 5′-Hydroxy-alternariol [160] | Diaporthe mahothocarpus | 0.31 | 0.116 | >40 | 0.00775 | F | [196] | |
| Mycoepoxydiene [161] | Diaporthe mahothocarpus | 8.7 | 3.76 | >40 | >0.21 | F | [196] | |
| Virodhamine [162] | 38.7 | 0.71 | 0.258 | 54.50 | F | [196] | ||
6. Natural MAO Inhibitors in Neuroblastoma
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edmondson, D.E.; Binda, C. Monoamine Oxidases. Subcell Biochem. 2018, 87, 117–139. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R. Monoamine oxidases: The biochemistry of the proteins as targets in medicinal chemistry and drug discovery. Curr. Top. Med. Chem. 2012, 12, 2189–2209. [Google Scholar] [CrossRef] [PubMed]
- Klinman, J.P. The multi-functional topa-quinone copper amine oxidases. Biochim. Biophys. Acta 2003, 1647, 131–137. [Google Scholar] [CrossRef]
- Abell, C.W.; Kwan, S.W. Molecular characterization of monoamine oxidases A and B. Prog. Nucleic Acid. Res. Mol. Biol. 2001, 65, 129–156. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Castillo, G.; Sullivan, P.; Sharabi, Y. Differential Susceptibilities of Catecholamines to Metabolism by Monoamine Oxidases. J. Pharmacol. Exp. Ther. 2021, 379, 253–259. [Google Scholar] [CrossRef]
- Ramonet, D.; Rodriguez, M.; Saura, J.; Lizcano, J.M.; Romera, M.; Unzeta, M.; Finch, C.; Billett, E.; Mahy, N. Localization of monoamine oxidase A and B and semicarbazide-sensitive amine oxidase in human peripheral tissues. Inflammopharmacology 2003, 11, 111–117. [Google Scholar] [CrossRef]
- Rodriguez, M.J.; Saura, J.; Billett, E.E.; Finch, C.C.; Mahy, N. Cellular localization of monoamine oxidase A and B in human tissues outside of the central nervous system. Cell Tissue Res. 2001, 304, 215–220. [Google Scholar] [CrossRef]
- Hubalek, F.; Pohl, J.; Edmondson, D.E. Structural comparison of human monoamine oxidases A and B: Mass spectrometry monitoring of cysteine reactivities. J. Biol. Chem. 2003, 278, 28612–28618. [Google Scholar] [CrossRef] [Green Version]
- Bach, A.W.; Lan, N.C.; Johnson, D.L.; Abell, C.W.; Bembenek, M.E.; Kwan, S.W.; Seeburg, P.H.; Shih, J.C. cDNA cloning of human liver monoamine oxidase A and B: Molecular basis of differences in enzymatic properties. Proc. Natl. Acad. Sci. USA 1988, 85, 4934–4938. [Google Scholar] [CrossRef] [Green Version]
- Hubalek, F.; Binda, C.; Khalil, A.; Li, M.; Mattevi, A.; Castagnoli, N.; Edmondson, D.E. Demonstration of isoleucine 199 as a structural determinant for the selective inhibition of human monoamine oxidase B by specific reversible inhibitors. J. Biol. Chem. 2005, 280, 15761–15766. [Google Scholar] [CrossRef] [Green Version]
- Son, S.Y.; Ma, J.; Kondou, Y.; Yoshimura, M.; Yamashita, E.; Tsukihara, T. Structure of human monoamine oxidase A at 2.2-A resolution: The control of opening the entry for substrates/inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 5739–5744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Colibus, L.; Li, M.; Binda, C.; Lustig, A.; Edmondson, D.E.; Mattevi, A. Three-dimensional structure of human monoamine oxidase A (MAO A): Relation to the structures of rat MAO A and human MAO B. Proc. Natl. Acad. Sci. USA 2005, 102, 12684–12689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Yoshimura, M.; Yamashita, E.; Nakagawa, A.; Ito, A.; Tsukihara, T. Structure of rat monoamine oxidase A and its specific recognitions for substrates and inhibitors. J. Mol. Biol. 2004, 338, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Kubota, F.; Yoshimura, M.; Yamashita, E.; Nakagawa, A.; Ito, A.; Tsukihara, T. Crystallization and preliminary crystallographic analysis of rat monoamine oxidase A complexed with clorgyline. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Binda, C.; Li, M.; Hubalek, F.; Restelli, N.; Edmondson, D.E.; Mattevi, A. Insights into the mode of inhibition of human mitochondrial monoamine oxidase B from high-resolution crystal structures. Proc. Natl. Acad. Sci. USA 2003, 100, 9750–9755. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.N.; Raghanti, M.A. The role of monoamine oxidase enzymes in the pathophysiology of neurological disorders. J. Chem. Neuroanat 2021, 114, 101957. [Google Scholar] [CrossRef]
- Fowler, J.S.; Volkow, N.D.; Logan, J.; Franceschi, D.; Wang, G.J.; MacGregor, R.; Shea, C.; Garza, V.; Pappas, N.; Carter, P.; et al. Evidence that L-deprenyl treatment for one week does not inhibit MAO A or the dopamine transporter in the human brain. Life Sci. 2001, 68, 2759–2768. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Leon, F.; Ding, Y.; Gomez-Betancur, I.; Benjumea, D.; Walker, L.A.; Cutler, S.J.; Tekwani, B.L. Interactions of Desmethoxyyangonin, a Secondary Metabolite from Renealmia alpinia, with Human Monoamine Oxidase-A and Oxidase-B. Evid. Based Compl. Alternat Med. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liebowitz, M.R.; Hollander, E.; Schneier, F.; Campeas, R.; Welkowitz, L.; Hatterer, J.; Fallon, B. Reversible and irreversible monoamine oxidase inhibitors in other psychiatric disorders. Acta Psychiatr. Scand. Suppl. 1990, 360, 29–34. [Google Scholar] [CrossRef]
- Priest, R.G.; Gimbrett, R.; Roberts, M.; Steinert, J. Reversible and selective inhibitors of monoamine oxidase A in mental and other disorders. Acta Psychiatr. Scand. Suppl. 1995, 386, 40–43. [Google Scholar] [CrossRef]
- Amrein, R.; Hetzel, W.; Stabl, M.; Schmid-Burgk, W. RIMA: A safe concept in the treatment of depression with moclobemide. Can. J. Psychiatry 1992, 37 (Suppl. S1), 7–11. [Google Scholar] [PubMed]
- Mann, J.J.; Aarons, S.F.; Frances, A.J.; Brown, R.D. Studies of selective and reversible monoamine oxidase inhibitors. J. Clin. Psychiatry 1984, 45, 62–66. [Google Scholar] [PubMed]
- Suchting, R.; Tirumalajaru, V.; Gareeb, R.; Bockmann, T.; de Dios, C.; Aickareth, J.; Pinjari, O.; Soares, J.C.; Cowen, P.J.; Selvaraj, S. Revisiting monoamine oxidase inhibitors for the treatment of depressive disorders: A systematic review and network meta-analysis. J. Affect Disord. 2021, 282, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.B.; Ding, C.Z.; Gates, K.S. Design and meachanism of monoamine oxidase inactivators from an organic chemical perspective. In Perspectives in Medicinal Chemistry; Testa, B., Kyburz, E., Fuhrer, W., Giger, R., Eds.; Verlag Helvetica Chimica: Basel, Switzerland, 1993; pp. 73–86. [Google Scholar]
