
In Search of Synergistic Insect Repellents: Modeling of Muscarinic GPCR Interactions with Classical and Bitopic Photoactive Ligands



Abstract
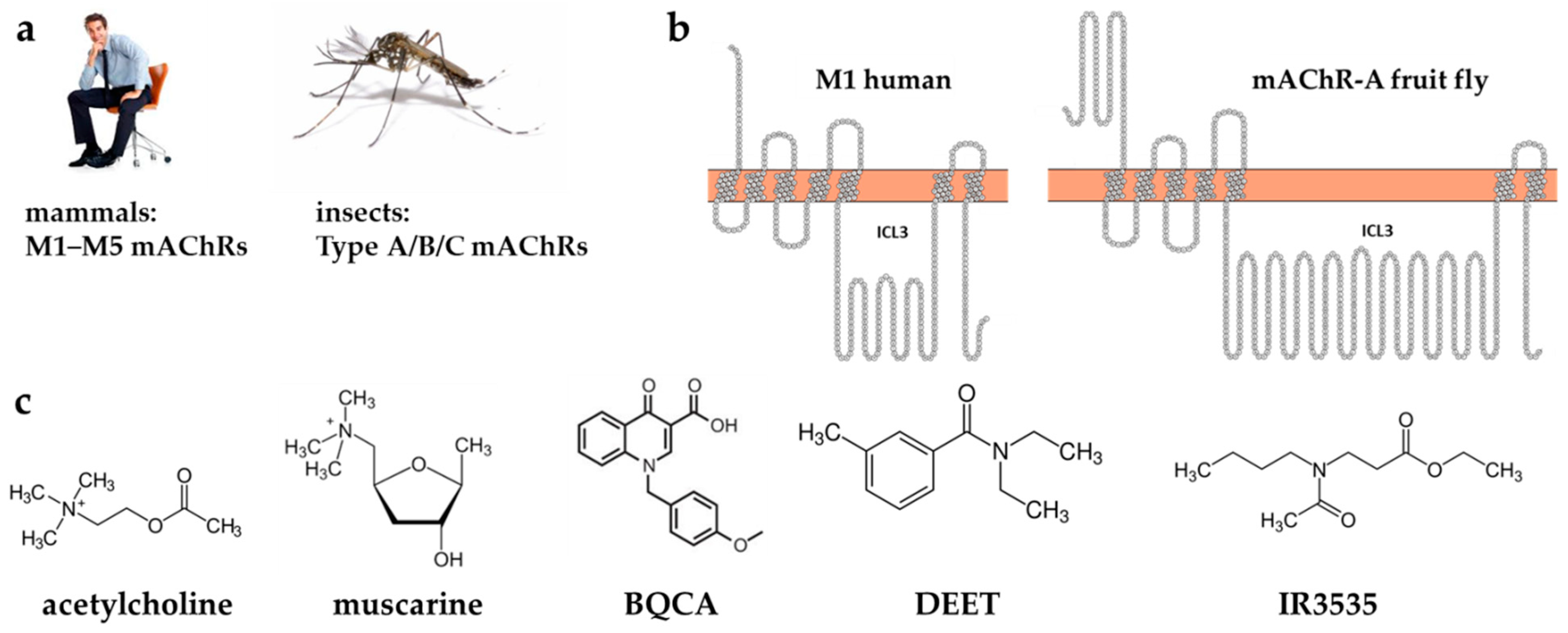
:1. Introduction
2. Results and Discussion
2.1. Single Ligands Docking
2.1.1. Single Ligands Docking to Human M1 mAChR
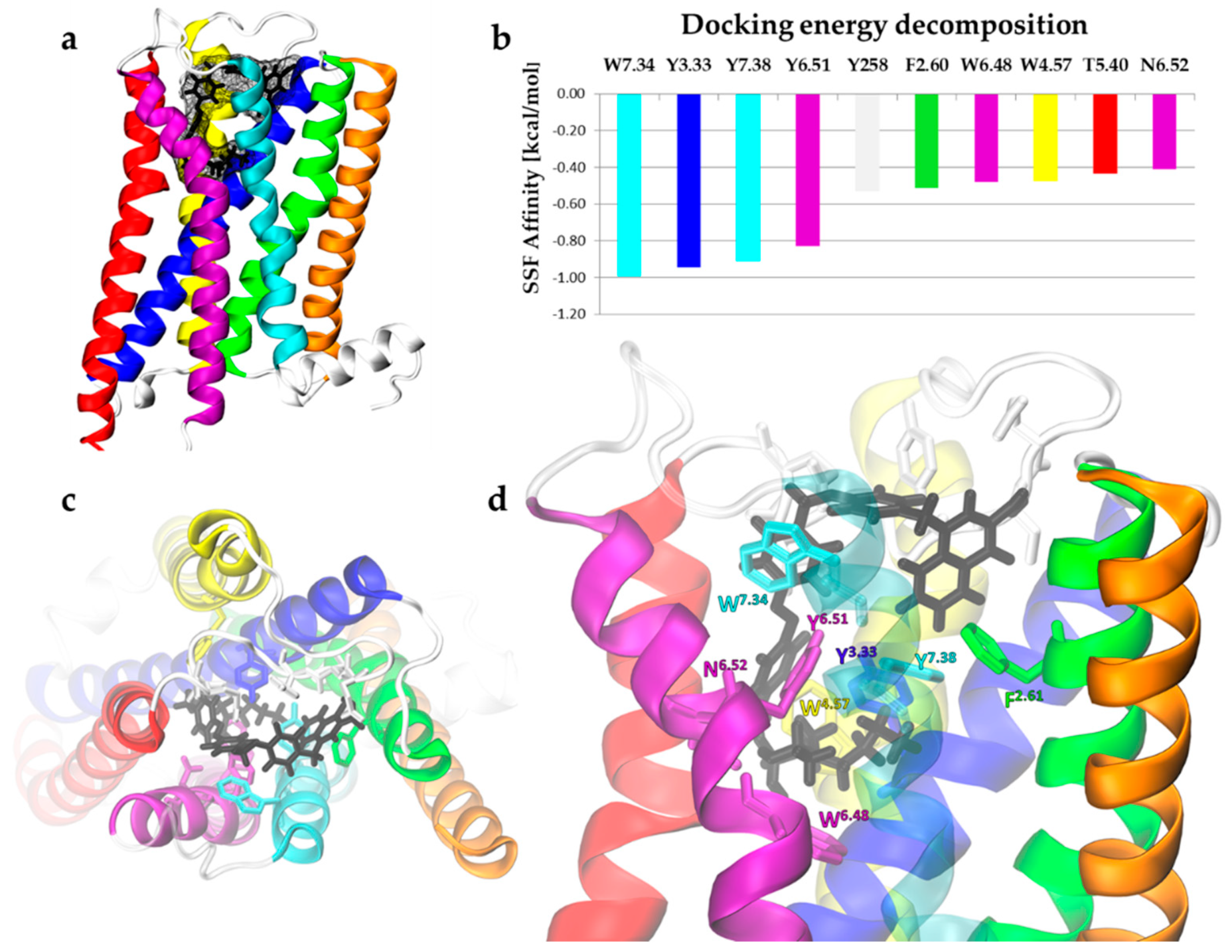
2.1.2. Single Ligands Docking to the Insect mAChR-A Model
2.2. What Happens upon Ligand Biding? Dynamical Response of Human M1 Receptor
2.2.1. GetContacts
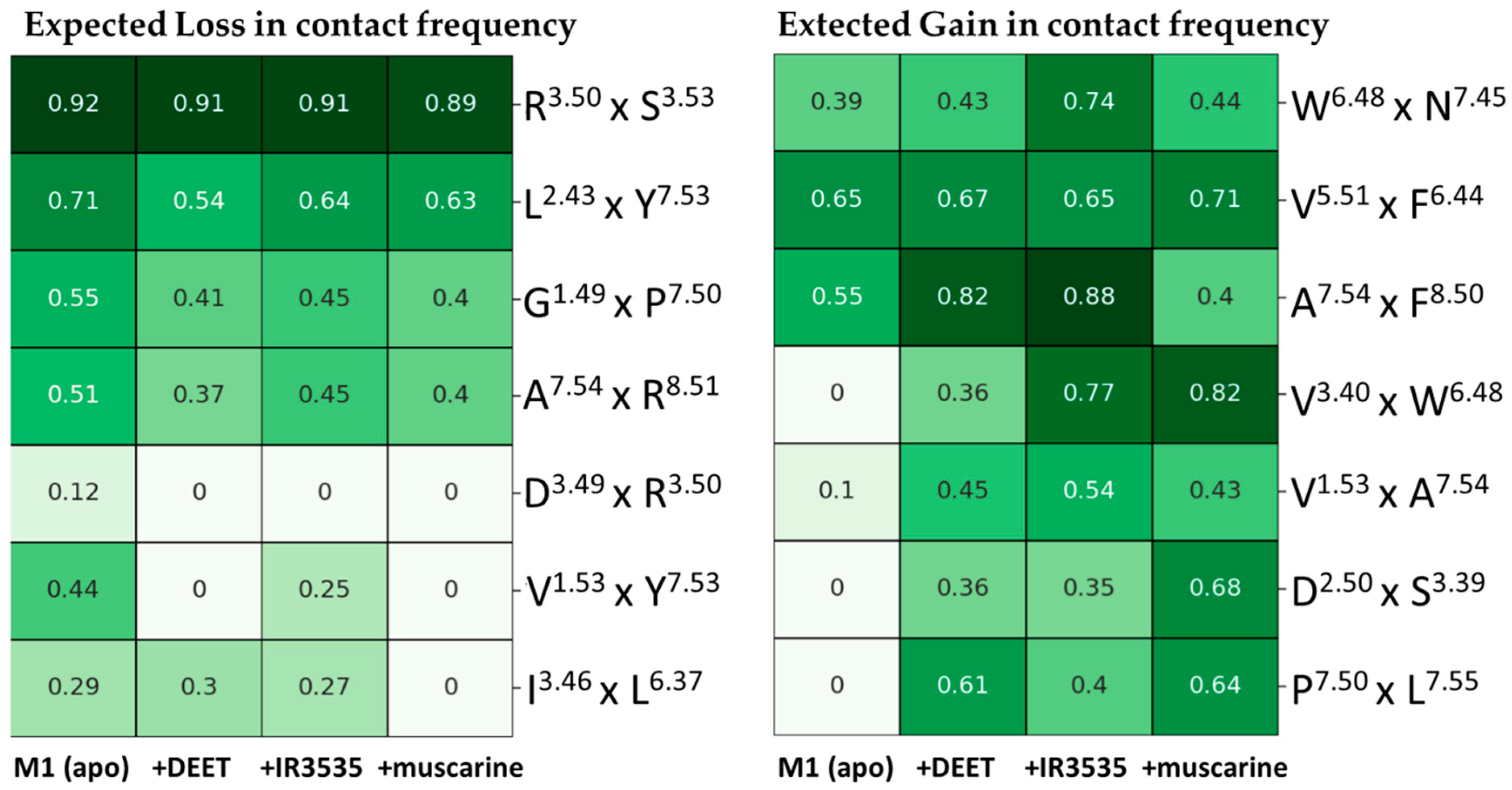
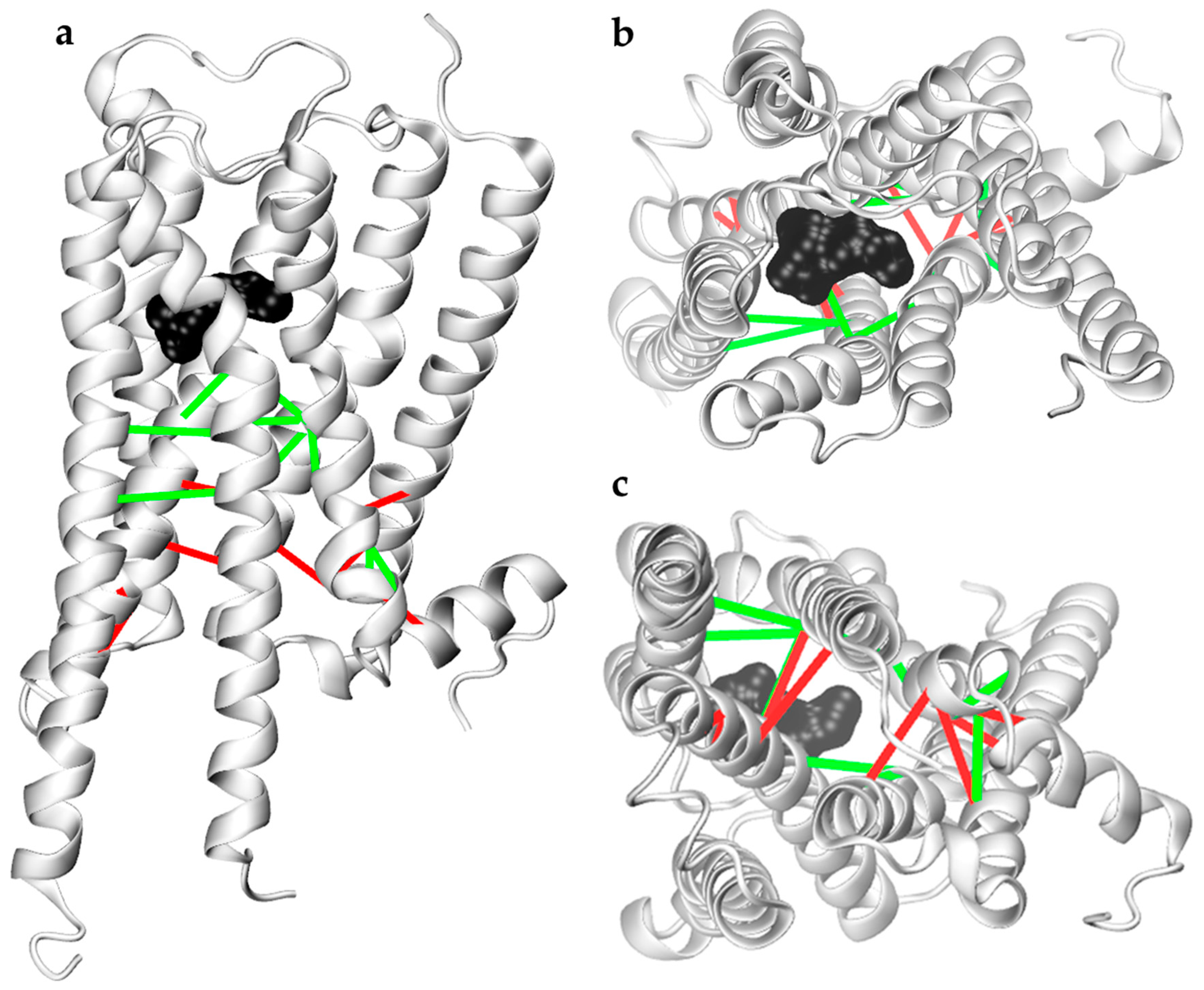
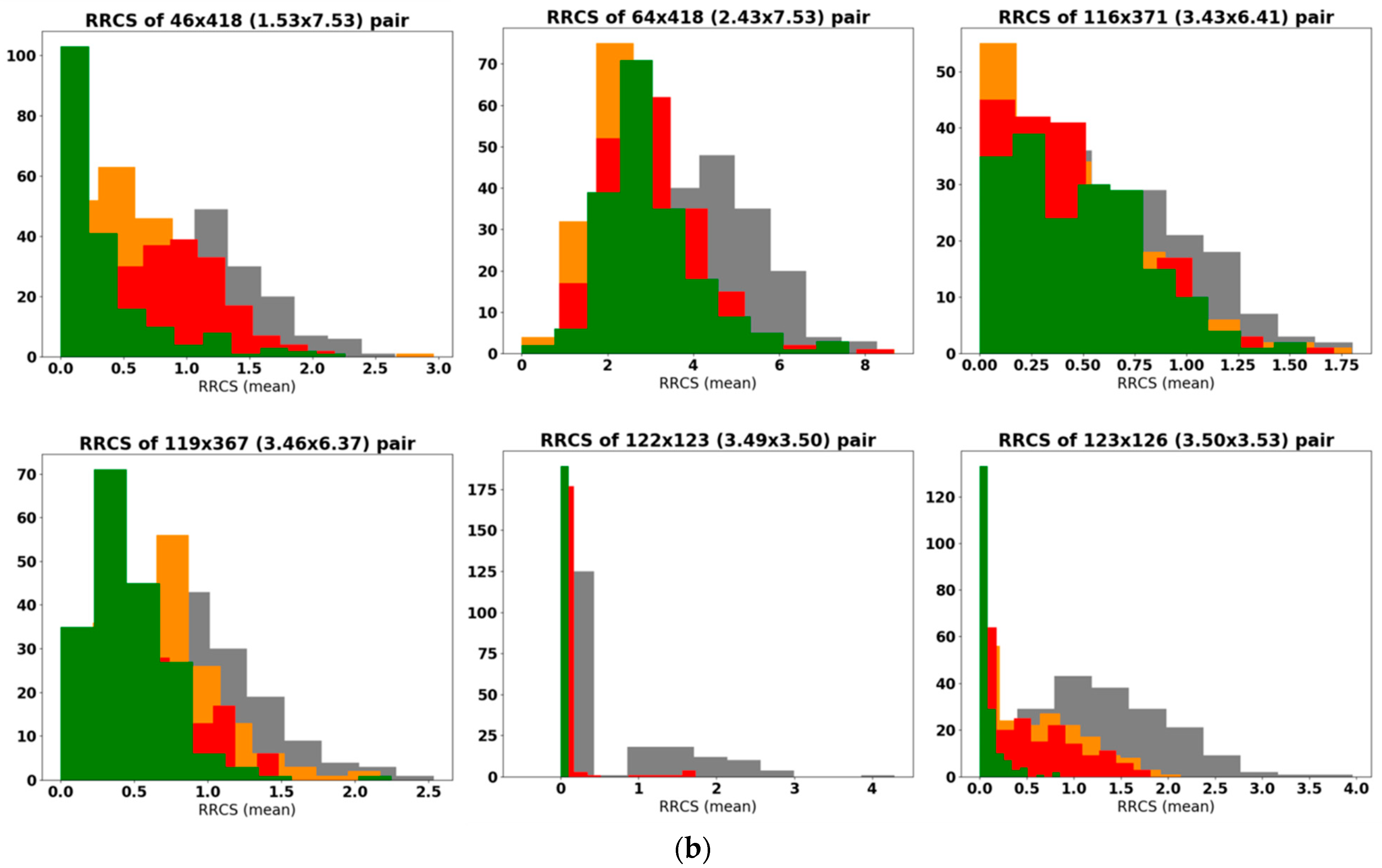
2.2.2. Analysis of Residue-Residue Contact Scores in Human M1
2.2.3. In Search for Repellent Modulation: Sequential Docking and Dynamics of the Human M1
2.3. Computer Modeling of Designed Bitopic Ligand (BQCA-azo-IR3535)
2.3.1. Bitopic BQCA-azo-IR3535 Ligand Effect on Human M1 GPCR
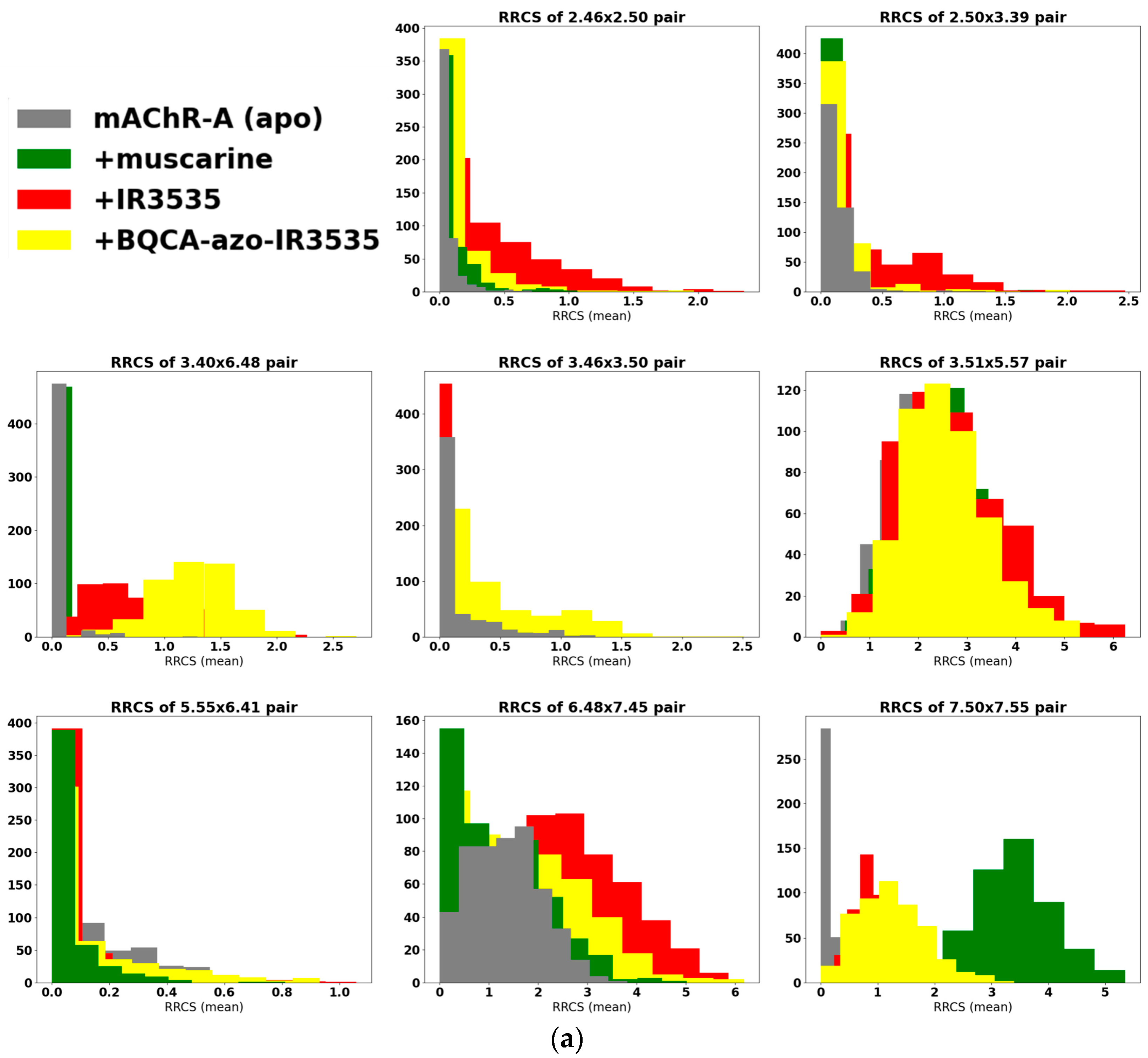
2.3.2. Bitopic BQCA-azo-IR3535 Ligand and Insect mAChR-A Dynamics
3. Conclusions
4. Materials and Methods
4.1. Molecular Docking
4.2. Molecular Dynamics (MD)
4.3. Homology Modeling
4.4. Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Campbell-Lendrum, D.; Manga, L.; Bagayoko, M.; Sommerfeld, J. Climate change and vector-borne diseases: What are the implications for public health research and policy? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20130552. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Wilke, A.B.; Bloomquist, J.R.; Desneux, N.; Beier, J.C. Overexposing mosquitoes to insecticides under global warming: A public health concern? Sci. Total Environ. 2021, 762, 143069. [Google Scholar] [CrossRef]
- Hemingway, J. Resistance: A problem without an easy solution. Pestic. Biochem. Physiol. 2018, 151, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Siegert, P.; Amimo, F.; Walker, E. Designation of chemicals in terms of the locomotor responses they elicit from insects: An update of Dethier et al.(1960). J. Econ. Entomol. 2009, 102, 2056–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, M.; da Silva, M.R.M.; de Siqueira, L.B.d.O.; Rodrigues, R.A.S.; Bodjolle-d’Almeida, L.; Dos Santos, E.P.; Ricci-Júnior, E. Trends in insect repellent formulations: A review. Int. J. Pharm. 2018, 539, 190–209. [Google Scholar] [CrossRef]
