
Phytochemical Analysis, Antimutagenic and Antiviral Activity of Moringa oleifera L. Leaf Infusion: In Vitro and In Silico Studies


Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Preparation of M. oleifera Leaf Infusion
3.2. Total Phenolic Content
3.3. Total Flavonoid Content
3.4. Phytochemical Analysis Using Liquid Chromatography-Mass Spectrometry (LC-MS)
3.5. Antioxidant Activity with DPPH Radical Scavenging
3.6. DNA Protection Activity Assay
3.7. In-Silico Study of the Active Compound Infusion of M. oleifera Leaves with COVID-19 Main Protease
3.8. Data Analysis
4. Conclusions
5. Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mbikay, M. Therapeutic potential of Moringa oleifera leaves in chronic hyperglycemia and dyslipidemia: A review. Front. Pharmacol. 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food. Sci. Hum. Well. 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Jimenez, M.; Almatrafi, M.M.; Fernandez, M.L. Bioactive components in Moringa oleifera leaves protect against chronic disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, S.; Santacroce, A.; Longini, M.; Proietti, F.; Bazzini, F.; Bionocore, G. The free radical diseases of prematurity: From cellular mechanisms to bedside. Oxid. Med. Cell. Longev. 2018, 2018, 7483062. [Google Scholar] [CrossRef] [Green Version]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Izzotti, A. DNA damage and alterations of gene expression in chronic-degenerative diseases. Acta. Biochim. Pol. 2002, 50, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Fenech, M. Micronuclei and their association with sperm abnormalities, infertility, pregnancy loss, pre-eclampsia and intra-uterine growth restriction in humans. Mutagenesis 2010, 26, 63–67. [Google Scholar] [CrossRef]
- Sharma, A.D.; Kaur, I. Eucalyptol (1,8 cineole) from eucalyptus essential oil: A potential inhibitor of COVID-19 corona virus infection by molecular docking studies. Preprints 2020, 1, 1–8. [Google Scholar]
- Asif, M.; Saleem, M.; Saadullah, M.; Yaseen, H.S.; Al Zarzour, R. COVID-19 and therapy with essential oils having antiviral, anti-inflammatory, and immunomodulatory properties. Inflammopharmacology 2020, 28, 1153–1161. [Google Scholar] [CrossRef]
- Amparo, T.R.; Seibert, J.B.; Silveira, B.M.; Costa, F.S.F.; Almeida, T.C.; Braga, S.F.P.; da Silva, G.N.; dos Santos, O.D.H.; Bianco de Souza, G.H. Brazilian essential oils as source for the discovery of new anti-COVID-19 drug: A review guided by in silico study. Phytochem. Rev. 2021, 20, 1013–1032. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogra. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Peng, B.; Bai, R.F.; Li, P.; Han, X.Y.; Wang, H.; Zhu, C.C.; Zeng, Z.P.; Chai, X.Y. Two new glycosides from Dryopteris fragrans with anti-inflammatory activities. J. Asian. Nat. Prod. Res. 2016, 18, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Atawodi, S.E.; Atawodi, J.C.; Idakwo, G.A.; Pfundstein, B.; Haubner, R.; Wurtele, G.; Bartsch, H.; Owen, R.W. Evaluation of the polyphenol content and antioxidant properties of methanol extracts of the leaves, stem, and root barks of Moringa oleifera Lam. J. Med. Food. 2010, 13, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, V.C.; Krishna, B.G.; Viswanatha, G.L. Simultaneous determination of quercetin, rutin and kaempferol in the leaf extracts of Moringa oleifera Lam. and Raphinus sativus Linn. by liquid chromatography-tandem mass spectrometry. Zhong Xi Yi Jie He Xue Bao 2019, 9, 1022–1030. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, A.; Foidl, N.; Pratt, J.H.; Dupont, M.S.; Perkins, L.; Kroon, P.A. Profiling glucosinolates and phenolics in vegetative and reproductive tissues of the multi-purpose trees Moringa oleifera L. (Horseradish tree) and Moringa stenopetala L. J. Agric. Food. Chem. 2003, 51, 3546–3553. [Google Scholar] [CrossRef]
- Maiyo, F.C.; Moodley, R.; Singh, M. Cytotoxicity, antioxidant and apoptosis studies of quercetin-3-O glucoside and 4-(β-D-glucopyranosyl-1→4-α-L-rhamnopyranosyloxy)-benzyl isothiocyanate from Moringa oleifera. Anti-Cancer Agents Med. Chem. 2016, 16, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant flavone apigenin: An emerging anticancer agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anticancer effects and mechanisms of action. Cell. Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Wei-Jian, Y.; Xiao-Jun, H.; Chun-Lin, F.; Lin, X.; Ying, W.; Wen-Cai, Y. Two new flavonoid glycosides from leaves of Moringa oleifera. Zhongguo Zhong Yao Za Zhi 2018, 43, 3150–3156. [Google Scholar]
- Razavi, S.M.; Zahri, S.; Zarrini, G.; Nazemiyeh, H.; Mohammadi, S. Biological activity of quercetin-3-O-glucoside, a known plant flavonoid. Russ. J. Bioorg. Chem. 2009, 35, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.X.; Silva, S.F.; Guedes, R.J.; Almeida, S. Biological Oxidations and Antioxidant Activity of Natural Products. Phytochemicals as Nutraceuticals-Global Approaches to Their Role in Nutrition and Health; Intech Open: London, UK, 2012; pp. 1–22. [Google Scholar]
- Sharma, H.; Kanwal, R.; Bhaskaran, N.; Gupta, S. Plant flavone apigenin binds to nucleic acid bases and reduces oxidative DNA damage in prostate epithelial cells. PLoS ONE 2014, 9, e91588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.F.; Chao, H.; Zhou, Y.F.; Ji, L.N. Synthesis, characterization, DNA-binding and DNA-photocleavage studies of [Ru(bpy)2(BPIP)]2+ and [Ru(phen)2(BPIP)]2+ (BPIP=2-(4′-biphenyl)imidazo[4,5-f][1,10]phenanthroline). Polyhedron 2007, 26, 3029–3036. [Google Scholar] [CrossRef]