- Kanazawa, I. Short review on monoamine oxidase and its inhibitors. Eur. Neurol. 1994, 34, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Binda, C.; Newton-Vinson, P.; Hubalek, F.; Edmondson, D.E.; Mattevi, A. Structure of human monoamine oxidase B, a drug target for the treatment of neurological disorders. Nat. Struct. Biol. 2002, 9, 22–26. [Google Scholar] [CrossRef]
- Dhiman, P.; Malik, N.; Sobarzo-Sanchez, E.; Uriarte, E.; Khatkar, A. Quercetin and Related Chromenone Derivatives as Monoamine Oxidase Inhibitors: Targeting Neurological and Mental Disorders. Molecules 2019, 24, 418. [Google Scholar] [CrossRef] [Green Version]
- Bolasco, A.; Carradori, S.; Fioravanti, R. Focusing on new monoamine oxidase inhibitors. Exp. Opin. Ther. Pat. 2010, 20, 909–939. [Google Scholar] [CrossRef]
- Riederer, P.L.G. MAO-inhibitors in Parkinson’s Disease. Exp. Neurobiol. 2011, 20, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kirwin, J.L.; Gören, J.L. Duloxetine: A dual serotonin-norepinephrinereuptake inhibitor for treatment of major depressive disorder. Pharmacotherapy 2005, 25, 396–410. [Google Scholar] [CrossRef]
- Moret, C.; Isaac, M.; Briley, M. Problems associated with long-termtreatment with selective serotonin reuptake inhibitors. J. Psychopharmacol. 2009, 23, 967–974. [Google Scholar] [CrossRef]
- Fowler, J.S.; Logan, J.; Azzaro, A.J.; Fielding, R.M.; Zhu, W.; Poshusta, A.K.; Burch, D.; Brand, B.; Free, J.; Asgharnejad, M.; et al. Reversible inhibitors of monoamine oxidase-A (RIMAs): Robust, reversible inhibition of human brain MAO-A by CX157. Neuropsychopharmacology 2010, 35, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Gleiter, C.H.; Muhlbauer, B.; Schulz, R.M.; Nilsson, E.; Antonin, K.H.; Bieck, P.R. Monoamine oxidase inhibition by the MAO-A inhibitors brofaromine and clorgyline in healthy volunteers. J. Neural Transm. Gen. Sect. 1994, 95, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, M.; Brown-Borg, H.; Ren, J.; Sharma, S.; Shavali, S.; El ReFaey, H.; Carlson, E.C. Therapeutic efficacy of selegiline in neurodegenerative disorders and neurological diseases. Curr. Drug Targets 2006, 7, 1513–1529. [Google Scholar] [CrossRef] [PubMed]
- Riederer, P.; Lachenmayer, L.; Laux, G. Clinical applications of MAO-inhibitors. Curr. Med. Chem. 2004, 11, 2033–2043. [Google Scholar] [CrossRef]
- Youdim, M.B.; Bakhle, Y.S. Monoamine oxidase: Isoforms and inhibitors in Parkinson’s disease and depressive illness. Br. J. Pharmacol. 2006, 147 (Suppl. S1), S287–S296. [Google Scholar] [CrossRef] [Green Version]
- Stocchi, F. Rasagiline: Defining the role of a novel therapy in the treatment of Parkinson’s disease. Int. J. Clin. Pract. 2006, 60, 215–221. [Google Scholar] [CrossRef]
- Lecht, S.; Haroutiunian, S.; Hoffman, A.; Lazarovici, P. Rasagiline-a novel MAO B inhibitor in Parkinson’s disease therapy. Ther. Clin. Risk Manag. 2007, 3, 467–474. [Google Scholar]
- Chen, J.J.; Ly, A.V. Rasagiline: A second-generation monoamine oxidase type-B inhibitor for the treatment of Parkinson’s disease. Am. J. Health Syst. Pharm. 2006, 63, 915–928. [Google Scholar] [CrossRef]
- McGrath, P.J.; Blood, D.K.; Stewart, J.W.; Harrison, W.; Quitkin, F.M.; Tricamo, E.; Markowitz, J. A comparative study of the electrocardiographic effects of phenelzine, tricyclic antidepressants, mianserin, and placebo. J. Clin. Psychopharmacol. 1987, 7, 335–339. [Google Scholar] [CrossRef]
- Tibrewal, P.; Kanigere, M.K.; Looi, J.C.; Allison, S.; Bastiampillai, T. The evidence for the use of long-term benzodiazepines in the setting of treatment-refractory anxiety disorders. Aust. N. Z. J. Psychiatry 2022, 56, 723–724. [Google Scholar] [CrossRef]
- Sathyanarayana Rao, T.S.; Yeragani, V.K. Hypertensive crisis and cheese. Indian J Psychiatry 2009, 51, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Flockhart, D.A. Dietary restrictions and drug interactions with monoamine oxidase inhibitors: An update. J. Clin. Psychiatry 2012, 73 (Suppl. S1), 17–24. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, D.S. Sexual dysfunction associated with antidepressant drugs. Exp. Opin. Drug Saf. 2004, 3, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Kuhn, K.U.; Strater, B.; Scherbaum, N.; Weig, W. Adverse side-effect on sexual function caused by psychotropic drugs and psychotropic substances. Nervenarzt 2010, 81, 1129–1137. [Google Scholar] [CrossRef]
- Mago, R.T.N.; Andrade, C. Cardiovascular adverse effects of newer antidepressants. Exp. Rev. Neurother. 2014, 14, 539–551. [Google Scholar] [CrossRef]
- Reichenpfader, U.G.G.; Morgan, L.C.; Greenblatt, A.; Nussbaumer, B.; Hansen, R.A. Sexual dysfunction associated with second-generation antidepressants in patients with major depressive disorder: Results from a systematic review with network meta-analysis. Drug Saf. 2014, 37, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Cappetta, K.B.C.; Johnson, J.A.; Bloch, M.H. Meta-analysis: Risk of dry mouth with second generation antidepressants. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 84, 282–293. [Google Scholar] [CrossRef]
- Rapaport, M.H. Dietary restrictions and drug interactions with monoamine oxidase inhibitors: The state of the art. J. Clin. Psychiatry 2007, 68 (Suppl. S8), 42–46. [Google Scholar]
- Remick, R.A. Anticholinergic side effects of tricyclic antidepressants and their management. Prog. Neuropsychopharmacol. Biol. Psychiatry 1988, 12, 225–231. [Google Scholar] [CrossRef]
- Brambilla, P.C.A.; Hotopf, M.; Barbui, C. Side-effect profile of fluoxetine in comparison with other SSRIs, tricyclic and newer antidepressants: A meta-analysis of clinical trial data. Pharmacopsychiatry 2005, 38, 69–77. [Google Scholar] [CrossRef]
- Nair, N.P.; Ahmed, S.K.; Kin, N.M.; West, T.E. Reversible and selective inhibitors of monoamine oxidase A in the treatment of depressed elderly patients. Acta Psychiatry Scand. Suppl. 1995, 386, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Sarris, J. Herbal medicines in the treatment of psychiatric disorders: 10-year updated review. Phytother. Res. 2018, 32, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Moragrega, I.; Rios, J.L. Medicinal Plants in the Treatment of Depression: Evidence from Preclinical Studies. Planta Med. 2021, 87, 656–685. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.S.B. Phytochemistry and pharmacology of anti-depressant medicinal plants: A review. Biomed. Pharmacother. 2018, 104, 343–365. [Google Scholar] [CrossRef]
- Geck, M.S.; Lecca, D.; Marchese, G.; Casu, L.; Leonti, M. Ethnomedicine and neuropsychopharmacology in Mesoamerica. J. Ethnopharmacol. 2021, 278, 114243. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef]