- Lupi, E.; Hatz, C.; Schlagenhauf, P. The efficacy of repellents against Aedes, Anopheles, Culex and Ixodes spp.—A literature review. Travel Med. Infect. Dis. 2013, 11, 374–411. [Google Scholar] [CrossRef]
- Corbel, V.; Stankiewicz, M.; Pennetier, C.; Fournier, D.; Stojan, J.; Girard, E.; Dimitrov, M.; Molgo, J.; Hougard, J.M.; Lapied, B. Evidence for inhibition of cholinesterases in insect and mammalian nervous systems by the insect repellent deet. BMC Biol. 2009, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Swale, D.R.; Sun, B.; Tong, F.; Bloomquist, J.R. Neurotoxicity and mode of action of N, N-diethyl-meta-toluamide (DEET). PLoS ONE 2014, 9, e103713. [Google Scholar] [CrossRef] [Green Version]
- Legeay, S.; Clere, N.; Hilairet, G.; Do, Q.T.; Bernard, P.; Quignard, J.F.; Apaire-Marchais, V.; Lapied, B.; Faure, S. The insect repellent N,N-diethyl-m-toluamide (DEET) induces angiogenesis via allosteric modulation of the M3 muscarinic receptor in endothelial cells. Sci. Rep. 2016, 6, 28546. [Google Scholar] [CrossRef] [Green Version]
- Osimitz, T.; Murphy, J.; Fell, L.; Page, B. Adverse events associated with the use of insect repellents containing N, N-diethyl-m-toluamide (DEET). Regul. Toxicol. Pharmacol. 2010, 56, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Legeay, S.; Clere, N.; Apaire-Marchais, V.; Faure, S.; Lapied, B. Unusual modes of action of the repellent DEET in insects highlight some human side effects. Eur. J. Pharmacol. 2018, 825, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Bohbot, J.D.; Dickens, J.C. Odorant receptor modulation: Ternary paradigm for mode of action of insect repellents. Neuropharmacology 2012, 62, 2086–2095. [Google Scholar] [CrossRef] [PubMed]
- Sanford, J.L.; Barski, S.A.; Seen, C.M.; Dickens, J.C.; Shields, V.D. Neurophysiological and behavioral responses of gypsy moth larvae to insect repellents: DEET, IR3535, and Picaridin. PLoS ONE 2014, 9, e99924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeGennaro, M. The mysterious multi-modal repellency of DEET. Fly 2015, 9, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Sparks, J.T.; Dickens, J.C. Bitter-sensitive gustatory receptor neuron responds to chemically diverse insect repellents in the common malaria mosquito Anopheles quadrimaculatus. Sci. Nat. 2016, 103, 39. [Google Scholar] [CrossRef]
