
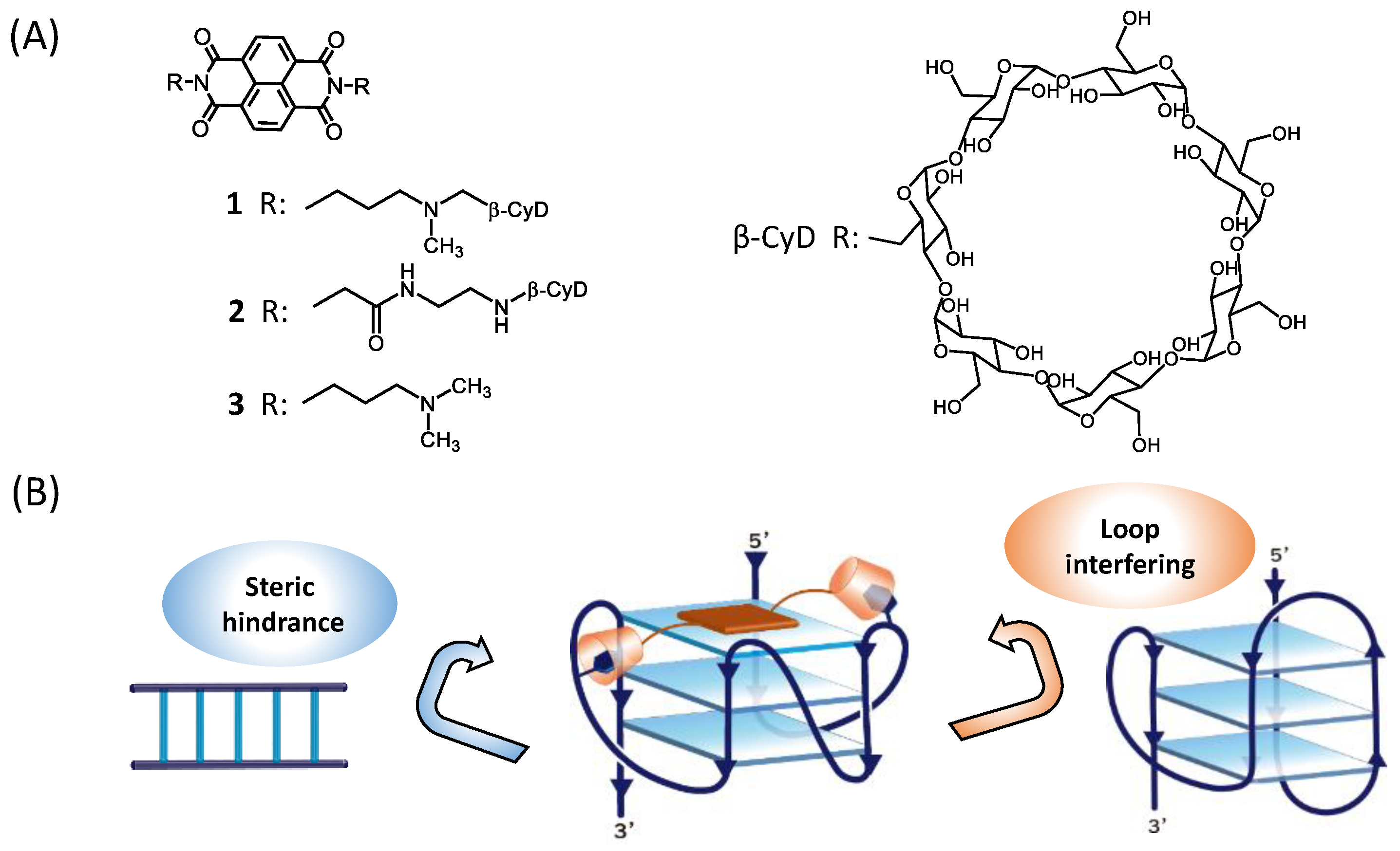
Naphthalene Diimides Carrying Two β-Cyclodextrins Prefer Telomere RNA G-Quadruplex Recognition

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
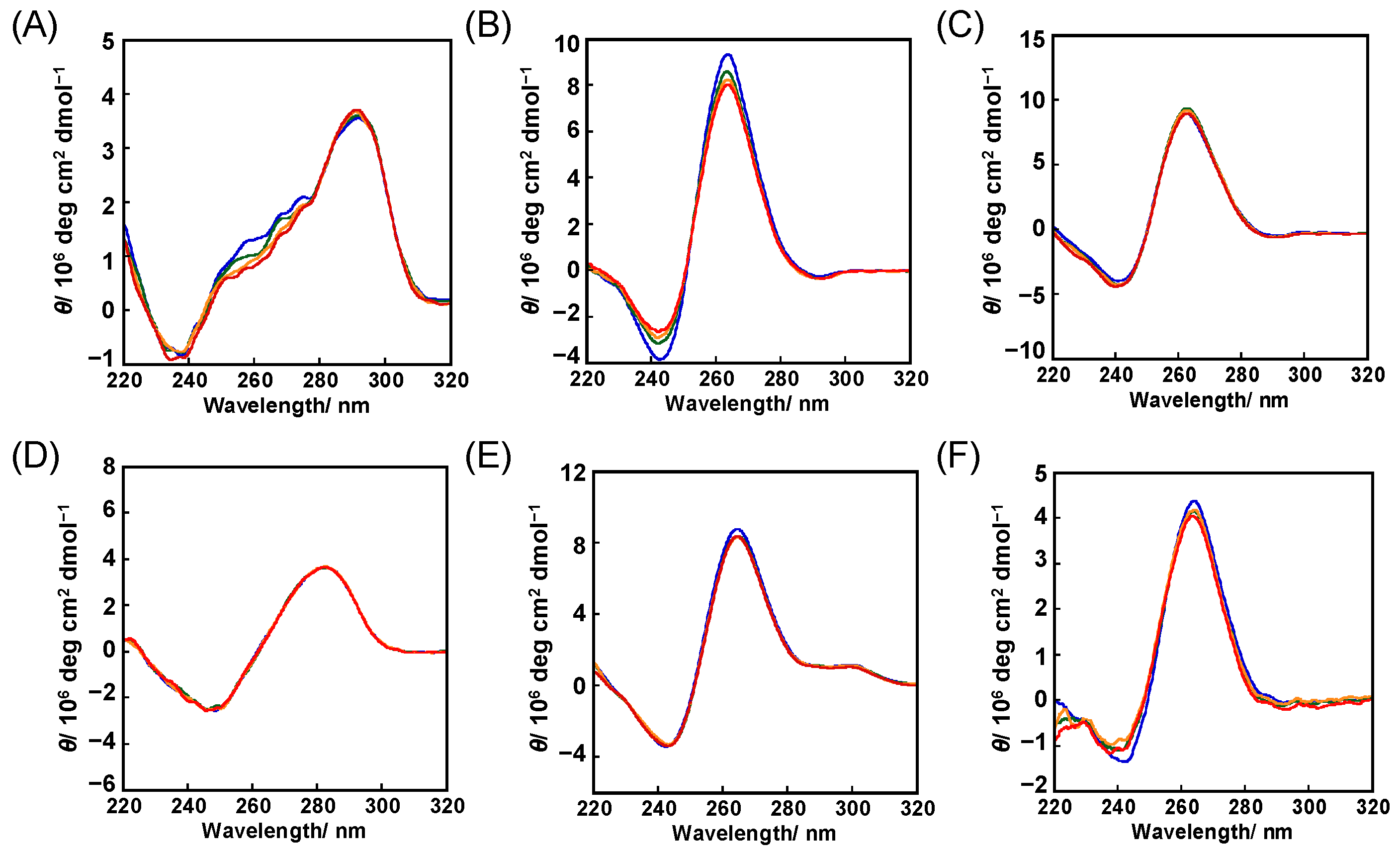
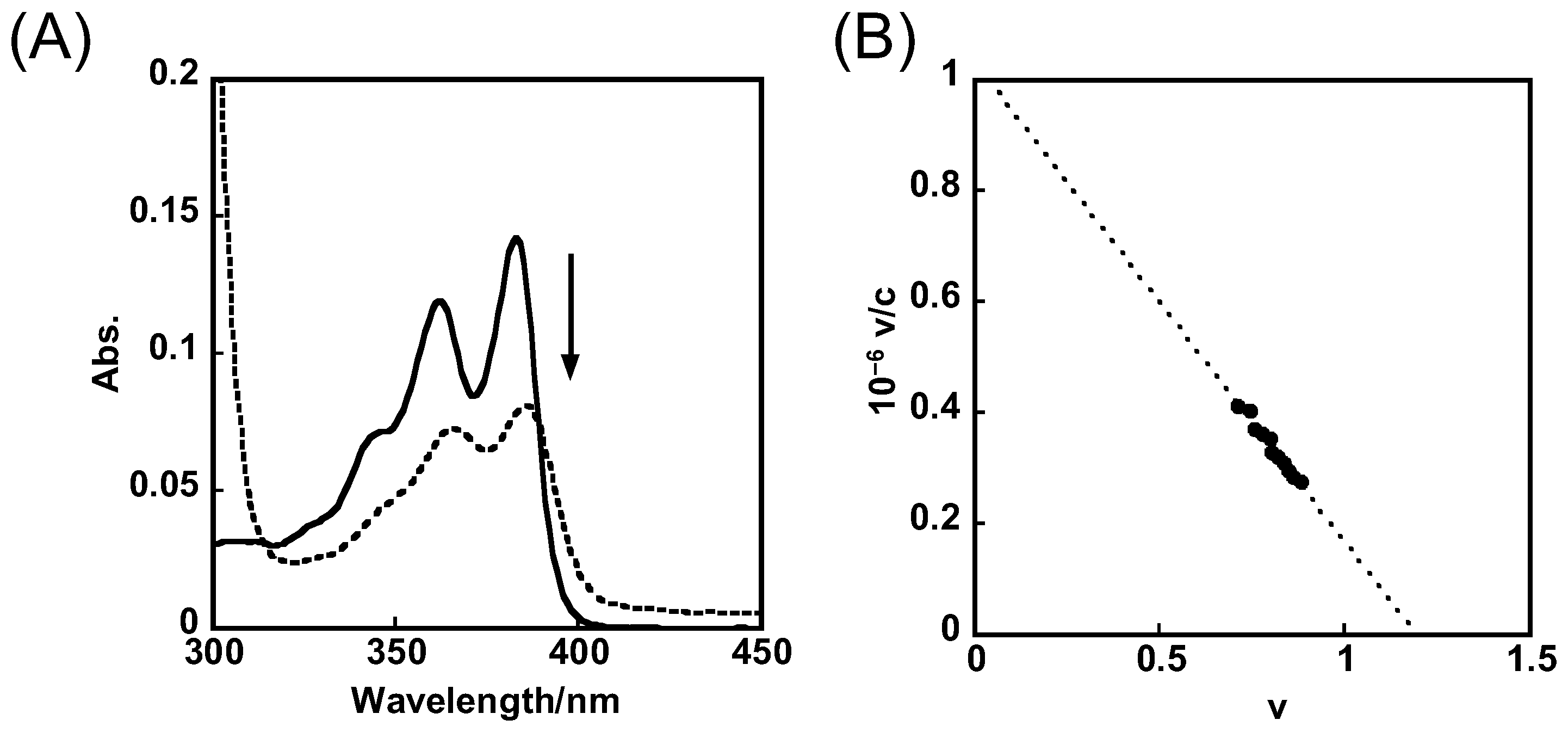
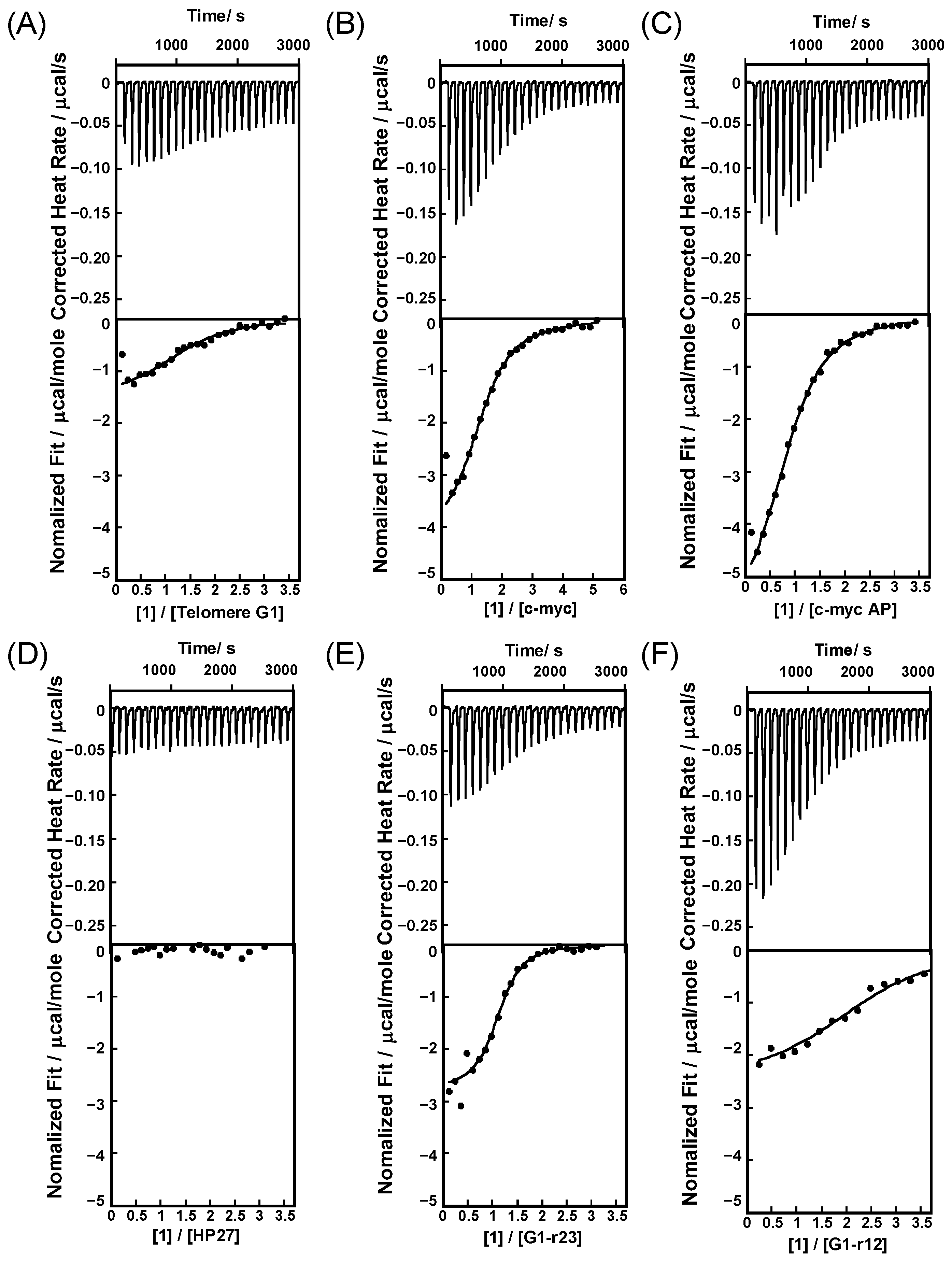
2.1. Binding Behaviors of 1 and 2 with DNA or RNA G-Quadruplex
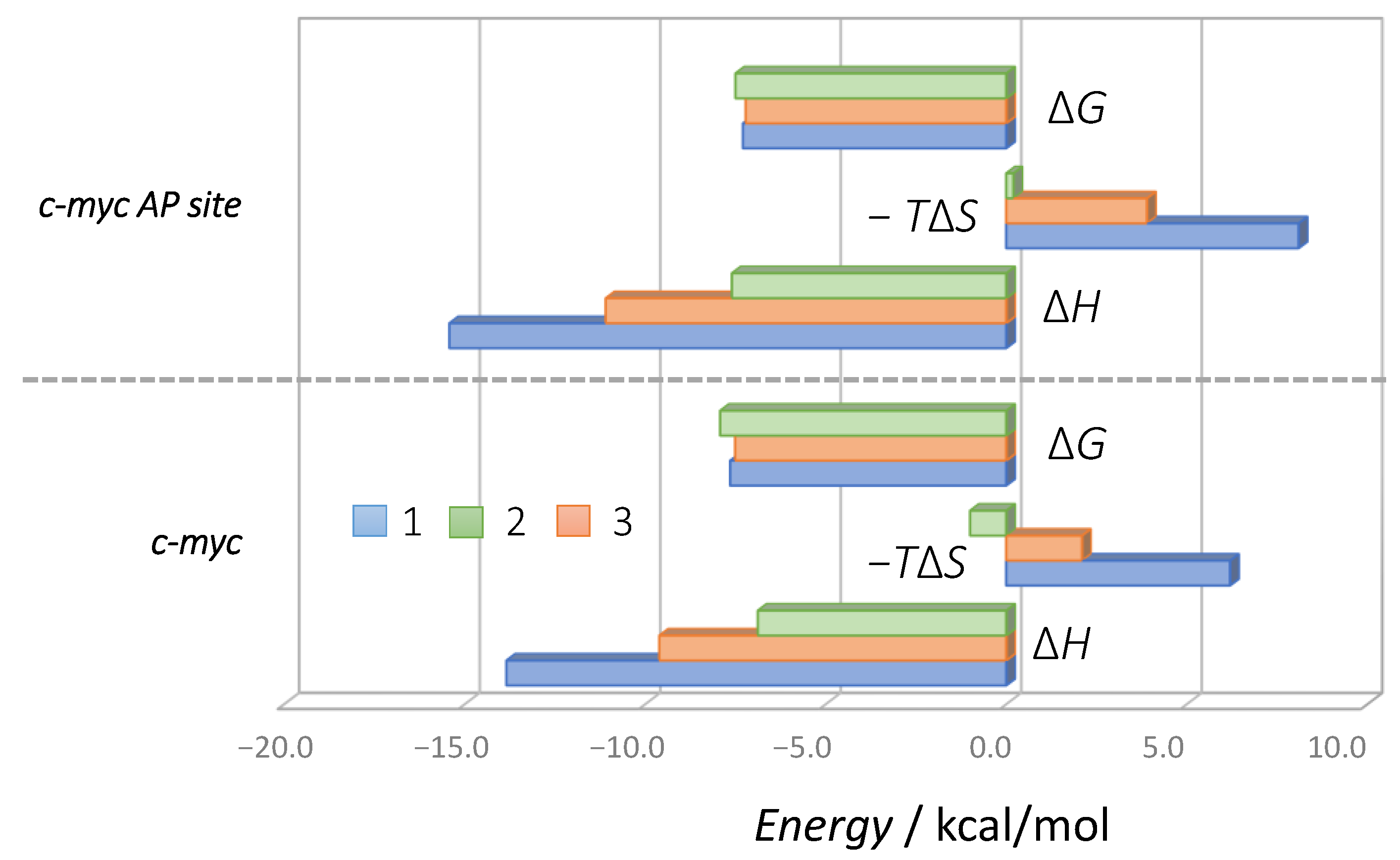
2.2. Binding Model of 1 and 2 with DNA or RNA G-Quadruplex
3. Materials and Methods
3.1. Generals
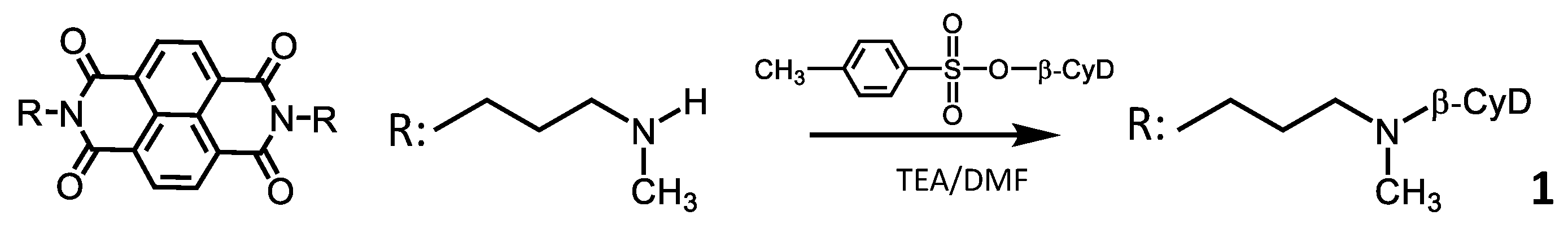
3.2. Synthesis of 1
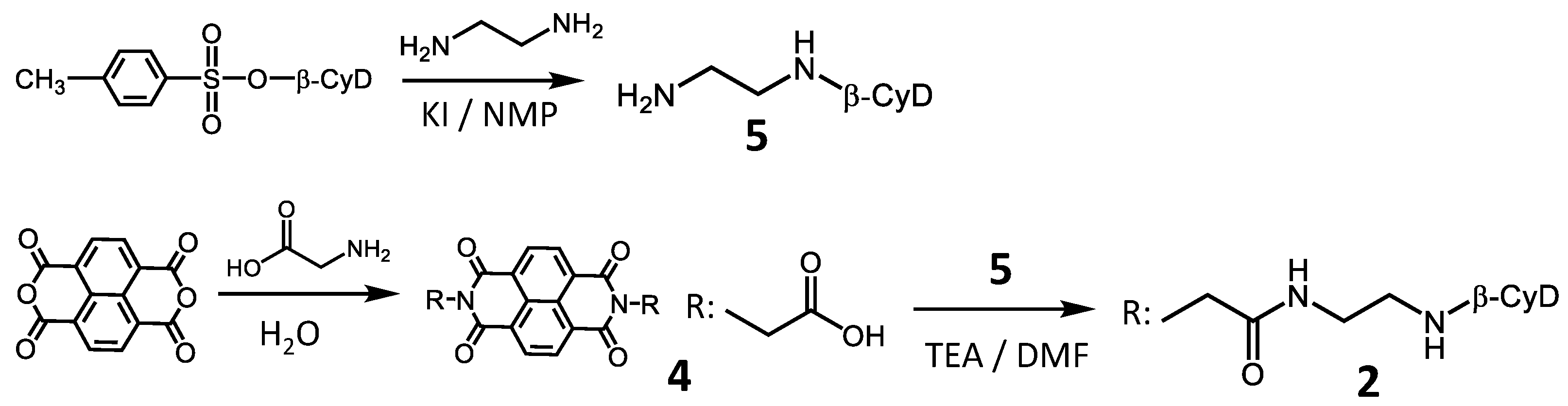
3.3. Synthesis of 2
3.4. Circular Dichroism (CD) Measurements
3.5. Isothermal Titration Calorimetry (ITC) Measurements
3.6. UV-Vis Absorption Spectroscopy
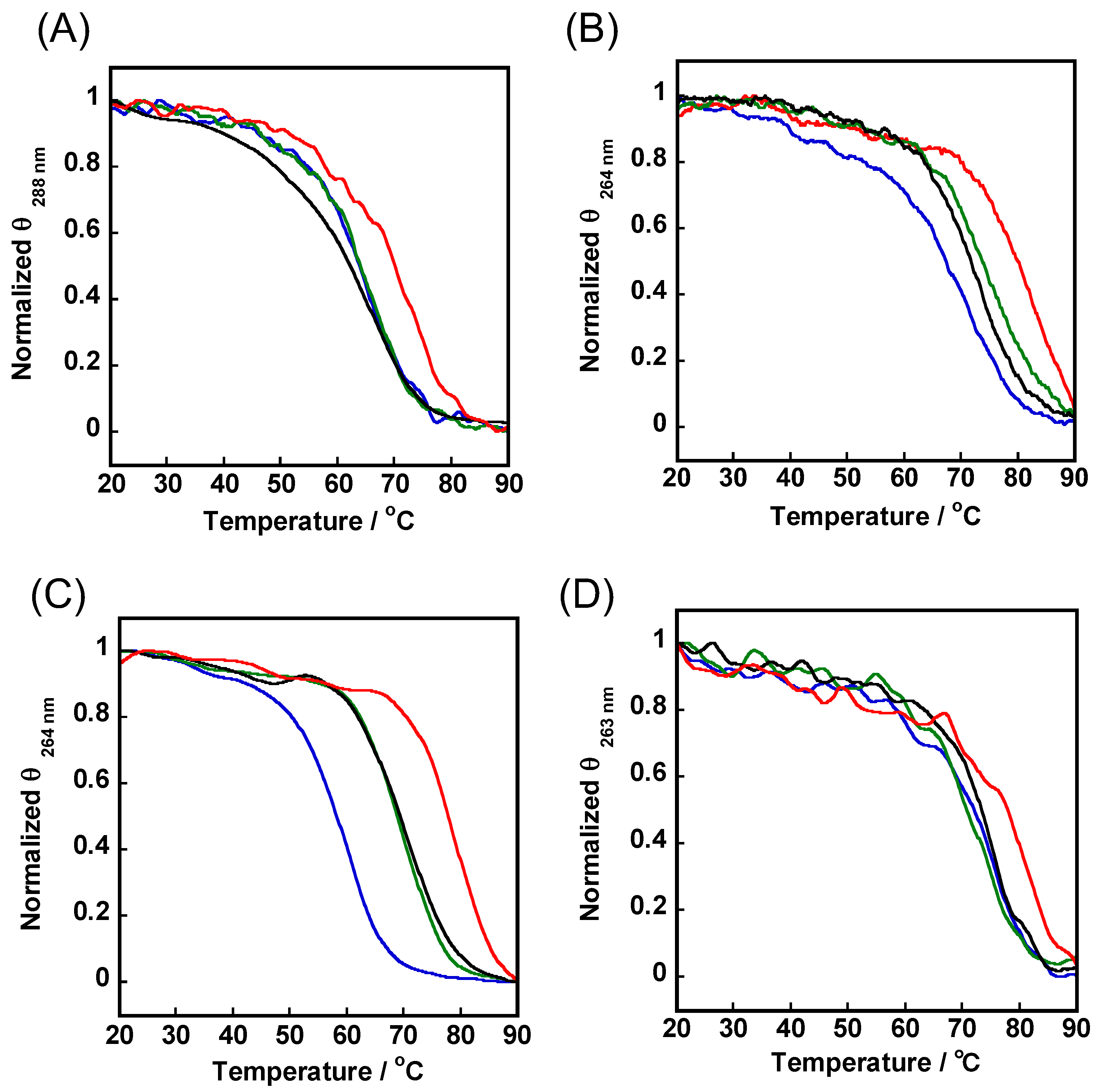
3.7. Apparent Melting Temperature (Tm) Detection
3.8. Modeling Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dolinnaya, N.G.; Ogloblina, A.M.; Yakubovskaya, M.G. Structure, Properties, and Biological Relevance of the DNA and RNA G-Quadruplexes: Overview 50 Years after Their Discovery. Biochemistry 2016, 81, 1602–1649. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.; Lipps, H.J. G-quadruplexes and their regulatory roles in biology. Nucleic Acids Res. 2015, 43, 8627–8637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, C.K.; Merrick, C.J. G-Quadruplexes: Prediction, Characterization, and Biological Application. Trends Biotechnol. 