- Ji, H.F.; Zhang, H.Y. Multipotent natural agents to combat Alzheimer’s disease. Functional spectrum and structural features. Acta Pharmacol. Sin. 2008, 29, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Schultes, R.E.; Raffauf, R.F. The Healing Forest: Medicinal and Toxic Plants of the Northwest Amazonia; Dioscorides Press: Portland, OR, USA, 1990. [Google Scholar]
- Samoylenko, V.; Rahman, M.M.; Tekwani, B.L.; Tripathi, L.M.; Wang, Y.H.; Khan, S.I.; Khan, I.A.; Miller, L.S.; Joshi, V.C.; Muhammad, I. Banisteriopsis caapi, a unique combination of MAO inhibitory and antioxidative constituents for the activities relevant to neurodegenerative disorders and Parkinson’s disease. J. Ethnopharmacol. 2010, 127, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Glover, V.; Liebowitz, J.; Armando, I.; Sandler, M. beta-Carbolines as selective monoamine oxidase inhibitors: In vivo implications. J. Neural Transm. 1982, 54, 209–218. [Google Scholar] [CrossRef]
- Morales-Garcia, J.A.; de la Fuente Revenga, M.; Alonso-Gil, S.; Rodriguez-Franco, M.I.; Feilding, A.; Perez-Castillo, A.; Riba, J. The alkaloids of Banisteriopsis caapi, the plant source of the Amazonian hallucinogen Ayahuasca, stimulate adult neurogenesis in vitro. Sci. Rep. 2017, 7, 5309. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Saha, S.C.; Sunita, K.; Majumder, M.; Ghorai, M.; Mane, A.B.; Prasanth, D.A.; Kumar, P.; Pandey, D.K.; Al-Tawaha, A.R.; et al. Promising botanical-derived monoamine oxidase (MAO) inhibitors: Pharmacological aspects and structure-activity studies. S. Afr. J. Bot. 2022, 146, 127–145. [Google Scholar] [CrossRef]
- Ege, T.S.H.D. Monoamine oxidase inhibitory effects of medicinal plants in management of Alzheimer’s dis-ease. J. Turk. Chem. Soc. 2021, 8, 239–248. [Google Scholar]
- Verpoorte, R. Alkaloids in Encyclopedia of Analytical Sciences, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier Sciences: London, UK, 2005. [Google Scholar]
- Fattorusso, E.; Taglialatela-Scafati, O. Modern Alkaloids: Structure, Isolation, Synthesis, and Biology; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2008; p. 689. [Google Scholar]
- Schurr, A.; Livne, A. Differential inhibition of mitochondrial monoamine oxidase from brain by hashish components. Biochem. Pharmacol. 1976, 25, 1201–1203. [Google Scholar] [CrossRef]
- Kong, L.D.; Cheng, C.H.; Tan, R.X. Inhibition of MAO A and B by some plant-derived alkaloids, phenols and anthraquinones. J. Ethnopharmacol. 2004, 91, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Hong, S.S.; Han, X.H.; Hwang, J.S.; Oh, G.J.; Lee, K.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Piperine from the fruits of Piper longum with inhibitory effect on monoamine oxidase and antidepressant-like activity. Chem. Pharm. Bull. 2005, 53, 832–835. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Hwang, J.S.; Han, X.H.; Lee, C.; Lee, M.H.; Choe, S.G.; Hong, S.S.; Lee, D.; Lee, M.K.; Hwang, B.Y. Methylpiperate derivatives from Piper longum and their inhibition of monoamine oxidase. Arch. Pharm. Res. 2008, 31, 679–683. [Google Scholar] [CrossRef]
- Mu, L.H.; Wang, B.; Ren, H.Y.; Liu, P.; Guo, D.H.; Wang, F.M.; Bai, L.; Guo, Y.S. Synthesis and inhibitory effect of piperine derivates on monoamine oxidase. Bioorg. Med. Chem. Lett. 2012, 22, 3343–3348. [Google Scholar] [CrossRef]
- Ro, J.S.; Lee, S.S.; Lee, K.S.; Lee, M.K. Inhibition of type A monoamine oxidase by coptisine in mouse brain. Life Sci. 2001, 70, 639–645. [Google Scholar] [CrossRef]
- Lee, S.S.; Lee, M.K. Effects of natural isoquinoline alkaloids on monoamine oxidase activity in mouse brain: Inhibition by berberine and palmatine. Med. Sci. Res. 1999, 27, 749–751. [Google Scholar]
- Lee, S.J.; Chung, H.Y.; Lee, I.K.; Oh, S.U.; Yoo, I.D. Phenolics with Inhibitory Activity on Mouse Brain Monoamine Oxidase (MAO) from Whole Parts of Artemisia vulgaris L (Mugwort). Food Sci. Biotechnol. 2000, 9, 179–182. [Google Scholar]
- Lee, S.S.; Yun-Choi, H.S.; Kim, E.I.; Lee, M.K. Inhibition of monoamine oxidase by higenamine. Med. Sci. Res. 1999, 27, 71–72. [Google Scholar]
- Plazas, E.; Hagenow, S.; Avila Murillo, M.; Stark, H.; Cuca, L.E. Isoquinoline alkaloids from the roots of Zanthoxylum rigidum as multi-target inhibitors of cholinesterase, monoamine oxidase A and Abeta1-42 aggregation. Bioorg. Chem. 2020, 98, 103722. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.J.; Yang, Z.D.; Li, Z.J.; Yang, S.H.; Shu, Z.M. Stephtetrandrine A-D, bisbenzylisoquinoline alkaloids from Stephania tetrandra. Nat. Prod. Res. 2021, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Hwang, B.Y.; Lee, S.A.; Oh, G.J.; Choi, W.H.; Hong, S.S.; Lee, K.S.; Ro, J.S. 1-methyl-2-undecyl-4(1H)-quinolone as an irreversible and selective inhibitor of type B monoamine oxidase. Chem. Pharm. Bull. 2003, 51, 409–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.F.; Chiou, W.F.; Chou, C.J.; Liao, J.F.; Lin, L.C.; Wang, G.J.; Ueng, Y.F. Pharmacological effects of Evodia rutaecarpa and its bioactive components. Chin. Pharm. J. 2002, 54, 419–435. [Google Scholar]
- Han, X.H.; Hong, S.S.; Lee, D.; Lee, J.J.; Lee, M.S.; Moon, D.C.; Han, K.; Oh, K.W.; Lee, M.K.; Ro, J.S.; et al. Quinolone alkaloids from evodiae fructus and their inhibitory effects on monoamine oxidase. Arch. Pharm. Res. 2007, 30, 397–401. [Google Scholar] [CrossRef]
- Mesiti, F.; Maruca, A.; Silva, V.; Rocca, R.; Fernandes, C.; Remiao, F.; Uriarte, E.; Alcaro, S.; Gaspar, A.; Borges, F. 4-Oxoquinolines and monoamine oxidase: When tautomerism matters. Eur. J. Med. Chem. 2021, 213, 113183. [Google Scholar] [CrossRef]
- Naidoo, D.; Roy, A.; Slavětínská, L.P.; Chukwujekwu, J.C.; Gupta, S.; Van Staden, J. New role for crinamine as a potent, safe and selective inhibitor of human monoamine oxidase B: In vitro and in silico pharmacology and modeling. J. Ethnopharmacol. 2020, 248, 112305. [Google Scholar] [CrossRef]
- Fernandez de Arriba, A.; Lizcano, J.M.; Balsa, M.D.; Unzeta, M. Inhibition of monoamine oxidase from bovine retina by beta-carbolines. J. Pharm. Pharmacol. 1994, 46, 809–813. [Google Scholar] [CrossRef]
- Temple, W.; Mendelsohn, L.; Kim, G.E.; Nekritz, E.; Gustafson, W.C.; Lin, L.; Giacomini, K.; Naranjo, A.; Van Ryn, C.; Yanik, G.A.; et al. Vesicular monoamine transporter protein expression correlates with clinical features, tumor biology, and MIBG avidity in neuroblastoma: A report from the Children’s Oncology Group. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Herraiz, T.; Chaparro, C. Analysis of monoamine oxidase enzymatic activity by reversed-phase high performance liquid chromatography and inhibition by beta-carboline alkaloids occurring in foods and plants. J. Chromatogr. 2006, 1120, 237–243. [Google Scholar] [CrossRef]