- Sparks, J.T.; Dickens, J.C. Mini review: Gustatory reception of chemicals affecting host feeding in aedine mosquitoes. Pestic. Biochem. Physiol. 2017, 142, 15–20. [Google Scholar] [CrossRef]
- Deshayes, C.; Moreau, E.; Pitti-Caballero, J.; Froger, J.-A.; Apaire-Marchais, V.; Lapied, B. Synergistic agent and intracellular calcium, a successful partnership in the optimization of insecticide efficacy. Curr. Opin. Insect Sci. 2018, 30, 52–58. [Google Scholar] [CrossRef]
- Abd-Ella, A.; Stankiewicz, M.; Mikulska, K.; Nowak, W.; Pennetier, C.; Goulu, M.; Fruchart-Gaillard, C.; Licznar, P.; Apaire-Marchais, V.; List, O.; et al. The Repellent DEET Potentiates Carbamate Effects via Insect Muscarinic Receptor Interactions: An Alternative Strategy to Control Insect Vector-Borne Diseases. PLoS ONE 2015, 10, e0126406. [Google Scholar] [CrossRef] [Green Version]
- Moreau, E.; Mikulska-Ruminska, K.; Goulu, M.; Perrier, S.; Deshayes, C.; Stankiewicz, M.; Apaire-Marchais, V.; Nowak, W.; Lapied, B. Orthosteric muscarinic receptor activation by the insect repellent IR3535 opens new prospects in insecticide-based vector control. Sci. Rep. 2020, 10, 6842. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, M.; Lapied, B.; Jankowski, W.; Stankiewicz, M. The unusual action of essential oil component, menthol, in potentiating the effect of the carbamate insecticide, bendiocarb. Pestic. Biochem. Physiol. 2019, 158, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Buckingham, S.D.; Matsuda, K.; Sattelle, D.B. Modes of action, resistance and toxicity of insecticides targeting nicotinic acetylcholine receptors. Curr. Med. Chem. 2017, 24, 2925–2934. [Google Scholar] [CrossRef] [PubMed]
- Casida, J.E. Neonicotinoids and other insect nicotinic receptor competitive modulators: Progress and prospects. Annu. Rev. Entomol. 2018, 63, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid insecticides: Molecular targets, resistance, and toxicity. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Collin, C.; Hauser, F.; de Valdivia, E.G.; Li, S.; Reisenberger, J.; Carlsen, E.M.; Khan, Z.; Hansen, N.Ø.; Puhm, F.; Søndergaard, L. Two types of muscarinic acetylcholine receptors in Drosophila and other arthropods. Cell. Mol. Life Sci. 2013, 70, 3231–3242. [Google Scholar] [CrossRef]