2017, 35, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Cimino-Reale, G.; Zaffaroni, N.; Folini, M. Emerging Role of G-quadruplex DNA as Target in Anticancer Therapy. Curr. Pharm. Des. 2016, 22, 6612–6624. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.M.; Rodriguez, R. G-quadruplexes: Selective DNA targeting for cancer therapeutics? Expert Rev. Clin. Pharmacol. 2011, 4, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Luu, K.N.; Phan, A.T.; Kuryavyi, V.; Lacroix, L.; Patel, D.J. Structure of the Human Telomere in K+ Solution: An Intramolecular (3 + 1) G-Quadruplex Scaffold. J. Am. Chem. Soc. 2006, 128, 9963–9970. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Okamoto, K. Structural insights into G-quadruplexes: Towards new anticancer drugs. Future Med. Chem. 2010, 2, 619–646. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Chen, D.; Dai, J.; Jones, R.A.; Yang, D. Solution structure of the biologically relevant G-quadruplex element in the human c-MYC promoter. Implications for G-quadruplex stabilization. Biochemistry 2005, 44, 2048–2058. [Google Scholar] [CrossRef]
- Asamitsu, S.; Obata, S.; Yu, Z.; Bando, T.; Sugiyama, H. Recent Progress of Targeted G-Quadruplex-Preferred Ligands Toward Cancer Therapy. Molecules 2019, 24, 429. [Google Scholar] [CrossRef] [Green Version]
- Asamitsu, S.; Bando, T.; Sugiyama, H. Ligand Design to Acquire Specificity to Intended G-Quadruplex Structures. Chem. A Euro. J. 2019, 25, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.R.; Cadoni, E.; Ressurreicao, A.S.; Moreira, R.; Paulo, A. Design of Modular G-quadruplex Ligands. ChemMedChem 2018, 13, 869–893. [Google Scholar] [CrossRef] [PubMed]