- Niering, P.; Michels, G.; Wätjen, W.; Ohler, S.; Steffan, B.; Chovolou, Y.; Kampkötter, A.; Proksch, P.; Kahl, R. Protective and detrimental effects of kaempferol in rat H4IIE cells: Implication of oxidative stress and apoptosis. Toxicol. Appl. Pharmacol. 2005, 209, 114–122. [Google Scholar] [CrossRef]
- Devisetti, R.; Sreerama, Y.N.; Bhattacharya, S. Processing effects on bioactive components and functional properties of moringa leaves: Development of a snack and quality evaluation. J. Food. Sci. Technol. 2016, 53, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, N.; Mohamed, S.; Rahman, H.S.; Rosli, R. Epicatechin and scopoletin-rich Morinda citrifolia leaf ameliorated leukemia via anti-inflammatory, anti-angiogenesis, and apoptosis pathways in vitro and in vivo. J. Food. Biochem. 2019, 43, e12868. [Google Scholar] [CrossRef]
- Hakki, Z.; Cao, B.; Heskes, A.M.; Goodger, J.Q.; Woodrow, I.E.; Williams, S.J. Synthesis of the monoterpenoid esters cypellocarpin C and cuniloside B and evidence for their widespread occurrence in eucalyptus. Carbohydr. Res. 2010, 345, 2079–2084. [Google Scholar] [CrossRef]
- Kim, S.B.; Ahn, J.H.; Han, S.B.; Hwang, B.Y.; Kim, S.Y.; Lee, M.K. Anti-adipogenic chromone glycosides from Conidium monnieri fruits in 3T3-L1 cells. Bioorg. Med. Chem. Lett. 2012, 22, 6267–6271. [Google Scholar] [CrossRef]
- Koz, O.; Pizza, C.; Kırmızıgül, S. Triterpene and flavone glycosides from Anchusa undulata subsp. Hybrida. Nat. Prod. Res. 2009, 23, 284–292. [Google Scholar] [CrossRef]
- Pereira, L.C.O.; Abreu, L.S.; Silva, J.P.R.E.; Machado, F.S.V.L.; Queiroga, C.S.; do Esp Rito-Santo, R.F.; de Agnelo-Silva, D.F.; Villareal, C.F.; Agra, M.d.F.; Scotti, M.T.; et al. Bioactive compounds from the aerial parts of Evolvulus linarioides. J. Nat. Prod. 2020, 83, 1515–1523. [Google Scholar] [CrossRef]
- Wang, Y.B.; Huang, R.; Zhang, H.B.; Li, L. Chromone glycosides from Knoxia corymbosa. J. Asian. Nat. Prod. Res 2006, 8, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Song, X.; Wang, D.; Wang, X.; Wang, X. Five new chromone glycosides from Scindapsus officinalis (Roxb.) Schott. Fitoterapia 2017, 22, 101–106. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Chanathong, B. Alpha-glucosidase inhibitory activity and lipid-lowering mechanisms of Moringa oleifera leaf extract. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 803–808. [Google Scholar] [PubMed]
- Ioannis, S.P.; Siomou, S.; Bozinou, E.; Lalas, S.I. Moringa oleifera leaves crude aqueous extract down-regulates of BRCA1, mta-1 and oncogenes c-myc and p53 in AsPC-1, MCF-7 and HTC-116 cells. Food Biosci. 2021, 43, 101221. [Google Scholar]
- Adebayo, I.A.; Arsad, H.; Samian, M.R. Total phenolics, total flavonoids, antioxidant capacities, and volatile compounds gas chromatography-mass spectrometry profiling of Moringa oleifera ripe seed polar fractions. Pharmacogn. Mag. 2018, 14, 191–194. [Google Scholar]