- Xia, R.Y.; Li, M.Q.; Wu, Y.S.; Qi, Y.X.; Ye, G.Y.; Huang, J. A new family of insect muscarinic acetylcholine receptors. Insect Mol. Biol. 2016, 25, 362–369. [Google Scholar] [CrossRef]
- Pilon, A.; Goven, D.; Raymond, V. Pharmacological and molecular characterization of the A-type muscarinic acetylcholine receptor from Anopheles gambiae. Insect Mol. Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.R.; Folke, J.; Hauser, F.; Li, S.; Grimmelikhuijzen, C.J. The A-and B-type muscarinic acetylcholine receptors from Drosophila melanogaster couple to different second messenger pathways. Biochem. Biophys. Res. Commun. 2015, 462, 358–364. [Google Scholar] [CrossRef]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, E.; Kumari, P.; Jaiman, D.; Shukla, A.K. Methodological advances: The unsung heroes of the GPCR structural revolution. Nat. Rev. Mol. Cell Biol. 2015, 16, 69–81. [Google Scholar] [CrossRef]
- Congreve, M.; de Graaf, C.; Swain, N.A.; Tate, C.G. Impact of GPCR structures on drug discovery. Cell 2020, 181, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, A. Advances in G protein-coupled receptor allostery: From function to structure. Mol. Pharmacol. 2014, 86, 463–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thal, D.M.; Glukhova, A.; Sexton, P.M.; Christopoulos, A. Structural insights into G-protein-coupled receptor allostery. Nature 2018, 559, 45–53. [Google Scholar] [CrossRef]
- Valant, C.; Gregory, K.J.; Hall, N.E.; Scammells, P.J.; Lew, M.J.; Sexton, P.M.; Christopoulos, A. A Novel Mechanism of G Protein-coupled Receptor Functional Selectivity: MUSCARINIC PARTIAL AGONIST McN-A-343 AS A BITOPIC ORTHOSTERIC/ALLOSTERIC LIGAND∗. J. Biol. Chem. 2008, 283, 29312–29321. [Google Scholar] [CrossRef] [Green Version]
- Davie, B.J.; Christopoulos, A.; Scammells, P.J. Development of M1 mAChR allosteric and bitopic ligands: Prospective therapeutics for the treatment of cognitive deficits. ACS Chem. Neurosci. 