- Garner, T.P.; Williams, H.E.; Gluszyk, K.I.; Roe, S.; Oldham, N.J.; Stevens, M.F.; Moses, J.E.; Searle, M.S. Selectivity of small molecule ligands for parallel and anti-parallel DNAG-quadruplex structures. Org. Biomol. Chem. 2009, 7, 4194–4200. [Google Scholar] [CrossRef]
- Platella, C.; Trajkovski, M.; Doria, F.; Freccero, M.; Plavec, J.; Montesarchio, D. On the interaction of an anticancer trisubstituted naphthalene diimide with G-quadruplexes of different topologies: A structural insight. Nucleic Acids Res. 2020, 48, 12380–12393. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.; Oxenford, S.; Angell, R.; Marchetti, C.; Ohnmacht, S.A.; Wilson, W.D.; Neidle, S. Substituted Naphthalenediimide Compounds Bind Selectively to Two Human Quadruplex Structures with Parallel Topology. ACS Med. Chem. Lett. 2020, 11, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zheng, Y.; Zhai, Q.; Wei, D. Recent advances in the development of small molecules targeting RNA G-quadruplexes for drug discovery. Bioorg. Chem. 2021, 110, 104804. [Google Scholar] [CrossRef] [PubMed]
- Esaki, Y.; Islam, M.M.; Fujii, S.; Sato, S.; Takenaka, S. Design of tetraplex specific ligands: Cyclic naphthalene diimide. Chem. Commun. 2014, 50, 5967–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, C.; Minarini, A.; Tumiatti, V.; Moraca, F.; Parrotta, L.; Alcaro, S.; Rigo, R.; Sissi, C.; Gunaratnam, M.; Ohnmacht, S.A.; et al. Macrocyclic naphthalene diimides as G-quadruplex binders. Bioorg. Med. Chem. 2015, 23, 3819–3830. [Google Scholar] [CrossRef]
