
Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264.7 Cells

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
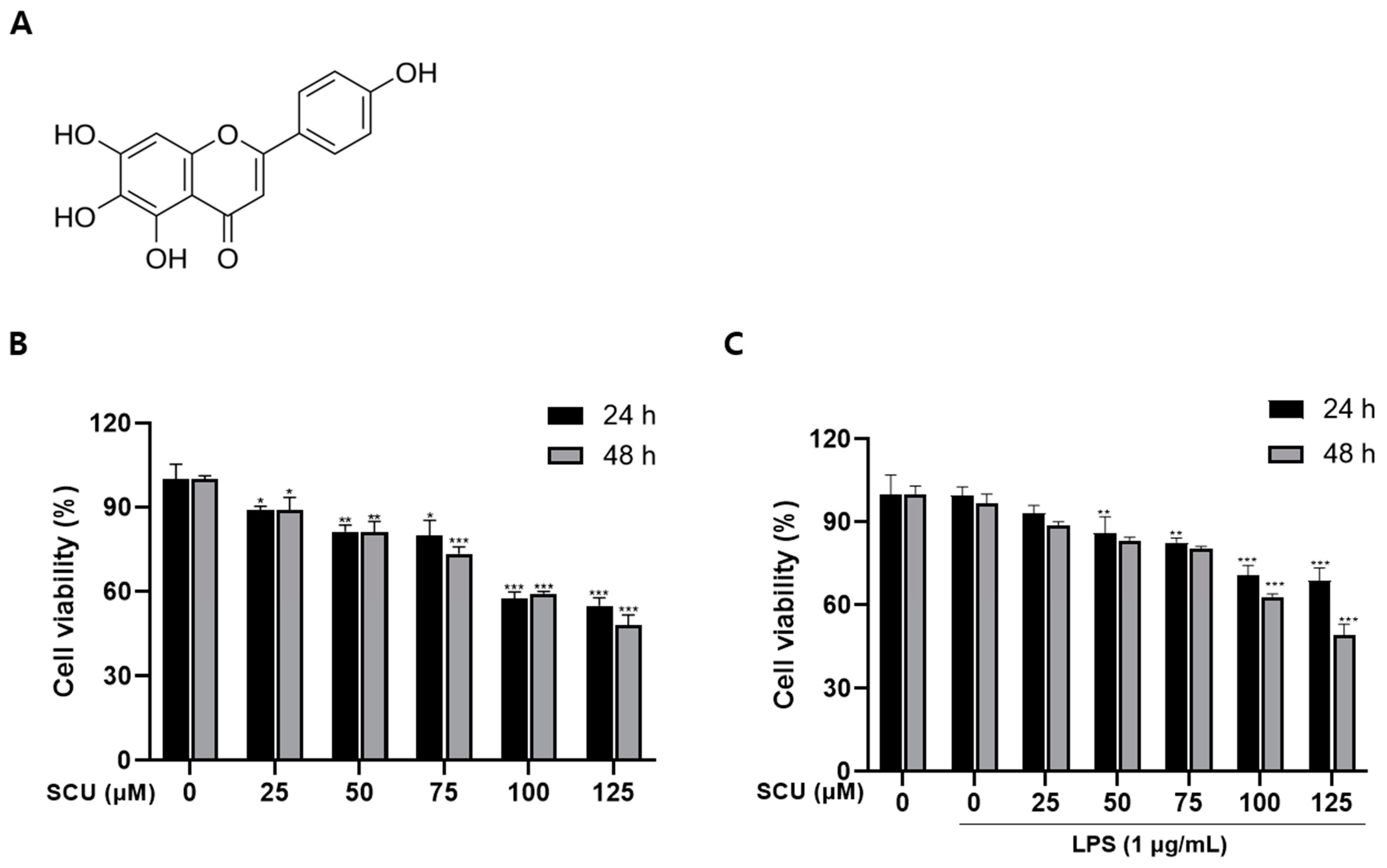
2.1. The Effect of SCU on the Cell Viability of RAW264.7 Cells
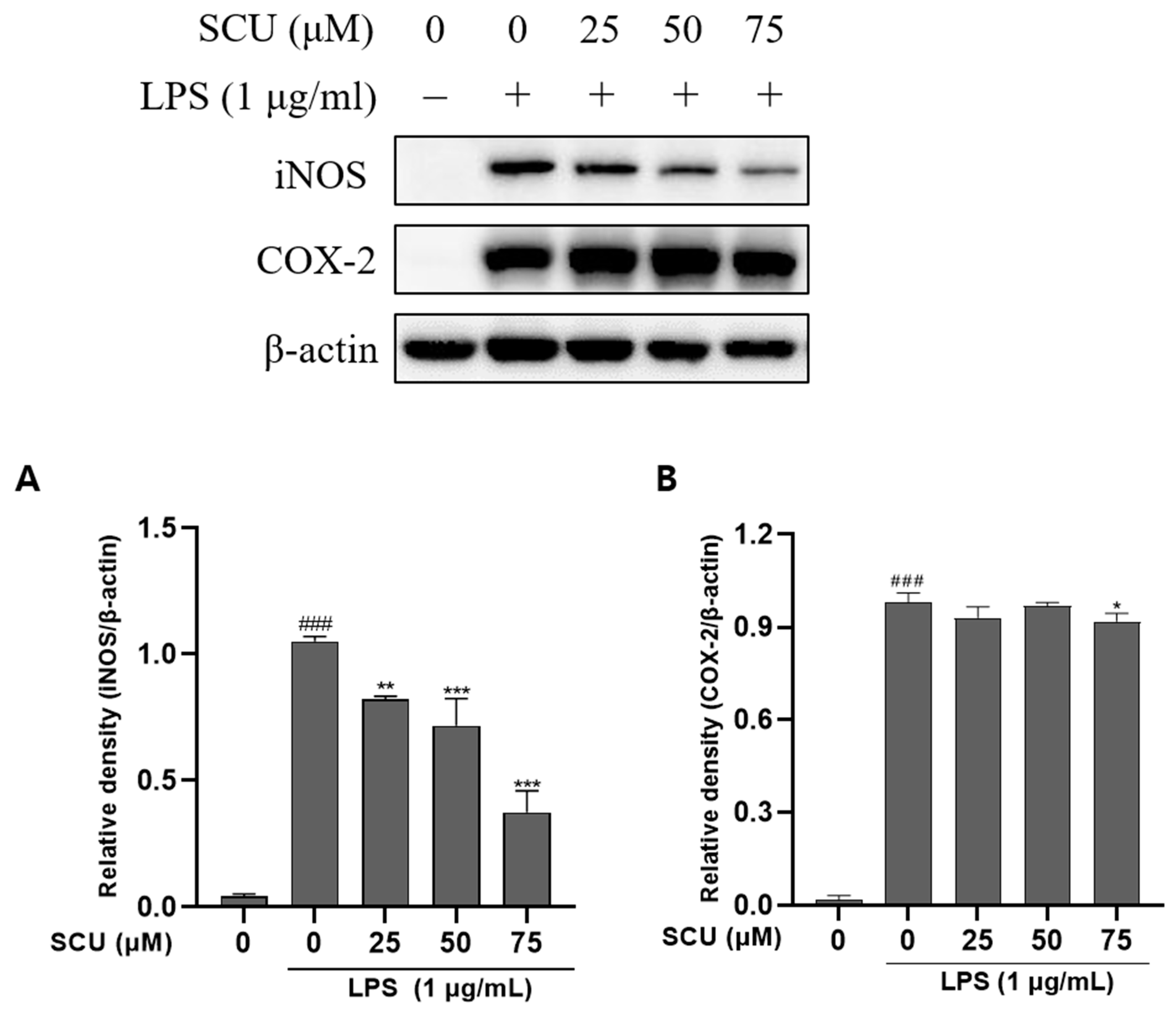
2.2. Effects of SCU on iNOS and COX-2 Expression of LPS-Induced RAW264.7 Cells
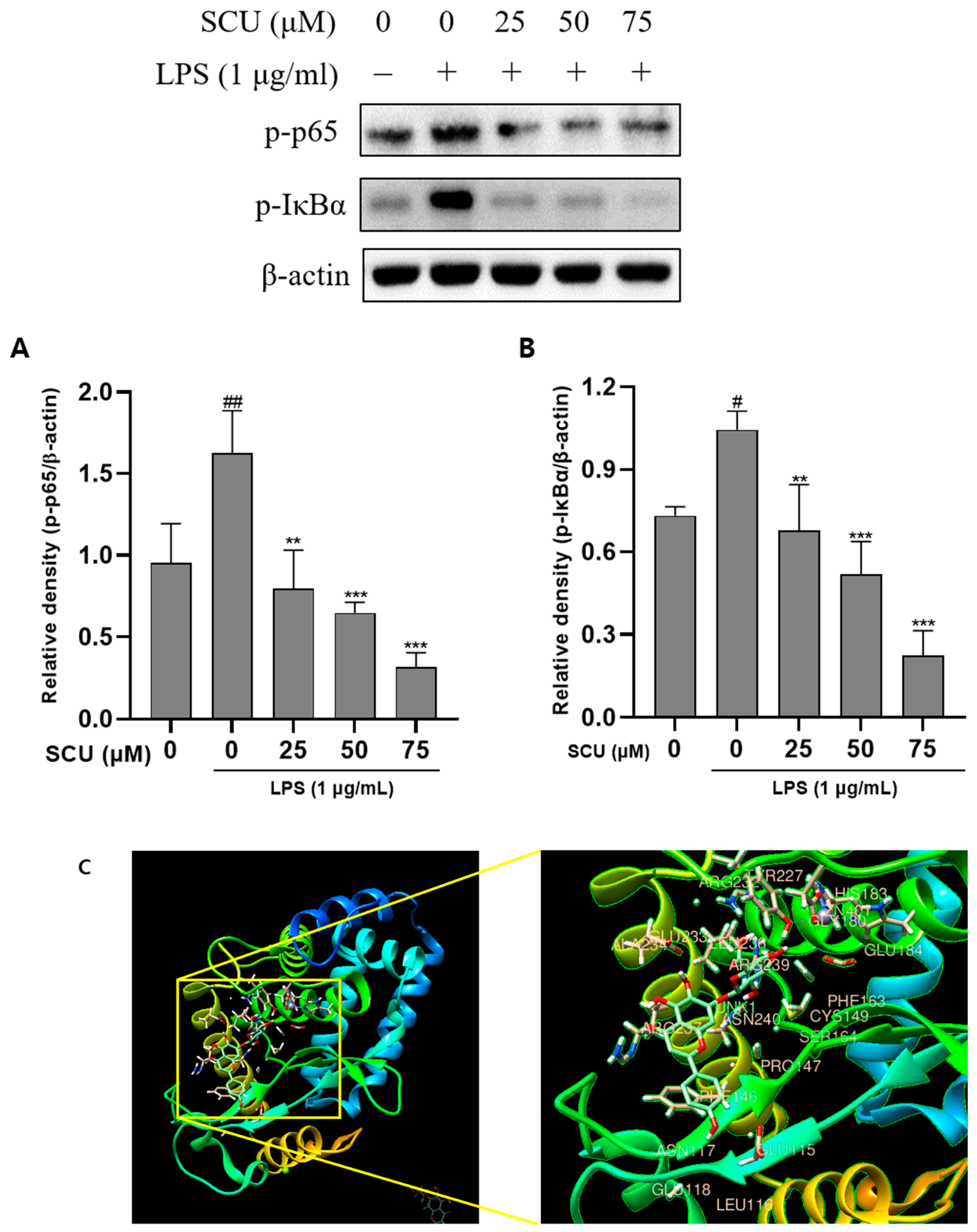
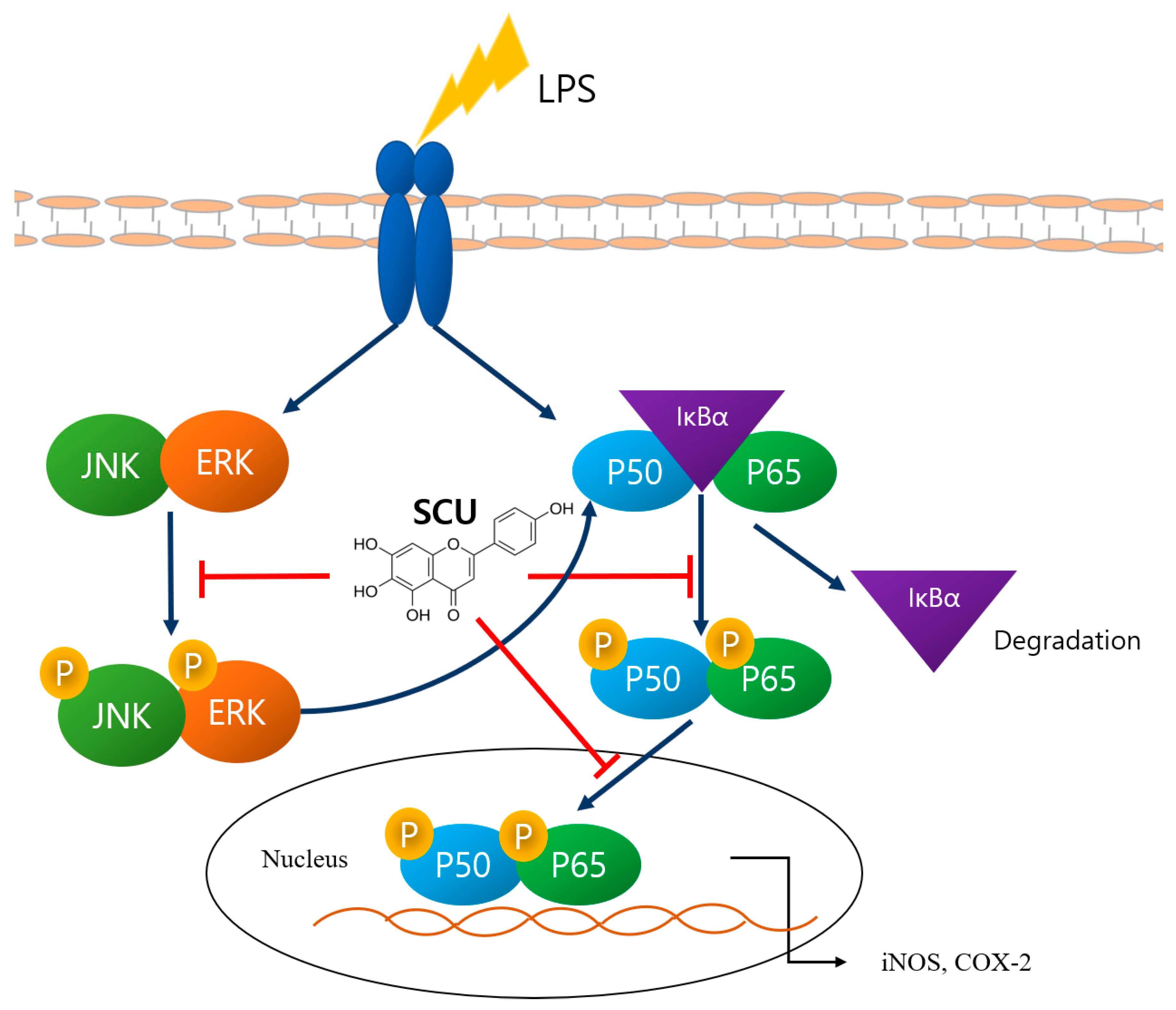
2.3. Inhibition of LPS-Induced NF-кB Pathways Activation by SCU
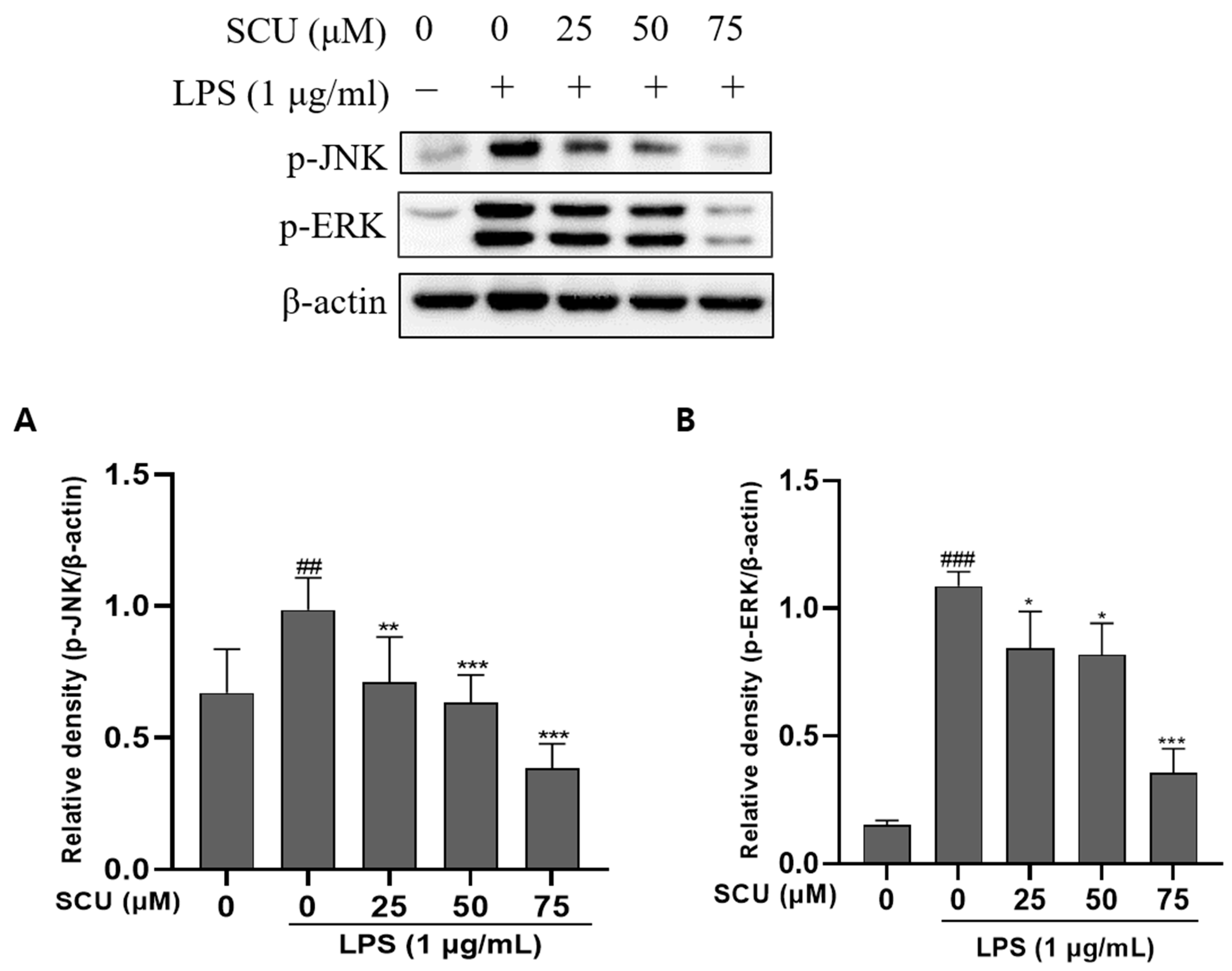
2.4. Inhibition of LPS-Induced MAPKs Pathways Activation by SCU
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Scutellarein (SCU) Treatment
4.3. Cell Viability Assay
4.4. Western Blot
4.5. Molecular Docking
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| TNF-α | Tumor necrosis factor |
| LPS | Lipopolysaccharide |
| COX-2 | Cytooxygenase |
| iNOS | Inducible nitric oxide synthase |
| NF-κB | Nuclear factor-kappa B |
| MAPK | Mitogen activated protein |
| ERK | Extracellular signal-regulated kinase |
| JNK | c-Jun NH2-terminal kinase |
| SCU | Scutellarein |
| NO | Nitric oxide |
| PG | Prostaglandin |