- George, V.C.; Kumar, D.R.N.; Suresh, P.K.; Kumar, R.A. Antioxidant, DNA protective efficacy and HPLC analysis of Annona muricata (soursop) extracts. J. Food. Sci. Technol. 2015, 52, 2328–2335. [Google Scholar] [CrossRef]
- Umamaheswari, M.; Chatterjee, T.K. In vitro antioxidant activities of the fractions of Coccinnia grandis L. leaf extract. Afr. J. Trad. Compl. Altern. Med. 2008, 5, 61–73. [Google Scholar]
- Chatsumpun, M.; Chuanasa, T.; Sritularak, B.; Likhitwitayawuid, K. Oxyresveratrol protects against DNA damage induced by photosensitized riboflavin. Nat. Prod. Commun. 2011, 6, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Salim, R.; Eliyarti, E. Aktivitas antioksidan infusa daun Kelor (Moringa Oleifera Lam.) terhadap warna daun. J. Katalisator 2019, 4, 91–102. [Google Scholar] [CrossRef]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of natural products as potential inhibitors of COVID-19 main protease (Mpro): In-silico evidences. Nat. Prod. Bioprospect. 2020, 10, 297–306. [Google Scholar] [CrossRef]
- Muhammad, S.; Hassan, S.H.; Shakir, H.A.; Khan, M.; Irfan, M. Exploring the New Potential Antiviral Constituents of Moringa oliefera for SARS-COV-2 Pathogenesis: An in Silico Molecular Docking and Dynamic Studies. Chem. Phys. Lett. 2021, 767, 138379. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.A.; James, T.J. Computational screening of phytocompounds from Moringa oleifera leaf as potential inhibitors of SARS-CoV-2 Mpro. Preprint 2020, 1–14. [Google Scholar]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the treatment of COVID-19- Final report. N. Eng. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef] [PubMed]
- Tahir ul Qamar, M.; Alqahtani, S.M.; Alamri, M.A.; Ling-ling, C. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef]
- Ferreira, J.C.; Fadl, S.; Villanueva, A.J.; Rabeh, W.M. Catalytic dyad residues His41 and Cys145 impact the catalytic activity and overall conformational fold of the main SARS-CoV-2 protease 3-Chymotrypsin-like protease. Front. Chem. 2021, 9, 491. [Google Scholar] [CrossRef]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali; Kepel, B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of plant bioactive compounds as SARS-CoV-2 main protease (Mpro) and spike (S) glycoprotein inhibitors: A molecular docking study. Scientifica 2020, 2020, 6307457. [Google Scholar] [CrossRef]