- Collie, G.W.; Promontorio, R.; Hampel, S.M.; Micco, M.; Neidle, S.; Parkinson, G.N. Structural Basis for Telomeric G-Quadruplex Targeting by Naphthalene Diimide Ligands. J. Am. Chem. Soc. 2012, 134, 2723–2731. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Angell, R.; Oxenford, S.; Worthington, J.; Williams, N.; Barton, N.; Fowler, T.G.; O’Flynn, D.E.; Sunose, M.; McConville, M.; et al. Asymmetrically substituted quadruplex-binding naphthalene diimide showing potent activity in pancreatic cancer models. ACS Med. Chem. Lett. 2020, 11, 1634–1644. [Google Scholar] [CrossRef]
- Tumiatti, V.A.; Milelli, A.; Minarini, A.; Micco, M.; Campani, A.G.; Roncuzzi, L.; Baiocchi, D.; Marinello, J.; Capranico, G.; Zini, M.; et al. Design, Synthesis, and Biological Evaluation of Substituted Naphthalene Imides and Diimides as Anticancer Agent. J. Med. Chem. 2009, 52, 7873–7877. [Google Scholar] [CrossRef] [PubMed]
- Recagni, M.; Tassinari, M.; Doria, F.; Cimino-Reale, G.; Zaffaroni, N.; Freccero, M.; Folini, M.; Richter, S.N. The Oncogenic Signaling Pathways in BRAF-Mutant Melanoma Cells Are Modulated by Naphthalene Diimide-Like G-Quadruplex Ligands. Cells 2019, 8, 1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collie, G.; Reszka, A.P.; Haider, S.M.; Gabelica, V.; Parkinson, G.N.; Neidle, S. Selectivity in small molecule binding to human telomeric RNA and DNA quadruplexes. Chem. Commun. 2009, 48, 7482–7484. [Google Scholar] [CrossRef] [PubMed]