| PGE2 | Prostaglandin E2 |
| AP | Activator protein |
| IKK | IкB-kinase |
| SAPKs | Stress-activated protein kinase |
References
- Lee, G.S.; Kang, T.J. Cuttlebone Shows Anti-inflammatory Activity Via Suppression of NF-κB Activation in LPS-induced RAW 264.7 Macrophages. Yakhak Hoeji 2021, 65, 41–45. [Google Scholar] [CrossRef]
- Alam, M.B.; Chowdhury, N.S.; Sohrab, M.H.; Rana, M.S.; Hasan, C.M.; Lee, S.-H. Cerevisterol Alleviates Inflammation via Suppression of MAPK/NF-κB/AP-1 and Activation of the Nrf2/HO-1 Signaling Cascade. Biomolecules 2020, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Vetrivel, P.; Kim, H.H.; Ha, S.E.; Venkatarame Gowda Saralamma, V.; Kim, G.S. Artemisia iwayomogi (Dowijigi) inhibits lipopolysaccharide-induced inflammation in RAW264.7 macrophages by suppressing the NF-κB signaling pathway. Exp. Ther. Med. 2020, 19, 2161–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.M.; Song, Y.-S. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-κB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. NRP 2013, 7, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Li, X.; Wu, H.; Long, W.; Jiang, X.; Shen, T.; Qiang, Q.; Si, C.; Wang, X.; Jiang, Y.; et