- Herraiz, T.; González, D.; Ancín-Azpilicueta, C.; Arán, V.J.; Guillén, H. Beta-Carboline alkaloids in Peganum harmala and inhibition of human monoamine oxidase (MAO). Arch. Pharm. Res. 2010, 48, 839–845. [Google Scholar]
- McKenna, D.J.; Towers, G.N.; Abbott, F. Monoamine oxidase inhibitors in south American Hallucinogenic Plants Part 2: Constituents of orally active myristicaceous hallucinogens. J. Ethanopharmacol. 1984, 12, 179–211. [Google Scholar] [CrossRef]
- Denis, J.M.; Towers, G.H.N.; Abbott, F.S. Monoamine oxidase inhibitors in south American Hallucinogenic Plants: Tryptamine and beta-carboline constituents of Ayahuasca. J. Ethanopharmacol. 1984, 10, 195–223. [Google Scholar]
- Schwarz, M.J.; Houghton, P.J.; Rose, S.; Jenner, P.; Lees, A.D. Activities of extract and constituents of Banisteriopsis caapi relevant to parkinsonism. Pharmacol. Biochem. Behav. 2003, 75, 627–633. [Google Scholar] [CrossRef]
- Hong, S.S.; Han, X.H.; Park, S.Y.; Choi, W.H.; Lee, M.K.; Hur, J.D.; Ro, J.S. Monoamine Oxidase inhibitors from Uncaria rhynchophylla. Nat. Prod. Sci. 2005, 11, 145–149. [Google Scholar]
- Reniers, J.; Robert, S.; Frederick, R.; Masereel, B.; Vincent, S.; Wouters, J. Synthesis and evaluation of beta-carboline derivatives as potential monoamine oxidase inhibitors. Bioorg. Med. Chem. 2011, 19, 134–144. [Google Scholar] [CrossRef]
- Calixto, N.O.; Pinto, M.E.F.; Ramalho, S.D.; Burger, M.; Bobey, A.F.; Young, M.C.M.; Pinto, A.C. The genus Psychotria: Phytochemistry, chemotaxonomy, ethnopharmacology and biological properties. J. Braz. Chem. Soc. 2016, 27, 1355–1378. [Google Scholar]
- Passos, C.S.; Simoes-Pires, C.A.; Nurisso, A.; Soldi, T.C.; Kato, L.; de Oliveira, C.M.; de Faria, E.O.; Marcourt, L.; Gottfried, C.; Carrupt, P.A.; et al. Indole alkaloids of Psychotria as multifunctional cholinesterases and monoamine oxidases inhibitors. Phytochemistry 2013, 86, 8–20. [Google Scholar] [CrossRef]
- Klein-Junior, L.C.; Cretton, S.; Vander Heyden, Y.; Gasper, A.L.; Nejad-Ebrahimi, S.; Christen, P.; Henriques, A.T. Bioactive Azepine-Indole Alkaloids from Psychotria nemorosa. J. Nat. Prod. 2020, 83, 852–863. [Google Scholar] [CrossRef]
- Milan, S. Neurodynamics of Prosocial Emotional Processing Following Serotonergic Stimulation with N,N-Dimethyltryptamine (DMT) and Harmine in Healthy Subjects. Available online: https://clinicaltrials.gov/ct2/show/NCT04716335 (accessed on 29 April 2022).
- Zhi, K.K.; Yang, Z.D.; Shi, D.F.; Yao, X.J.; Wang, M.G. Desmodeleganine, a new alkaloid from the leaves of Desmodium elegans as a potential monoamine oxidase inhibitor. Fitoterapia 2014, 98, 160–165. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are Flavonoids Effective Antioxidants in Plants? Twenty Years of Our Investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef]
- Gogineni, V.; Nael, M.A.; Chaurasiya, N.D.; Elokely, K.M.; McCurdy, C.R.; Rimoldi, J.M.; Cutler, S.J.; Tekwani, B.L.; Leon, F. Computationally Assisted Lead Optimization of Novel Potent and Selective MAO-B Inhibitors. Biomedicines 2021, 9, 1304. [Google Scholar] [CrossRef]
- Carradori, S.; Gidaro, M.C.; Petzer, A.; Costa, G.; Guglielmi, P.; Chimenti, P.; Alcaro, S.; Petzer, J.P. Inhibition of Human Monoamine Oxidase: Biological and Molecular Modeling Studies on Selected Natural Flavonoids. J. Agric. Food Chem. 2016, 64, 9004–9011. [Google Scholar] [CrossRef]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. Biomed. Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kregiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [Green Version]
- Chaurasiya, N.D.; Ibrahim, M.A.; Muhammad, I.; Walker, L.A.; Tekwani, B.L. Monoamine oxidase inhibitory constituents of propolis: Kinetics and mechanism of inhibition of recombinant human MAO-A and MAO-B. Molecules 2014, 19, 18936–18952. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Lazaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Park, S.E.; Paudel, P.; Wagle, A.; Seong, S.H.; Kim, H.R.; Fauzi, F.M.; Jung, H.A.; Choi, J.S. Luteolin, a Potent Human Monoamine Oxidase-A Inhibitor and Dopamine D4 and Vasopressin V1A Receptor Antagonist. J. Agric. Food Chem. 2020, 68, 10719–10729. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Gogineni, V.; Elokely, K.M.; Leon, F.; Nunez, M.J.; Klein, M.L.; Walker, L.A.; Cutler, S.J.; Tekwani, B.L. Isolation of Acacetin from Calea urticifolia with Inhibitory Properties against Human Monoamine Oxidase-A and -B. J. Nat. Prod. 2016, 79, 2538–2544. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Zhao, J.; Pandey, P.; Doerksen, R.J.; Muhammad, I.; Tekwani, B.L. Selective Inhibition of Human Monoamine Oxidase B by Acacetin 7-Methyl Ether Isolated from Turnera diffusa (Damiana). Molecules 2019, 24, 810. [Google Scholar] [CrossRef] [Green Version]
- Larit, F.; Elokely, K.M.; Chaurasiya, N.D.; Benyahia, S.; Nael, M.A.; Leon, F.; Abu-Darwish, M.S.; Efferth, T.; Wang, Y.H.; Belouahem-Abed, D.; et al. Inhibition of human monoamine oxidase A and B by flavonoids isolated from two Algerian medicinal plants. Phytomedicine 2018, 40, 27–36. [Google Scholar] [CrossRef]
- Baek, S.C.; Kang, M.G.; Park, J.E.; Lee, J.P.; Lee, H.; Ryu, H.W.; Kim, H. Osthenol, a prenylated coumarin, as a monoamine oxidase A inhibitor with high selectivity. Bioorg. Med. Chem. Lett. 2019, 29, 839–843. [Google Scholar] [CrossRef]
- Turkmenoglu, F.P.; Baysal, I.; Ciftci-Yabanoglu, S.; Yelekci, K.; Temel, H.; Pasa, S.; Ezer, N.; Calis, I.; Ucar, G. Flavonoids from Sideritis Species: Human Monoamine Oxidase (hMAO) Inhibitory Activities, Molecular Docking Studies and Crystal Structure of Xanthomicrol. Molecules 2015, 20, 7454–7473. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Doseff, A.I.; Grotewold, E. Flavones: From Biosynthesis to Health Benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef]
- Bello, O.M.; Ogbesejana, A.B.; Adetunji, C.O.; Oguntoye, S.O. Flavonoids Isolated from Vitex grandifolia, an Underutilized Vegetable, Exert Monoamine A & B Inhibitory and Anti-inflammatory Effects and Their Structure-activity Relationship. Turk J. Pharm. Sci. 2019, 16, 437–443. [Google Scholar] [CrossRef]