2013, 4, 1026–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keov, P.; López, L.; Devine, S.M.; Valant, C.; Lane, J.R.; Scammells, P.J.; Sexton, P.M.; Christopoulos, A. Molecular mechanisms of bitopic ligand engagement with the M1 muscarinic acetylcholine receptor. J. Biol. Chem. 2014, 289, 23817–23837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, J.; Kellershohn, K.; Mohr-Andrä, M.; Kebig, A.; Prilla, S.; Muth, M.; Heller, E.; Disingrini, T.; Dallanoce, C.; Bertoni, S. Dualsteric GPCR targeting: A novel route to binding and signaling pathway selectivity. FASEB J. 2009, 23, 442–450. [Google Scholar] [CrossRef]
- Mohr, K.; Tränkle, C.; Kostenis, E.; Barocelli, E.; De Amici, M.; Holzgrabe, U. Rational design of dualsteric GPCR ligands: Quests and promise. Br. J. Pharmacol. 2010, 159, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Agnetta, L.; Kauk, M.; Canizal, M.C.A.; Messerer, R.; Holzgrabe, U.; Hoffmann, C.; Decker, M. A photoswitchable dualsteric ligand controlling receptor efficacy. Angew. Chem. Int. Ed. 2017, 56, 7282–7287. [Google Scholar] [CrossRef]
- Hoorens, M.W.; Szymanski, W. Reversible, spatial and temporal control over protein activity using light. Trends Biochem. Sci. 2018, 43, 567–575. [Google Scholar] [CrossRef]
- Ricart-Ortega, M.; Font, J.; Llebaria, A. GPCR photopharmacology. Mol. Cell. Endocrinol. 2019, 488, 36–51. [Google Scholar] [CrossRef]
- Nargeot, J.; Lester, H.A.; Birdsall, N.; Stockton, J.; Wassermann, N.H.; Erlanger, B.F. A photoisomerizable muscarinic antagonist. Studies of binding and of conductance relaxations in frog heart. J. Gen. Physiol. 1982, 79, 657–678. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common activation mechanism of class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef]
- Filipek, S. Molecular switches in GPCRs. Curr. Opin. Struct. Biol. 2019, 55, 114–120. [Google Scholar] [CrossRef]
- Katritch, V.; Fenalti, G.; Abola, E.E.; Roth, B.L.; Cherezov, V.; Stevens, R.C. Allosteric sodium in class A GPCR signaling. Trends Biochem. Sci. 2014, 39, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Koes, D.R.; Baumgartner, M.P.; Camacho, C.J. Lessons learned in empirical scoring with smina from the CSAR 2011 benchmarking exercise. J. Chem. Inf. Modeling 2013, 53, 1893–1904. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Ngan, C.-H.; Hall, D.R.; Zerbe, B.; Grove, L.E.; Kozakov, D.; Vajda, S. FTSite: High accuracy detection of ligand binding sites on unbound protein structures. Bioinformatics 2012, 28, 286–287. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Hulme, E.; Lu, Z.; Bee, M. Scanning mutagenesis studies of the M 1 muscarinic acetylcholine receptor. Recept. Channels 2003, 9, 215–228. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Weinstein, H. [19] Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. In Methods in Neurosciences; Academic Press: New York, NY, USA, 1995; Volume 25, pp. 366–428. [Google Scholar] [CrossRef]
- Venkatakrishnan, A.; Fonseca, R.; Ma, A.K.; Hollingsworth, S.A.; Chemparathy, A.; Hilger, D.; Kooistra, A.J.; Ahmari, R.; Babu, M.M.; Kobilka, B.K. Uncovering patterns of atomic interactions in static and dynamic structures of proteins. bioRxiv 2019, 840694. [Google Scholar] [CrossRef]
- Riefolo, F.; Matera, C.; Garrido-Charles, A.; Gomila, A.M.; Sortino, R.; Agnetta, L.; Claro, E.; Masgrau, R.; Holzgrabe, U.; Batlle, M. Optical control of cardiac function with a photoswitchable muscarinic agonist. J. Am. Chem. Soc. 2019, 141, 7628–7636. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Raniolo, S.; Limongelli, V.; Xu, Y. The molecular mechanism underlying ligand binding to the membrane-embedded site of a G-protein-coupled receptor. J. Chem. Theory Comput. 2018, 14, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Dawaliby, R.; Trubbia, C.; Delporte, C.; Noyon, C.; Ruysschaert, J.-M.; Van Antwerpen, P.; Govaerts, C. Phosphatidylethanolamine is a key regulator of membrane fluidity in eukaryotic cells. J. Biol. Chem. 2016, 291, 3658–3667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Dávila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M. CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [Green Version]
- Ragoza, M.; Hochuli, J.; Idrobo, E.; Sunseri, J.; Koes, D.R. Protein–ligand scoring with convolutional neural networks. J. Chem. Inf. Modeling 2017, 57, 942–957. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.S.; Wang, J.; Palczewski, K.; Filipek, S.; Vogel, H.; Liu, Z.-J.; Yuan, S. Exploring a new ligand binding site of G protein-coupled receptors. Chem. Sci. 2018, 9, 6480–6489. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.S.; Xu, Y.; Tan, L.; Vogel, H.; Cheng, J.; Wu, D.; Yuan, S. Enhancing the signaling of GPCRs via orthosteric ions. ACS Cent. Sci. 2020, 6, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Draper-Joyce, C.J.; Verma, R.K.; Michino, M.; Shonberg, J.; Kopinathan, A.; Herenbrink, C.K.; Scammells, P.J.; Capuano, B.; Abramyan, A.M.; Thal, D.M. The action of a negative allosteric modulator at the dopamine D 2 receptor is dependent upon sodium ions. Sci. Rep. 2018, 8, 1208. [Google Scholar] [CrossRef] [Green Version]
- Livingston, K.E.; Traynor, J.R. Disruption of the Na+ ion binding site as a mechanism for positive allosteric modulation of the mu-opioid receptor. Proc. Natl. Acad. Sci. USA 2014, 111, 18369–18374. [Google Scholar] [CrossRef] [Green Version]
- Pravda, L.; Sehnal, D.; Toušek, D.; Navrátilová, V.; Bazgier, V.; Berka, K.; Svobodová Vařeková, R.; Koča, J.; Otyepka, M. MOLEonline: A web-based tool for analyzing channels, tunnels and pores (2018 update). Nucleic Acids Res. 2018, 46, W368–W373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chovancova, E.; Pavelka, A.; Benes, P.; Strnad, O.; Brezovsky, J.; Kozlikova, B.; Gora, A.; Sustr, V.; Klvana, M.; Medek, P. CAVER 3.0: A tool for the analysis of transport pathways in dynamic protein structures. PLoS Comput. Biol. 2012, 8, e1002708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, J.N.; Broadhurst, R.W.; Bostock, M.J.; Solt, A.; Jones, A.J.; Gabriel, F.; Tandale, A.; Shrestha, B.; Nietlispach, D. Conformational plasticity of ligand-bound and ternary GPCR complexes studied by 19 F NMR of the β 1-adrenergic receptor. Nat. Commun. 2020, 11, 669. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, S.A.; Dror, R.O. Molecular dynamics simulation for all. Neuron 2018, 99, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Latorraca, N.R.; Venkatakrishnan, A.; Dror, R.O. GPCR dynamics: Structures in motion. Chem. Rev. 2017, 117, 139–155. [Google Scholar] [CrossRef]
- Fleetwood, O.; Carlsson, J.; Delemotte, L. Identification of ligand-specific G protein-coupled receptor states and prediction of downstream efficacy via data-driven modeling. eLife 2021, 10, e60715. [Google Scholar] [CrossRef]