- Umar, H.I.; Josiah, S.S.; Saliu, T.P.; Jimoh, T.O.; Ajayi, A.; Danjuma, J.B. In-silico analysis of the inhibition of the SARS-CoV-2 main protease by some active compounds from selected African plants. J. Taibah. Univ. Med. Sci. 2021, 16, 162–176. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Park, J.; Kim, D.H.; Hong, S.Y.; Lee, J.S.; Kim, M. Total phenolic content, antioxidant, tyrosinase and α-glucosidase inhibitory activities of water soluble infusions of noble starter culture Doenjang, a Korean fermented soybean sauce variety. Food. Control. 2016, 59, 854–861. [Google Scholar] [CrossRef]
- Chia-Chi, C.; Ming Hua, Y.; Hwei-Mei, W.; Jiing-Chuan, C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food. Drug. Anal. 2002, 10, 178–182. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Jeong, J.B.; Park, J.H.; Lee, H.K.; Ju, S.Y.; Hong, S.C.; Lee, J.R.; Chung, G.Y.; Lim, J.H.; Jeong, H.J. Protective effect of the extracts from Conidium officinale against oxidative damage induced by hydrogen peroxide via antioxidant effect. Food. Chem. Toxicol. 2009, 47, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Rahayu, I.; Timotius, K.H. Antimutagenic and antiviral activity of hydroethanolic extract from Moringa oleifera L. leaves: In vitro and in silico studies. Int. J. Herb. Med. 2021, 9, 25–32. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
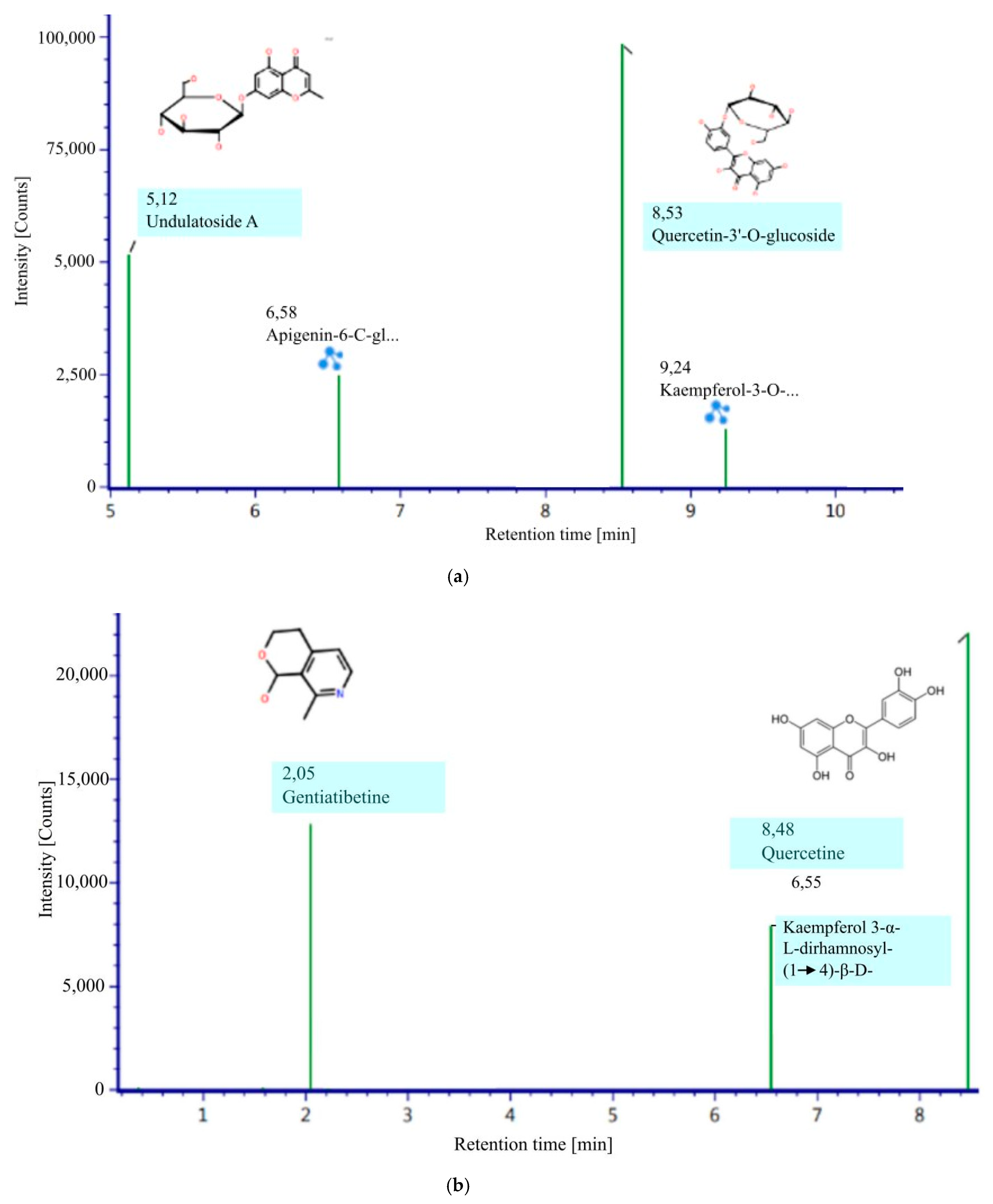
| No | Identified Compounds | Ionization Mode | RT | MZ | Molecular Formula | Response |
|---|---|---|---|---|---|---|
| Alkaloid | ||||||
| 1 | Gentiatibetine | positive | 3.37 | 77, 103, 120 | C9H11NO2 | 12.820 |
| Flavonoid | ||||||
| 2 | Quercetin | positive | 8.48 | 303, 304, 487 | C15H10O7 | 22.083 |
| 3 | Kaempferol 3-α-L-dirhamnosyl-(1 → 4)-β-D-glucopyranoside | positive | 6.55 | 457, 495, 633 | C27H30O15 | 7.934 |
| 4 | Apigenin-6-C- glucosylglucoside | negative | 6.58 | 353, 593, 646 | C27H30O15 | 24.929 |
| 5 | Quercetin-3′-O-glucoside | negative | 8.53 | 271, 300, 463 | C21H20O12 | 98.283 |
| 6 | Undulatoside A | negative | 5.12 | 173, 191, 353 | C16H18O9 | 51.556 |
| 7 | Kaempferol-3-Oneohesperidoside | negative | 9.24 | 301, 593, 607 | C27H30O15 | 12.952 |
| Code | Treatment | Nick (%) | Linear (%) | SC (%) |
|---|---|---|---|---|
| A | Plasmid + H2O2 + Fe2SO4 | 80.6 | 19.4 | |
| B | 1.25 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 79.4 | 20.6 | |
| C | 2.5 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 76.8 | 23.2 | |
| D | 5 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 76.4 | 23.5 | |
| E | 10 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 79.6 | 20.4 | |
| F | 20 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 35.7 | 47.5 | 16.8 |
| G | 40 mg/mL infusion + Plasmid + H2O2 + Fe2SO4 | 29.5 | 49.6 | 20.8 |
| H | Non treated Plasmid | 17.9 | 60.6 | 21.4 |
| No | Active Compound | Mpro (6lu7) (kcal/mol) | rmsd/ub | rmsd/lb |
|---|---|---|---|---|
| 1 | Gentiatibetine | −5.1 | 0.00 | 0.00 |
| 2 | Quercetin | −7.5 | 0.00 | 0.00 |
| 3 | Quercetin-3′-O-glucoside | −8.2 | 0.00 | 0.00 |