al. 5-Methoxyl Aesculetin Abrogates Lipopolysaccharide-Induced Inflammation by Suppressing MAPK and AP-1 Pathways in RAW 264.7 Cells. Int. J. Mol. Sci. 2016, 17, 315. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Biol. Interact 2010, 188, 659–667. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Mulero, M.C.; Huxford, T.; Ghosh, G. NF-κB, IκB, and IKK: Integral Components of Immune System Signaling. In Structural Immunology; Jin, T., Yin, Q., Eds.; Springer: Singapore, 2019; pp. 207–226. [Google Scholar] [CrossRef]
- Han, Y.; Li, X.; Zhang, X.; Gao, Y.; Qi, R.; Cai, R.; Qi, Y. Isodeoxyelephantopin, a sesquiterpene lactone from Elephantopus scaber Linn., inhibits pro-inflammatory mediators’ production through both NF-κB and AP-1 pathways in LPS-activated macrophages. Int. Immunopharmacol. 2020, 84, 106528. [Google Scholar] [CrossRef]
- Xie, C.; Li, X.; Zhu, J.; Wu, J.; Geng, S.; Zhong, C. Magnesium isoglycyrrhizinate suppresses LPS-induced inflammation and oxidative stress through inhibiting NF-κB and MAPK pathways in RAW264.7 cells. Bioorganic Med. Chem. 2019, 27, 516–524. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer activity of the plant flavonoid luteolin against preclinical models of various cancers and insights on different signalling mechanisms modulated. Phytother. Res. 2021, 35, 3509–3532. [Google Scholar] [CrossRef]
- Khan, A.; Ikram, M.; Hahm, J.R.; Kim, M.O. Antioxidant and Anti-Inflammatory Effects of Citrus Flavonoid Hesperetin: Special Focus on Neurological Disorders. Antioxidants 2020, 9, 609. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Han, W.; Fan, T.; Wang, X.; Cheng, Z.; Wan, B.; Chen, J. Scutellarin inhibits human renal cancer cell proliferation and migration via upregulation of PTEN. Biomed. Pharmacother. 2018, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Sang Eun, H.; Seong Min, K.; Ho Jeong, L.; Vetrivel, P.; Venkatarame Gowda Saralamma, V.; Jeong Doo, H.; Eun Hee, K.; Sang Joon, L.; Gon Sup, K. Scutellarein Induces Fas-Mediated Extrinsic Apoptosis and G2/M Cell Cycle Arrest in Hep3B Hepatocellular Carcinoma Cells. Nutrients 2019, 11, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Zhang, C.; Hu, M.; Su, K. Scutellarein relieves the death and inflammation of tubular epithelial cells in ischemic kidney injury by degradation of COX-2 protein. Int. Immunopharmacol. 