- Pirota, V.; Platella, C.; Musumeci, D.; Benassi, A.; Amato, J.; Pagano, B.; Colombo, G.; Freccero, M.; Doria, F.; Montesarchio, D. On the binding of naphthalene diimides to a human telomeric G-quadruplex multimer model. Int. J. Biol. Macromol. 2021, 166, 1320–1334. [Google Scholar] [CrossRef]
- Belmonte-Reche, E.; Benassi, A.; Peñalver, P.; Cucchiarini, A.; Guédin, A.; Mergny, J.L.; Rosu, F.; Gabelica, V.; Freccero, M.; Doria, F.; et al. Thiosugar naphthalene diimide conjugates: G-quadruplex ligands with antiparasitic and anticancer activity. Eur. J. Med. Chem. 2022, 15, 114183. [Google Scholar] [CrossRef]
- Prato, G.; Silvent, S.; Saka, S.; Lamberto, M.; Kosenkov, M. Thermodynamics of Binding of Di- and Tetrasubstituted Naphthalene Diimide Ligands to DNA G-Quadruplex. J. Phys. Chem. B 2015, 119, 3335–3347. [Google Scholar] [CrossRef]
- Marzano, S.; Pagano, B.; Iaccarino, N.; Porzio, A.D.; Tito, S.D.; Vertecchi, E.; Salvati, E.; Randazzo, A.; Amato, J. Targeting of Telomeric Repeat-Containing RNA G-Quadruplexes: From Screening to Biophysical and Biological Characterization of a New Hit Compound. Int. J. Mol. Sci. 2021, 22, 10315. [Google Scholar] [CrossRef]
- Silva, B.; Arora, R.; Bione, S.; Azzali, C.M. TERRA transcription destabilizes telomere integrity to initiate break-induced replication in human ALT cells. Nat. Commun. 2021, 12, 3760. [Google Scholar] [CrossRef]
- Martadinata, H.; Phan, A.T. Structure of Propeller-Type Parallel-Stranded RNA G-Quadruplexes, Formed by Human Telomeric RNA Sequences in K+ Solution. J. Am. Chem. Soc. 2009, 131, 2570–2578. [Google Scholar] [CrossRef]
