
GlycoTAIL and FlexiTAIL as Half-Life Extension Modules for Recombinant Antibody Fragments


Abstract
:1. Introduction
2. Results
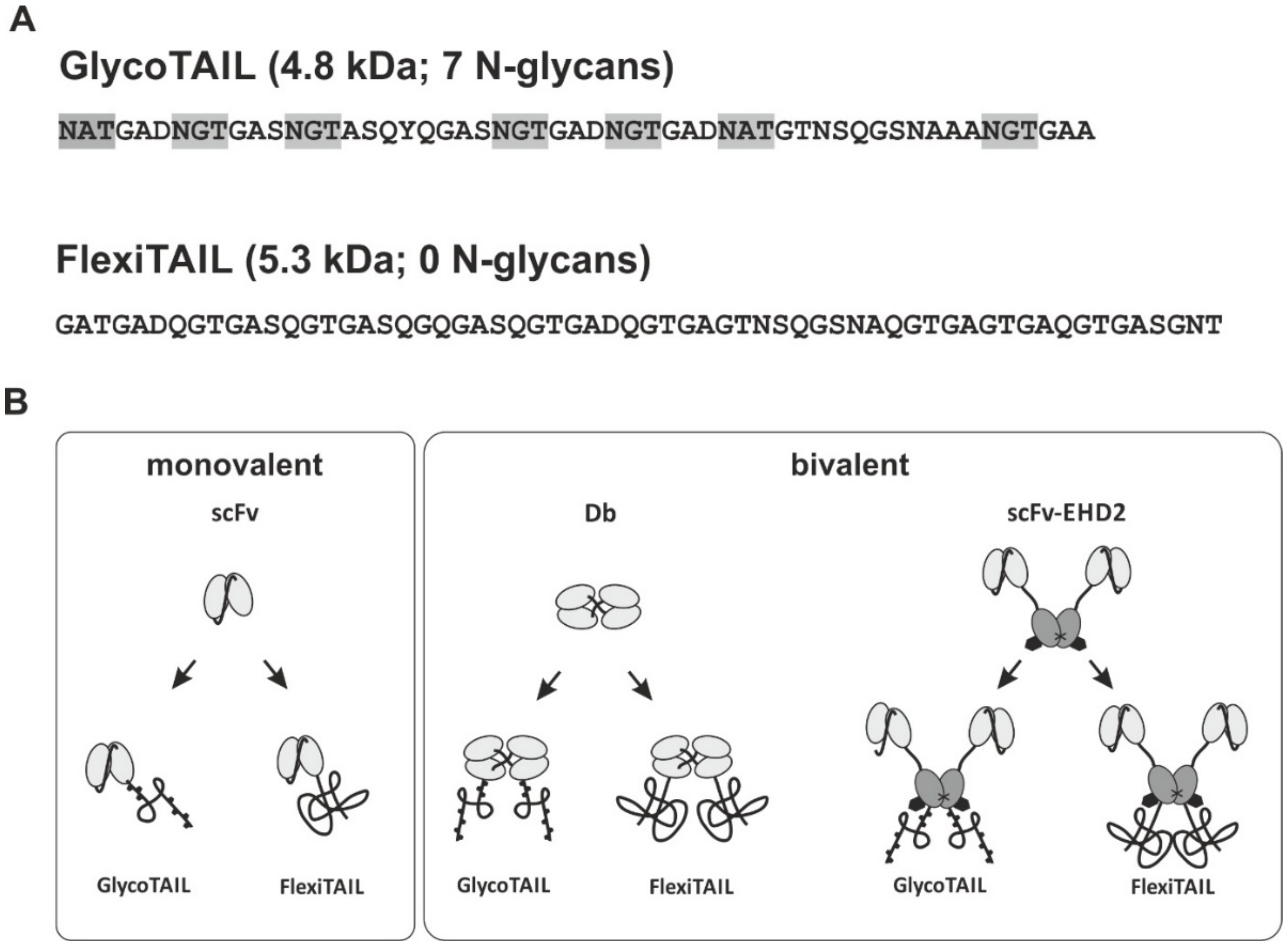