2021, 101, 108193. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Shi, X.; Zhao, L.; Lv, T.; Yuan, Q.; Hao, W.; Zhu, J. Anti-proliferative and anti-migratory effects of Scutellaria strigillosa Hemsley extracts against vascular smooth muscle cells. J. Ethnopharmacol. 2019, 235, 155–163. [Google Scholar] [CrossRef]
- Lang, X.; Chen, Z.; Yang, X.; Yan, Q.; Xu, M.; Liu, W.; He, Q.; Zhang, Y.; Cheng, W.; Zhao, W. Scutellarein induces apoptosis and inhibits proliferation, migration, and invasion in ovarian cancer via inhibition of EZH2/FOXO1 signaling. J. Biochem. Mol. Toxicol. 2021, 35, e22870. [Google Scholar] [CrossRef]
- Li, S.T.; Dai, Q.; Zhang, S.X.; Liu, Y.J.; Yu, Q.Q.; Tan, F.; Lu, S.H.; Wang, Q.; Chen, J.W.; Huang, H.Q.; et al. Ulinastatin attenuates LPS-induced inflammation in mouse macrophage RAW264.7 cells by inhibiting the JNK/NF-kappaB signaling pathway and activating the PI3K/Akt/Nrf2 pathway. Acta Pharmacol. Sin. 2018, 39, 1294–1304. [Google Scholar] [CrossRef]