- Lee, H.W.; Ryu, H.W.; Baek, S.C.; Kang, M.G.; Park, D.; Han, H.Y.; An, J.H.; Oh, S.R.; Kim, H. Potent inhibitions of monoamine oxidase A and B by acacetin and its 7-O-(6-O-malonylglucoside) derivative from Agastache rugosa. Int. J. Biol. Macromol. 2017, 104, 547–553. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: Chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef]
- Chimenti, F.; Cottiglia, F.; Bonsignore, L.; Casu, L.; Casu, M.; Floris, C.; Secci, D.; Bolasco, A.; Chimenti, P.; Granese, A.; et al. Quercetin as the active principle of Hypericum hircinum exerts a selective inhibitory activity against MAO-A: Extraction, biological analysis, and computational study. J. Nat. Prod. 2006, 69, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Gidaro, M.C.; Astorino, C.; Petzer, A.; Carradori, S.; Alcaro, F.; Costa, G.; Artese, A.; Rafele, G.; Russo, F.M.; Petzer, J.P.; et al. Kaempferol as Selective Human MAO-A Inhibitor: Analytical Detection in Calabrian Red Wines, Biological and Molecular Modeling Studies. J. Agric. Food Chem. 2016, 64, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Novaroli, L.; Reist, M.; Favre, E.; Carotti, A.; Catto, M.; Carrupt, P.A. Human recombinant monoamine oxidase B as reliable and efficient enzyme source for inhibitor screening. Bioorg. Med. Chem. 2005, 13, 6212–6217. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.C.; Park, M.H.; Ryu, H.W.; Lee, J.P.; Kang, M.G.; Park, D.; Park, C.M.; Oh, S.R.; Kim, H. Rhamnocitrin isolated from Prunus padus var. seoulensis: A potent and selective reversible inhibitor of human monoamine oxidase A. Bioorg. Chem. 2019, 83, 317–325. [Google Scholar] [CrossRef]
- Hwang, J.S.; Lee, S.A.; Hong, S.S.; Lee, K.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Monoamine oxidase inhibitory components from the roots of Sophora flavescens. Arch. Pharm. Res. 2005, 28, 190–194. [Google Scholar] [CrossRef]
- Zarmouh, N.O.; Eyunni, S.K.; Soliman, K.F. The Benzopyrone Biochanin-A as a reversible, competitive, and selective monoamine oxidase B inhibitor. BMC Compl. Altern Med. 2017, 17, 34. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.M.; Lee, C.; Nam, S.J.; Kim, H. Chromenone Derivatives as Monoamine Oxidase Inhibitors from Marine-Derived MAR4 Clade Streptomyces sp. CNQ-031. J. Microbiol. Biotechnol. 2021, 31, 1022–1027. [Google Scholar] [CrossRef]
- Prajapati, R.; Park, S.E.; Park, H.J.; Jung, H.A.; Choi, J.S. Identification of a potent and selective human monoamine oxidase A inhibitor, glycitein, an isoflavone isolated from Pueraria lobata flowers. ACS Food Sci. Technol. 2021, 1, 538–550. [Google Scholar] [CrossRef]
- Mohamed, E.I.; Zaki, M.A.; Chaurasiya, N.D.; Owis, A.I.; AbouZid, S.; Wang, Y.H.; Avula, B.; Seida, A.A.; Tekwani, B.L.; Ross, S.A. Monoamine oxidases inhibitors from Colvillea racemosa: Isolation, biological evaluation, and computational study. Fitoterapia 2018, 124, 217–223. [Google Scholar] [CrossRef]
- Minders, C.; Petzer, J.P.; Petzer, A.; Lourens, A.C. Monoamine oxidase inhibitory activities of heterocyclic chalcones. Bioorg. Med. Chem. Lett. 2015, 25, 5270–5276. [Google Scholar] [CrossRef] [PubMed]
- Guglielmi, P.; Mathew, B.; Secci, D.; Carradori, S. Chalcones: Unearthing their therapeutic possibility as monoamine oxidase B inhibitors. Eur. J. Med. Chem. 2020, 205, 112650. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.C.; Lin, R.D.; Chen, C.T.; Lee, M.H. Monoamine oxidase B (MAO-B) inhibition by active principles from Uncaria rhynchophylla. J. Ethnopharmacol. 2005, 100, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Recalde-Gil, M.A.; Klein-Junior, L.C.; Passos, C.D.S.; Salton, J.; Bordignon, S.A.L.; Monace, F.D.; Cechinel, V.; Teresinha Henriquesa, A. Monoamine Oxidase Inhibitory Activity of Biflavonoids from Branches of Garcinia gardneriana (Clusiaceae). Nat. Prod. Commun. 2017, 12, 505–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haraguchi, H.; Tanaka, Y.; Kabbash, A.; Fujioka, T.; Ishizu, T.; Yagi, A. Monoamine oxidase inhibitors from Gentiana lutea. Phytochemistry 2004, 65, 2255–2260. [Google Scholar] [CrossRef]
- Prajapati, R.; Seong, S.H.; Park, S.E.; Paudel, P.; Jung, H.A.; Choi, J.S. Isoliquiritigenin, a potent human monoamine oxidase inhibitor, modulates dopamine D1, D3, and vasopressin V1A receptors. Sci. Rep. 2021, 11, 23528. [Google Scholar] [CrossRef]
- Kim, J.H.; Son, Y.K.; Kim, G.H.; Hwang, K.H. Xanthoangelol and 4-Hydroxyderricin Are the Major Active Principles of the Inhibitory Activities against Monoamine Oxidases on Angelica keiskei K. Biomol. Ther. 2013, 21, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.C.; Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Kim, S.H.; Cho, M.L.; Oh, S.R.; Kim, H. Selective inhibition of monoamine oxidase A by hispidol. Bioorg. Med. Chem. Lett. 2018, 28, 584–588. [Google Scholar] [CrossRef]
- Stefanachi, A.; Leonetti, F.; Pisani, L.; Catto, M.; Carotti, A. Coumarin: A Natural, Privileged and Versatile Scaffold for Bioactive Compounds. Molecules 2018, 23, 250. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Cruz-Martins, N.; Lopez-Jornet, P.; Lopez, E.P.; Harun, N.; Yeskaliyeva, B.; Beyatli, A.; Sytar, O.; Shaheen, S.; Sharopov, F.; et al. Natural Coumarins: Exploring the Pharmacological Complexity and Underlying Molecular Mechanisms. Oxid. Med. Cell Longev. 2021, 2021, 6492346. [Google Scholar] [CrossRef]
- Rauhamaki, S.; Postila, P.A.; Niinivehmas, S.; Kortet, S.; Schildt, E.; Pasanen, M.; Manivannan, E.; Ahinko, M.; Koskimies, P.; Nyberg, N.; et al. Structure-Activity Relationship Analysis of 3-Phenylcoumarin-Based Monoamine Oxidase B Inhibitors. Front. Chem. 2018, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyiparambath, V.P.; Prayaga Rajappan, K.; Rangarajan, T.M.; Al-Sehemi, A.G.; Pannipara, M.; Bhaskar, V.; Nair, A.S.; Sudevan, S.T.; Kumar, S.; Mathew, B. Deciphering the detailed structure-activity relationship of coumarins as Monoamine oxidase enzyme inhibitors-An updated review. Chem. Biol. Drug. Des. 2021, 98, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Hossain, C.F.; Okuyama, E.; Yamazaki, M. A new series of coumarin derivatives having monoamine oxidase inhibitory activity from Monascus anka. Chem. Pharm. Bull. 1996, 44, 1535–1539. [Google Scholar] [CrossRef] [Green Version]
- Huong, D.T.; Choi, H.C.; Rho, T.C.; Lee, H.S.; Lee, M.K.; Kim, Y.H. Inhibitory activity of monoamine oxidase by coumarins from Peucedanum japonicum. Arch. Pharm. Res. 1999, 22, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.S.; Lee, I.K.; Ryoo, I.J.; Yoo, I.D. Coumarins with monoamine oxidase inhibitory activity and antioxidative coumarino-lignans from Hibiscus syriacus. J. Nat. Prod. 2001, 64, 1238–1240. [Google Scholar] [CrossRef]