- Birgül Iyison, N.; Shahraki, A.; Kahveci, K.; Düzgün, M.B.; Gün, G. Are insect GPCRs ideal next-generation pesticides: Opportunities and challenges. FEBS J. 2021, 288, 2727–2745. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Zhou, Y.; Tanaka, I.; Yao, M. Roll: A new algorithm for the detection of protein pockets and cavities with a rolling probe sphere. Bioinformatics 2010, 26, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Zoete, V.; Cuendet, M.A.; Grosdidier, A.; Michielin, O. SwissParam: A fast force field generation tool for small organic molecules. J. Comput. Chem. 2011, 32, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Lomize, M.A.; Pogozheva, I.D.; Joo, H.; Mosberg, H.I.; Lomize, A.L. OPM database and PPM web server: Resources for positioning of proteins in membranes. Nucleic Acids Res. 2012, 40, D370–D376. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y. CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Lüthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of protein models with three-dimensional profiles. Nature 1992, 356, 83–85. [Google Scholar] [CrossRef]
- Pontius, J.; Richelle, J.; Wodak, S.J. Deviations from standard atomic volumes as a quality measure for protein crystal structures. J. Mol. Biol. 1996, 264, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Van Rossum, G.; Drake, F.L. Python reference manual. In Department of Computer Science [CS]; CWI: Amsterdam, The Netherlands, 1995; p. R9525. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | DEET | IR3535 | Muscarine |
|---|---|---|---|
| GE+ | 0.276 | 0.070 | 0.334 |
| GE− | 0.829 | 1.300 | 1.682 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niklas, B.; Lapied, B.; Nowak, W. In Search of Synergistic Insect Repellents: Modeling of Muscarinic GPCR Interactions with Classical and Bitopic Photoactive Ligands. Molecules 2022, 27, 3280. https://doi.org/10.3390/molecules27103280
Niklas B, Lapied B, Nowak W. In Search of Synergistic Insect Repellents: Modeling of Muscarinic GPCR Interactions with Classical and Bitopic Photoactive Ligands. Molecules. 2022; 27(10):3280. https://doi.org/10.3390/molecules27103280
Chicago/Turabian StyleNiklas, Beata, Bruno Lapied, and Wieslaw Nowak. 2022. "In Search of Synergistic Insect Repellents: Modeling of Muscarinic GPCR Interactions with Classical and Bitopic Photoactive Ligands" Molecules 27, no. 10: 3280. https://doi.org/10.3390/molecules27103280