| 4 | Undulatoside A | −7.7 | 0.00 | 0.00 |
| 5 | Kaempferol-3-O-neohesperidoside | −5.7 | 0.00 | 0.00 |
| 6 | Apigenin | −7.8 | 0.00 | 0.00 |
| 7 | Kaempferol | −7.8 | 0.00 | 0.00 |
| 8 | Remdesivir | −7.3 | 0.00 | 0.00 |
| No | Active Compound | Interaction | Amino Acid Residues |
|---|---|---|---|
| 1 | Gentiatibetine | pi-donor hydrogen | His41 |
| pi-alkyl | Met165 | ||
| 2 | Quercetin | pi-sulfur | Cys145 |
| pi-alkyl | Met49 | ||
| pi-donor hydrogen | Glu166 | ||
| van der Waals | His163, Phe140, Ser144, Leu141, His164, His41, Asp187, Tyr54, Arg188, and Gln189 | ||
| 3 | Quercetin-3′-O-glucoside | conventional hydrogen | Phe140 and Glu166 |
| van der Waals | His172, Leu141, His163, Ser144, His164, Asn142, Gly143, Arg188, Gln189, Asp187, Tyr54, His41 | ||
| pi-sulphur | Cys145 | ||
| pi-alkyl | Met49 and Met165 | ||
| 4 | Undulatoside A | pi-sigma | His41 |
| conventional hydrogen | His163, Ser144, Cys145 | ||
| carbon hydrogen | Gln189, Asn142 | ||
| van der Waals | Arg188, His164, Glu166, His172, Phe140, Leu141 and Gly143 | ||
| 5 | Kaempferol-3-O-neohesperidoside | conventional hydrogen | Glu166 and Gly143 |
| pi-anion | Gly143 | ||
| unfavorable donor-donor | Gly143 | ||
| carbon-hydrogen | Arg188 | ||
| van der Waals | Phe140, Thr190, Leu167, Gln192, Met165, Leu141, Gln149, Asn142, Ser144, His164, His41, Met49, Cys145 | ||
| pi-alkyl | Met49 | ||
| 6 | Apigenin | pi-sulphur | Cys145 |
| hydrogen-donor pi | Glu166 | ||
| van der Waals force | Gln189, Arg188, Asp187, Tyr54, Pro52, His41, His164, Met165, Leu141, Phe140, His163, and Ser144 | ||
| 7 | Kaempferol | conventional hydrogen bond | Gln189 and Asp187 |
| pi-donor hydrogen | Glu166 | ||
| pi-sulphur | Cys145, Met165 | ||
| pi-alkyl | Met49 | ||
| pi-pi stacked | His-41 | ||
| Unfavorable acceptor-acceptor | Leu141 | ||
| van der Waals | Asn142, Phe140, Ser144, His163, His164, Arg188, Tyr54 | ||
| 8 | Remdesivir | pi-alkyl | Met165, Pro168, Leu167 |
| carbon-hydrogen | Gln189 | ||
| pi-cation | His41 | ||
| hydrogen | His164 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahayu, I.; Timotius, K.H. Phytochemical Analysis, Antimutagenic and Antiviral Activity of Moringa oleifera L. Leaf Infusion: In Vitro and In Silico Studies. Molecules 2022, 27, 4017. https://doi.org/10.3390/molecules27134017
Rahayu I, Timotius KH. Phytochemical Analysis, Antimutagenic and Antiviral Activity of Moringa oleifera L. Leaf Infusion: In Vitro and In Silico Studies. Molecules. 2022; 27(13):4017. https://doi.org/10.3390/molecules27134017
Chicago/Turabian StyleRahayu, Ika, and Kris Herawan Timotius. 2022. "Phytochemical Analysis, Antimutagenic and Antiviral Activity of Moringa oleifera L. Leaf Infusion: In Vitro and In Silico Studies" Molecules 27, no. 13: 4017. https://doi.org/10.3390/molecules27134017