- Shinohara, K.; Sannohe, Y.; Kaieda, S.; Tanaka, K.; Osuga, H.; Tahara, H.; Xu, Y.; Kawase, T.; Bando, T.; Sugiyama, H. A Chiral Wedge Molecule Inhibits Telomerase Activity. J. Am. Chem. Soc. 2010, 132, 3778–3782. [Google Scholar] [CrossRef]
- Phan, A.T.; Kuryavyi, V.; Burge, S.; Neidle, S.; Patel, D.J. Structure of an Unprecedented G-Quadruplex Scaffold in the Human c-kit Promoter. J. Am. Chem. Soc. 2007, 129, 4386–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formoso, C. The interaction of β-cyclodextrin with nucleic acid monomer units. Biochem. Biophys. Res. Commun. 1973, 50, 999–1005. [Google Scholar] [CrossRef]
- Tanious, F.A.; Yen, S.-F.; Wilson, W.D. Kinetic and equilibrium analysis of a threading intercalation mode: DNA sequence and ion effects. Biochemistry 1991, 30, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Nojima, T.; Waki, M.; Takenaka, S. Supramolecular Complex Formation by β-Cyclodextrin and Ferrocenylnaphthalene Diimide-intercalated Double Stranded DNA and Improved Electrochemical Gene Detection. Molecules 2005, 10, 693–707. [Google Scholar] [CrossRef] [Green Version]
- Matsue, T.; Evans, D.H.; Osa, T.; Kobayashi, N. Electron-transfer reactions associated with host-guest complexation. Oxidation of ferrocenecarboxylic acid in the presence of β-cyclodextrin. J. Am. Chem. Soc. 1985, 107, 3411–3417. [Google Scholar] [CrossRef]
- Benesi, H.A.; Hildbrand, J.H. A Spectrophotometric Investigation of the Interaction of Iodine with Aromatic Hydrocarbons. J. Am. Chem. Soc. 1949, 71, 2703–2707. [Google Scholar] [CrossRef]