- Carotti, A.; Carrieri, A.; Chimichi, S.; Boccalini, M.; Cosimelli, B.; Gnerre, C.; Carotti, A.; Carrupt, P.A.; Testa, B. Natural and synthetic geiparvarins are strong and selective MAO-B inhibitors. Synthesis and SAR studies. Bioorg. Med. Chem. Lett. 2002, 12, 3551–3555. [Google Scholar] [CrossRef]
- Jeong, S.H.; Han, X.H.; Hong, S.S.; Hwang, J.S.; Hwang, J.H.; Lee, D.; Lee, M.K.; Ro, J.S.; Hwang, B.Y. Monoamine oxidase inhibitory coumarins from the aerial parts of Dictamnus albus. Arch. Pharm. Res. 2006, 29, 1119–1124. [Google Scholar] [CrossRef]
- Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Lee, H.; Shin, H.M.; Oh, S.R.; Kim, H. Potent inhibition of monoamine oxidase A by decursin from Angelica gigas Nakai and by wogonin from Scutellaria baicalensis Georgi. Int. J. Biol. Macromol. 2017, 97, 598–605. [Google Scholar] [CrossRef]
- Wang, Y.S.; Li, B.T.; Liu, S.X.; Wen, Z.Q.; Yang, J.H.; Zhang, H.B.; Hao, X.J. Anisucoumaramide, a Bioactive Coumarin from Clausena anisum-olens. J. Nat. Prod. 2017, 80, 798–804. [Google Scholar] [CrossRef]
- Seong, S.H.; Ali, M.Y.; Jung, H.A.; Choi, J.S. Umbelliferone derivatives exert neuroprotective effects by inhibiting monoamine oxidase A, self-amyloidbeta aggregation, and lipid peroxidation. Bioorg. Chem. 2019, 92, 103293. [Google Scholar] [CrossRef]
- Farina, R.; Pisani, L.; Catto, M.; Nicolotti, O.; Gadaleta, D.; Denora, N.; Soto-Otero, R.; Mendez-Alvarez, E.; Passos, C.S.; Muncipinto, G.; et al. Structure-Based Design and Optimization of Multitarget-Directed 2H-Chromen-2-one Derivatives as Potent Inhibitors of Monoamine Oxidase B and Cholinesterases. J. Med. Chem. 2015, 58, 5561–5578. [Google Scholar] [CrossRef] [PubMed]
- Rehuman, N.A.; Oh, J.M.; Nath, L.R.; Khames, A.; Abdelgawad, M.A.; Gambacorta, N.; Nicolotti, O.; Jat, R.K.; Kim, H.; Mathew, B. Halogenated Coumarin-Chalcones as Multifunctional Monoamine Oxidase-B and Butyrylcholinesterase Inhibitors. ACS Omega 2021, 6, 28182–28193. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Al-Taweel, A.M. Phenolic Compounds from the Natural Sources and Their Cytotoxicity. In Phenolic Compounds—Natural Sources, Importance and Applications; Soto-Hernandez, M., Palma-Tenango, M., Garcia-Mateos, M.d.R., Eds.; IntechOpen: London, UK, 2017. [Google Scholar]
- Han, Y.S.; Van der Heijden, R.; Verpoorte, R. Biosynthesis of anthraquinones in cell cultures of the Rubiaceae. Plant Cell Tissue Organ. Cult. 2001, 67, 201–220. [Google Scholar] [CrossRef]
- El-Awaad, I.; Bocola, M.; Beuerle, T.; Liu, B.; Beerhues, L. Bifunctional CYP81AA proteins catalyse identical hydroxylations but alternative regioselective phenol couplings in plant xanthone biosynthesis. Nat. Commun. 2016, 7, 11472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, Y.; Gotfredsen, C.H.; Larsen, T.O. Genetic Characterization of Neosartorin Biosynthesis Provides Insight into Heterodimeric Natural Product Generation. Org. Lett. 2018, 20, 7197–7200. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y.; Wu, H.; Yuan, M.; Zheng, C.; Xu, H. Xanthone Glucosides: Isolation, Bioactivity and Synthesis. Molecules 2021, 26, 5575. [Google Scholar] [CrossRef]
- Ruan, J.; Zheng, C.; Liu, Y.; Qu, L.; Yu, H.; Han, L.; Zhang, Y.; Wang, T. Chemical and Biological Research on Herbal Medicines Rich in Xanthones. Molecules 2017, 22, 1698. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, O.; Katsumata, Y.; Oya, M.; Chari, V.M.; Vermes, B.; Wagner, H.; Hostettmann, K. Inhibition of type A and type B monoamine oxidases by naturally occurring xanthones. Planta Med. 1981, 42, 17–21. [Google Scholar] [CrossRef]
- Schaufelberger, D.; Hostettmann, K. Chemistry and pharmacology of Gentiana lactea. Planta Med. 1988, 54, 219–221. [Google Scholar] [CrossRef]
- Rocha, L.; Marston, A.; Kaplan, M.A.; Stoeckli-Evans, H.; Thull, U.; Testa, B.; Hostettmann, K. An antifungal gamma-pyrone and xanthones with monoamine oxidase inhibitory activity from Hypericum brasiliense. Phytochemistry 1994, 36, 1381–1385. [Google Scholar] [CrossRef]
- Gnerre, C.; Thull, U.; Gaillard, P.; Carrupt, P.A.; Testa, B.; Fernandes, E.; Cruciani, G. Natural and synthetic xanthones as monoamine oxidase inhibitors: Biological assay and 3D-QSAR. Helv. Chim. Acta 2001, 84, 552–570. [Google Scholar] [CrossRef]
- Wolfender, J.L.; Hamburger, M.; Msonthi, J.D.; Hostettmann, K. Xanthones from Chironia krebsii. Phytochemistry 1991, 30, 3625–3629. [Google Scholar] [CrossRef]
- Wairata, J.; Sukandar, E.R.; Fadlan, A.; Purnomo, A.S.; Taher, M.; Ersam, T. Evaluation of the Antioxidant, Antidiabetic, and Antiplasmodial Activities of Xanthones Isolated from Garcinia forbesii and Their In Silico Studies. Biomedicines 2021, 9, 1380. [Google Scholar] [CrossRef]
- Yang, J.L.; Liu, L.L.; Shi, Y.P. Phytochemicals and biological activities of Gentiana species. Nat. Prod. Commun. 2010, 5, 649–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomic, M.; Tovilovic, G.; Butorovic, B.; Krstic, D.; Jankovic, T.; Aljancic, I.; Menkovic, N. Neuropharmacological evaluation of diethylether extract and xanthones of Gentiana kochiana. Pharmacol. Biochem. Behav. 2005, 81, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Hong, S.S.; Han, X.H.; Hwang, J.S.; Lee, D.; Lee, H.; Yun, Y.P.; Kim, Y.; Ro, J.S.; Hwang, B.Y. Prenylated xanthones from the root bark of Cudrania tricuspidata. J. Nat. Prod. 2007, 70, 1207–1209. [Google Scholar] [CrossRef]
- Urbain, A.; Marston, A.; Grilo, L.S.; Bravo, J.; Purev, O.; Purevsuren, B.; Batsuren, D.; Reist, M.; Carrupt, P.A.; Hostettmann, K. Xanthones from Gentianella amarella ssp. acuta with acetylcholinesterase and monoamine oxidase inhibitory activities. J. Nat. Prod. 2008, 71, 895–897. [Google Scholar] [CrossRef]
- Dimitrov, M.; Nikolova, I.; Benbasat, N.; Kitanov, G.; Danchev, N. Acute toxicity, antidepressive and MAO inhibitory ac-tivity of mangiferin isolated from Hypericum aucheri. Biothechnol. Biotechnol. Equip. 2011, 25, 2668–2671. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, M.; Satoh, Y.; Maebayashi, Y.; Horie, Y. Monoamine oxidase inhibitors from a fungus, Emericella navahoensis. Chem. Pharm. Bull. 1988, 36, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Andujar, I.; Recio, M.C.; Giner, R.M.; Rios, J.L. Traditional chinese medicine remedy to jury: The pharmacological basis for the use of shikonin as an anticancer therapy. Curr. Med. Chem. 2013, 20, 2892–2898. [Google Scholar] [CrossRef]