- Huang, G.J.; Huang, S.S.; Deng, J.S. Anti-inflammatory activities of inotilone from Phellinus linteus through the inhibition of MMP-9, NF-kappaB, and MAPK activation in vitro and in vivo. PLoS ONE 2012, 7, e35922. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhou, Y.; Chen, S.; Hu, Y.; Zhu, Z.; Wang, Y.; Du, N.; Song, T.; Yang, Y.; Guo, A.; et al. Macrophage migration inhibitory factor facilitates prostaglandin E2 production of astrocytes to tune inflammatory milieu following spinal cord injury. J. Neuroinflammation 2019, 16, 85. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin inhibits TNF-α induced HUVECs apoptosis and inflammation via downregulating NF-kB and AP-1 signaling pathway in vitro. Medicine 2020, 99, e22241. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Feng, L.; Zhang, Z.H.; Jia, X.B. The anti-inflammation effect of baicalin on experimental colitis through inhibiting TLR4/NF-κB pathway activation. Int. Immunopharmacol. 2014, 23, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.-Y.; Ju, J.-M.; Mo, L.-H.; Ma, L.; Hu, W.-H.; You, R.-R.; Chen, X.-Q.; Chen, Y.-Y.; Liu, Z.-Q.; Qiu, S.-Q.; et al. Anti-inflammation action of xanthones from Swertia chirayita by regulating COX-2/NF-κB/MAPKs/Akt signaling pathways in RAW 264.7 macrophage cells. Phytomedicine 2019, 55, 214–221. [Google Scholar] [CrossRef]
- Shi, X.; Chen, G.; Liu, X.; Qiu, Y.; Yang, S.; Zhang, Y.; Fang, X.; Zhang, C.; Liu, X. Scutellarein inhibits cancer cell metastasis in vitro and attenuates the development of fibrosarcoma in vivo. Int. J. Mol. Med. 2015, 35, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.Y.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Abusaliya, A.; Jeong, S.H.; Park, J.-S.; Heo, J.D.; Kim, G.S. Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264.7 Cells. Molecules 2022, 27, 3782. https://doi.org/10.3390/molecules27123782
Park MY, Ha SE, Kim HH, Bhosale PB, Abusaliya A, Jeong SH, Park J-S, Heo JD, Kim GS. Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264.7 Cells. Molecules. 2022; 27(12):3782. https://doi.org/10.3390/molecules27123782
Chicago/Turabian StylePark, Min Yeong, Sang Eun Ha, Hun Hwan Kim, Pritam Bhagwan Bhosale, Abuyaseer Abusaliya, Se Hyo Jeong, Joon-Suk Park, Jeong Doo Heo, and Gon Sup Kim. 2022. "Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264.7 Cells" Molecules 27, no. 12: 3782. https://doi.org/10.3390/molecules27123782