- Chemical Computing Group ULC. Molecular Operating Environment (MOE); Chemical Computing Group: Montreal, QC, Canada, 2022. [Google Scholar]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.R.; Müller, K. MAB, a generally applicable molecular force field for structure modelling in medicinal chemistry. J. Comput. Aided Mol. Des. 1995, 9, 251–268. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | ||
|---|---|---|
| DNA | ds-oligo | 12-ss (+)5′-GGGAGGTTTCGC-3′ 12-ss (−)5′-GCGAAACCTCCC-3′ |
| Telomere G1 | 5′-TAG GGT TAG GGT TAG GGT TAG GG-3′ | |
| c-myc | 5′-TGA GGG TGG GGA GGG TGG GGA A-3′ | |
| c-kit | 5′-AGG GAG GGC GCT GGG AGG AGG G-3′ | |
| c-myc AP sites | 5′-TGA GGG XGG GGA GGG XGG GGA A-3′ | |
| RNA | G1-r23nt | 5′-UAG GGU UAG GGU UAG GGU UAG GG-3′ |
| G1-r12nt | 5′-UAG GGU UAG GGU-3′ |
| Telomere G4 | c-myc | ds-oligo | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| 10−5 Ka/M−1 (n) | 0.80 ± 0.01 b (-) | 0.89 ± 0.06 b (-) | 9.5 ± 0.9 a (1.6 ± 0.0) | 8.6 ± 0.5 a (1.2 ± 0.0) | 1.0 ± 0.2 b (-) | 10 ± 0.0 a (1.6 ± 0.0) | 0.68 ± 0.02 b (-) | 0.59 ± 0.08 b (-) | 3.9 ± 0.1 a (2.3 ± 0.0) |
| Hypochromicity/% | 32 | 26 | 54 | 44 | 36 | 51 | 25 | 17 | 54 |
| Telomere G4 | c-myc | c-myc AP Site | G1-r23nt | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| 10−5 Ka/M−1 | n.d. | n.d. | 9.5 ± 0.6 | 4.1 ± 0.5 | 3.2 ± 0.3 | 6.4 ± 0.5 | 2.2 ± 0.3 | 2.0 ± 0.8 | 2.9 ± 0.2 | 19.0 ± 2.2 | 12.3 ± 2.2 | n.d. |
| n | 2.1 ± 0.1 | 1.4 ± 0.0 | 1.7 ± 0.0 | 1.8 ± 0.1 | 0.92 ± 0.3 | 2.0 ± 0.2 | 2.4 ± 0.2 | 1.2 ± 0.2 | 1.2 ± 0.0 | |||