- Choi, W.H.; Hong, S.S.; Lee, S.A.; Han, X.H.; Lee, K.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Monoamine oxidase inhibitory naphthoquinones from the roots of Lithospermum erythrorhizon. Arch. Pharm. Res. 2005, 28, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.A.; Steyn, S.; Castagnoli, N., Jr. Isolation and characterization of a monoamine oxidase inhibitor from tobacco leaves. Chem. Res. Toxicol. 2000, 13, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Jaka, O.; Iturria, I.; van der Toorn, M.; Hurtado de Mendoza, J.; Latino, D.; Alzualde, A.; Peitsch, M.C.; Hoeng, J.; Koshibu, K. Effects of Natural Monoamine Oxidase Inhibitors on Anxiety-Like Behavior in Zebrafish. Front. Pharmacol. 2021, 12, 669370. [Google Scholar] [CrossRef] [PubMed]
- Coelho Cerqueira, E.; Netz, P.A.; Diniz, C.; Petry do Canto, V.; Follmer, C. Molecular insights into human monoamine oxidase (MAO) inhibition by 1,4-naphthoquinone: Evidences for menadione (vitamin K3) acting as a competitive and reversible inhibitor of MAO. Bioorg. Med. Chem. 2011, 19, 7416–7424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostert, S.; Petzer, A.; Petzer, J.P. Evaluation of Natural and Synthetic 1,4-naphthoquinones as Inhibitors of Monoamine Oxidase. Chem. Biol. Drug Des. 2016, 87, 737–746. [Google Scholar] [CrossRef]
- Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Oh, S.R.; Kim, H. Selective inhibition of monoamine oxidase A by purpurin, an anthraquinone. Bioorg. Med. Chem. Lett. 2017, 27, 1136–1140. [Google Scholar] [CrossRef]
- Paudel, P.; Seong, S.H.; Shrestha, S.; Jung, H.A.; Choi, J.S. In Vitro and in Silico Human Monoamine Oxidase Inhibitory Potential of Anthraquinones, Naphthopyrones, and Naphthalenic Lactones from Cassia obtusifolia Linn Seeds. ACS Omega 2019, 4, 16139–16152. [Google Scholar] [CrossRef] [Green Version]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and phenylalanine biosynthesis in the green lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Dhiman, P.; Malik, N.; Khatkar, A. Hybrid caffeic acid derivatives as monoamine oxidases inhibitors: Synthesis, radical scavenging activity, molecular docking studies and in silico ADMET analysis. Chem. Cent. J. 2018, 12, 112. [Google Scholar] [CrossRef]
- Mei, Y.; Pan, D.; Jiang, Y.; Zhang, W.; Yao, X.; Dai, Y.; Yu, Y.; Yao, X. Target discovery of chlorogenic acid derivatives from the flower buds of Lonicera macranthoides and their MAO B inhibitory mechanism. Fitoterapia 2019, 134, 297–304. [Google Scholar] [CrossRef]
- Wu, G.F.; Jiang, X.L.; Gong, Y.Z.; Hu, Y.D.; Bai, X.L.; Liao, X. Ligand fishing of anti-neurodegenerative components from Lonicera japonica using magnetic nanoparticles immobilised with monoamine oxidase B. J. Sep. Sci. 2019, 42, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Meng, X.; Li, S.; Gan, R.Y.; Li, Y.; Li, H.B. Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients 2018, 10, 1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajeswari, A.; Sabesan, M. Inhibition of monoamine oxidase-B by the polyphenolic compound, curcumin and its metabolite tetrahydrocurcumin, in a model of Parkinson’s disease induced by MPTP neurodegeneration in mice. Inflammopharmacology 2008, 16, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.C.; Choi, B.; Nam, S.J.; Kim, H. Inhibition of monoamine oxidase A and B by demethoxycurcumin and bisde-methoxycurcumin. J. Appl. Biol. Chem. 2018, 61, 187–190. [Google Scholar] [CrossRef]
- Arvizu-Espinosa, M.G.; von Poser, G.L.; Henriques, A.T.; Mendoza-Ruiz, A.; Cardador-Martinez, A.; Gesto-Borroto, R.; Nunez-Aragon, P.N.; Villarreal-Ortega, M.L.; Sharma, A.; Cardoso-Taketa, A. Bioactive Dimeric Acylphloroglucinols from the Mexican Fern Elaphoglossum paleaceum. J. Nat. Prod. 2019, 82, 785–791. [Google Scholar] [CrossRef]
- Perveen, S. Introductory Chapter: Terpenes and Terpenoids in Terpenes and Terpenoids; Perveen, S., Al-Taweel, A., Eds.; IntechOpen: London, UK, 2018; p. 79683. [Google Scholar]
- Dittmann, K.; Gerhauser, C.; Klimo, K.; Hamburger, M. HPLC-based activity profiling of Salvia miltiorrhiza for MAO A and iNOS inhibitory activities. Planta Med. 2004, 70, 909–913. [Google Scholar] [CrossRef]
- Subaraja, M.; Vanisree, A.J. The novel phytocomponent asiaticoside-D isolated from Centella asiatica exhibits monoamine oxidase-B inhibiting potential in the rotenone degenerated cerebral ganglions of Lumbricus terrestris. Phytomedicine 2019, 58, 152833. [Google Scholar] [CrossRef]
- Gaston, R., Jr.; Geldenhuys, W.J.; Dudley, G.B. Synthesis of Illudinine from Dimedone and Identification of Activity as a Monoamine Oxidase Inhibitor. J. Org. Chem. 2020, 85, 13429–13437. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Koch, W.; Czernicka, L.; Glowniak, K.; Asakawa, Y.; Umeyama, A.; Marzec, Z.; Kuzuhara, T. MAO-A Inhibitory Potential of Terpene Constituents from Ginger Rhizomes-A Bioactivity Guided Fractionation. Molecules 2018, 23, 1301. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.Y.; Li, H.J.; Li, Q.Y.; Wu, Y.C. Application of marine natural products in drug research. Bioorg. Med. Chem. 2021, 35, 116058. [Google Scholar] [CrossRef] [PubMed]
- Hong, A.; Tu, L.C.; Yang, I.; Lim, K.M.; Nam, S.J. Marine natural products with monoamine oxidase (MAO) inhibitory activity. Pharm. Biol. 2020, 58, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Roy, A.; Jung, J.H.; Choi, J.S. Evaluation of the inhibitory effects of eckol and dieckol isolated from edible brown alga Eisenia bicyclis on human monoamine oxidases A and B. Arch. Pharm. Res. 2017, 40, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Paudel, P.; Choi, J.W.; Ahn, D.H.; Nam, T.J.; Jung, H.A.; Choi, J.S. Probing Multi-Target Action of Phlorotannins as New Monoamine Oxidase Inhibitors and Dopaminergic Receptor Modulators with the Potential for Treatment of Neuronal Disorders. Mar. Drugs 2019, 17, 377. [Google Scholar] [CrossRef] [Green Version]
- Paudel, P.; Park, S.E.; Seong, S.H.; Jung, H.A.; Choi, J.S. Bromophenols from Symphyocladia latiuscula Target Human Monoamine Oxidase and Dopaminergic Receptors for the Management of Neurodegenerative Diseases. J. Agric. Food Chem. 2020, 68, 2426–2436. [Google Scholar] [CrossRef]
- Mikell, J.R.; Schaneberg, B.T.; Khan, I.A. Isolation and purification of Kava lactones by high performance centrifugal partition chromatography. J. Liq. Chromatogr. Relat. Technol. 2003, 26, 3069–3074. [Google Scholar] [CrossRef]
- Aporosa, S.A. De-mythologizing and re-branding of kava as the new world drug of choice. Drug Sci. Pol. Law 2019, 5, 1–13. [Google Scholar] [CrossRef]
- Prinsloo, D.; van Dyk, S.; Petzer, A.; Petzer, J.P. Monoamine Oxidase Inhibition by Kavalactones from Kava (Piper Methysticum). Planta Med. 2019, 85, 1136–1142. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.S.; Lee, E.Y.; Kang, M.G.; Nam, S.J.; Park, D.; Kim, H. (S)-5-Methymellein isolated from an endogenous lichen fungus Rosellinia corticium as a potent inhibitor of human monoamine oxidase A. Processes 2022, 10, 166. [Google Scholar] [CrossRef]
- Huang, C.; Xiong, J.; Guan, H.D.; Wang, C.H.; Lei, X.; Hu, J.F. Discovery, synthesis, biological evaluation and molecular docking study of (R)-5-methylmellein and its analogs as selective monoamine oxidase A inhibitors. Bioorg. Med. Chem. 2019, 27, 2027–2040. [Google Scholar] [CrossRef]
- Jeong, G.S.; Hillman, P.F.; Kang, M.G.; Hwang, S.; Park, J.E.; Nam, S.J.; Park, D.; Kim, H. Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus. J. Fungi 2021, 7, 876. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Chaurasiya, N.D.; Tekwani, B.L.; Doerksen, R.J. Interactions of endocannabinoid virodhamine and related analogs with human monoamine oxidase-A and -B. Biochem. Pharmacol. 2018, 155, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, J.C.; Ugun-Klusek, A.; Allen, G.; De Girolamo, L.A.; Hargreaves, I.; Ufer, C.; Abramov, A.Y.; Billett, E.E. Monoamine oxidase-A knockdown in human neuroblastoma cells reveals protection against mitochondrial toxins. FASEB J. 2014, 28, 218–229. [Google Scholar] [CrossRef]
- Wang, X.; Li, B.; Kim, Y.J.; Wang, Y.C.; Li, Z.; Yu, J.; Zeng, S.; Ma, X.; Choi, I.Y.; Di Biase, S.; et al. Targeting monoamine oxidase A for T cell-based cancer immunotherapy. Sci. Immunol. 2021, 6, 2383. [Google Scholar] [CrossRef]
- Aljanabi, R.; Alsous, L.; Sabbah, D.A.; Gul, H.I.; Gul, M.; Bardaweel, S.K. Monoamine Oxidase (MAO) as a Potential Target for Anticancer Drug Design and Development. Molecules 2021, 26, 6019. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Kapur, G.; Baracco, R.; Valentini, R.P.; Mattoo, T.K.; Jain, A. Treatment of Hypertension in Children With Catecholamine-Secreting Tumors: A Systematic Approach. J. Clin. Hypertens. 2015, 17, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Shitara, T.; Hatakeyama, S.I.; Suzuki, N.; Maruyama, K.; Kobayashi, T.; Tsuchida, Y. An infant with systemic hypertension, renal artery stenosis, and neuroblastoma. J. Pediatr. Surg. 2004, 39, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.B. Childhood neuroblastoma masquerading as pheochromocytoma: Case report. Int. Med. Case Rep. J. 2016, 9, 65–67. [Google Scholar] [CrossRef] [Green Version]
- Harding, M.; Deyell, R.J.; Blydt-Hansen, T. Catecholamines in neuroblastoma: Driver of hypertension, or solely a marker of disease? Cancer Rep. 2021, e1569. [Google Scholar] [CrossRef]
- Laug, W.E.; Siegel, S.E.; Shaw, K.N.; Landing, B.; Baptista, J.; Gutenstein, M. Initial urinary catecholamine metabolite concentrations and prognosis in neuroblastoma. Pediatrics 1978, 62, 77–83. [Google Scholar] [CrossRef]
- Liu, Y.L.; Cheng, A.T.; Chen, H.R.; Hsu, Y.P. Simultaneous HPLC of twelve monoamines and metabolites shows neuroblastoma cell line releases HVA and HIAA. Biomed. Chromatogr. 2000, 14, 544–548. [Google Scholar] [CrossRef]
- Ugun-Klusek, A.; Theodosi, T.S.; Fitzgerald, J.C.; Burte, F.; Ufer, C.; Boocock, D.J.; Yu-Wai-Man, P.; Bedford, L.; Billett, E.E. Monoamine oxidase-A promotes protective autophagy in human SH-SY5Y neuroblastoma cells through Bcl-2 phosphorylation. Redox. Biol. 2019, 20, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Igosheva, N.; Lorz, C.; O’Conner, E.; Glover, V.; Mehmet, H. Isatin, an endogenous monoamine oxidase inhibitor, triggers a dose- and time-dependent switch from apoptosis to necrosis in human neuroblastoma cells. Neurochem. Int. 2005, 47, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Primers 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Himpel, S.; Panzer, P.; Eirmbter, K.; Czajkowska, H.; Sayed, M.; Packman, L.C.; Blundell, T.; Kentrup, H.; Grotzinger, J.; Joost, H.G.; et al. Identification of the autophosphorylation sites and characterization of their effects in the protein kinase DYRK1A. Biochem. J. 2001, 359, 497–505. [Google Scholar] [CrossRef]
- Uhl, K.L.; Schultz, C.R.; Geerts, D.; Bachmann, A.S. Harmine, a dual-specificity tyrosine phosphorylation-regulated kinase (DYRK) inhibitor induces caspase-mediated apoptosis in neuroblastoma. Cancer Cell Int. 2018, 18, 82. [Google Scholar] [CrossRef]
- Gockler, N.; Jofre, G.; Papadopoulos, C.; Soppa, U.; Tejedor, F.J.; Becker, W. Harmine specifically inhibits protein kinase DYRK1A and interferes with neurite formation. FEBS J. 2009, 276, 6324–6337. [Google Scholar] [CrossRef] [Green Version]
- Uezono, T.; Maruyama, W.; Matsubara, K.; Naoi, M.; Shimizu, K.; Saito, O.; Ogawa, K.; Mizukami, H.; Hayase, N.; Shiono, H. Norharman, an indoleamine-derived beta-carboline, but not Trp-P-2, a gamma-carboline, induces apoptotic cell death in human neuroblastoma SH-SY5Y cells. J. Neural Transm. 2001, 108, 943–953. [Google Scholar] [CrossRef]
- Naveen, C.R.; Gaikwad, S.; Agrawal-Rajput, R. Berberine induces neuronal differentiation through inhibition of cancer stemness and epithelial-mesenchymal transition in neuroblastoma cells. Phytomedicine 2016, 23, 736–744. [Google Scholar] [CrossRef]
- Calvani, M.; Subbiani, A.; Bruno, G.; Favre, C. Beta-Blockers and Berberine: A Possible Dual Approach to Contrast Neuroblastoma Growth and Progression. Oxid. Med. Cell Longev. 2020, 7, 534–693. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaurasiya, N.D.; Leon, F.; Muhammad, I.; Tekwani, B.L. Natural Products Inhibitors of Monoamine Oxidases—Potential New Drug Leads for Neuroprotection, Neurological Disorders, and Neuroblastoma. Molecules 2022, 27, 4297. https://doi.org/10.3390/molecules27134297
Chaurasiya ND, Leon F, Muhammad I, Tekwani BL. Natural Products Inhibitors of Monoamine Oxidases—Potential New Drug Leads for Neuroprotection, Neurological Disorders, and Neuroblastoma. Molecules. 2022; 27(13):4297. https://doi.org/10.3390/molecules27134297
Chicago/Turabian StyleChaurasiya, Narayan D., Francisco Leon, Ilias Muhammad, and Babu L. Tekwani. 2022. "Natural Products Inhibitors of Monoamine Oxidases—Potential New Drug Leads for Neuroprotection, Neurological Disorders, and Neuroblastoma" Molecules 27, no. 13: 4297. https://doi.org/10.3390/molecules27134297