| ΔH/kcal mol−1 | −5.5 ± 0.1 | −13.8 ± 0.3 | −9.6 ± 0.5 | −6.9 ± 0.1 | −15.4 ± 0.2 | −11.1 ± 0.9 | −7.6 ± 0.7 | −14.0 ± 0.3 | −14.3 ± 0.0 | |||
| −TΔS/kcal mol−1 | −2.7 ± 0.2 | 6.2 ± 0.2 | 2.1 ± 0.5 | −1.0 ± 0.1 | 8.1 ± 0.3 | 3.9 ± 1.2 | −0.4 ± 0.2 | 5.4 ± 0.3 | 6.0 ± 0.1 | |||
| ΔG/kcal mol−1 | −8.2 ± 0.0 | −7.6 ± 0.1 | −7.5 ± 0.1 | −7.9 ± 0.0 | −7.3 ± 0.1 | −7.2 ± 0.3 | −7.5 ± 0.0 | −8.6 ± 0.1 | −8.3 ± 0.1 | |||
| Sequence | ΔTm (°C) | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| c-myc a | 4.8 ± 0.8 | 2.5 ± 0.5 | 14.0 ± 1.0 |
| Telomere G4 b | <0.5 | <0.5 | 5.1 ± 1.1 |
| G1-r23nt a | 10.3 ± 0.3 | 10.6 ± 0.6 | 19.9 ± 0.1 |
| G1-r12nt c | 2.1 ± 0.9 | <0.5 | 6.1 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, T.; Sato, Y.; Kaneyoshi, S.; Mano, K.; Yasukawa, R.; Nakano, Y.; Fujii, S.; Sato, S.; Takenaka, S. Naphthalene Diimides Carrying Two β-Cyclodextrins Prefer Telomere RNA G-Quadruplex Recognition. Molecules 2022, 27, 4053. https://doi.org/10.3390/molecules27134053
Zou T, Sato Y, Kaneyoshi S, Mano K, Yasukawa R, Nakano Y, Fujii S, Sato S, Takenaka S. Naphthalene Diimides Carrying Two β-Cyclodextrins Prefer Telomere RNA G-Quadruplex Recognition. Molecules. 2022; 27(13):4053. https://doi.org/10.3390/molecules27134053
Chicago/Turabian StyleZou, Tingting, Yuka Sato, Shuma Kaneyoshi, Kota Mano, Rui Yasukawa, Yoshifumi Nakano, Satoshi Fujii, Shinobu Sato, and Shigeori Takenaka. 2022. "Naphthalene Diimides Carrying Two β-Cyclodextrins Prefer Telomere RNA G-Quadruplex Recognition" Molecules 27, no. 13: 4053. https://doi.org/10.3390/molecules27134053