2.1. Generation of Antibody Fragments Comprising GlycoTAILs and FlexiTAILs
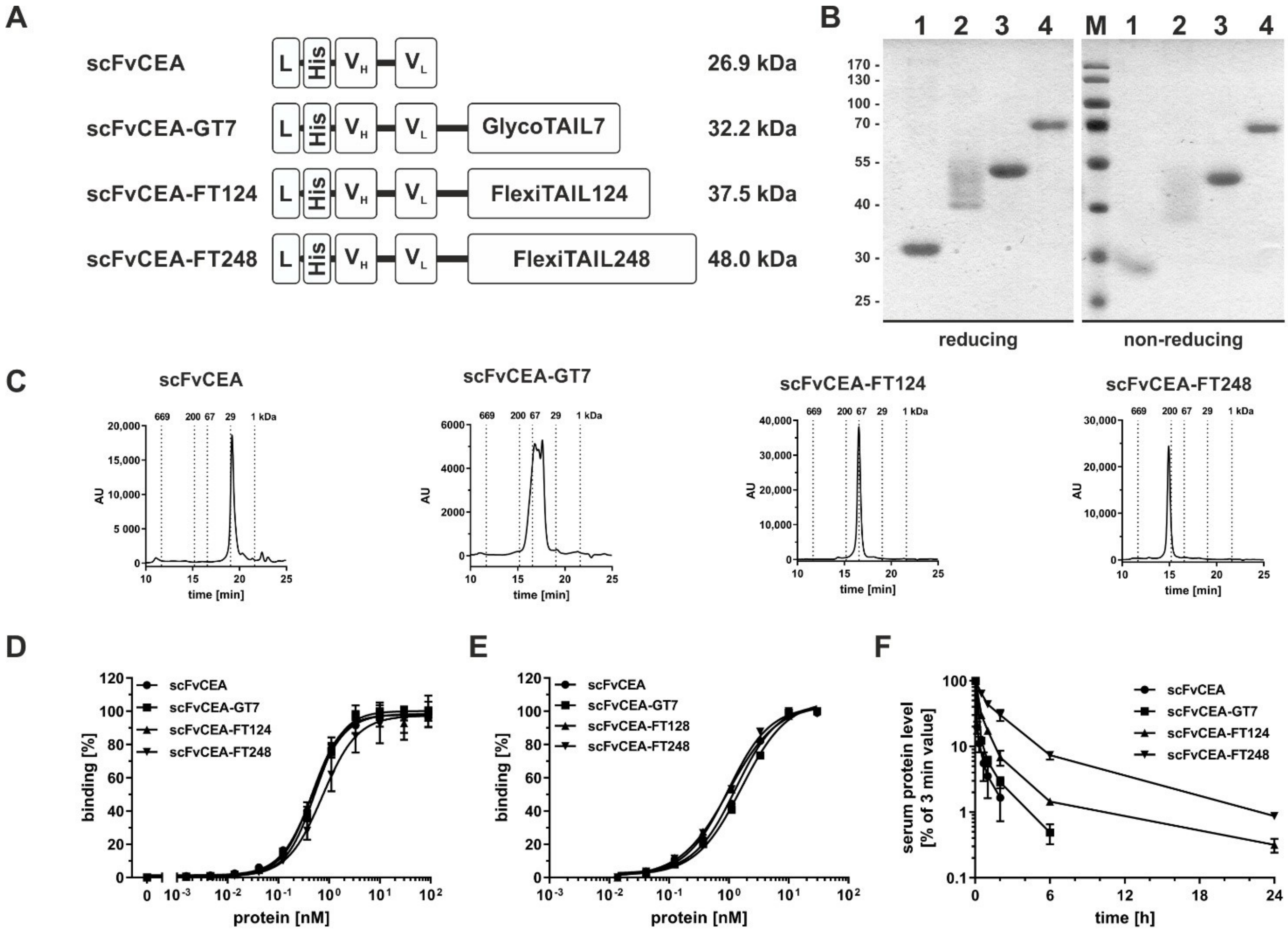
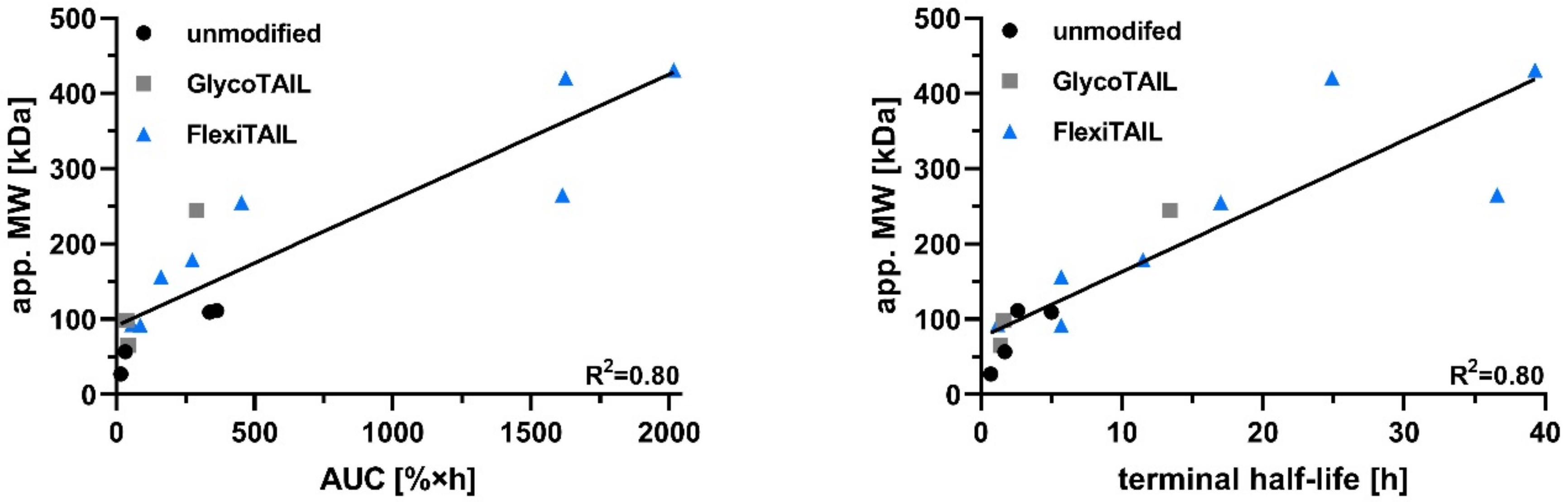
2.2. Half-Life Extension of an scFv
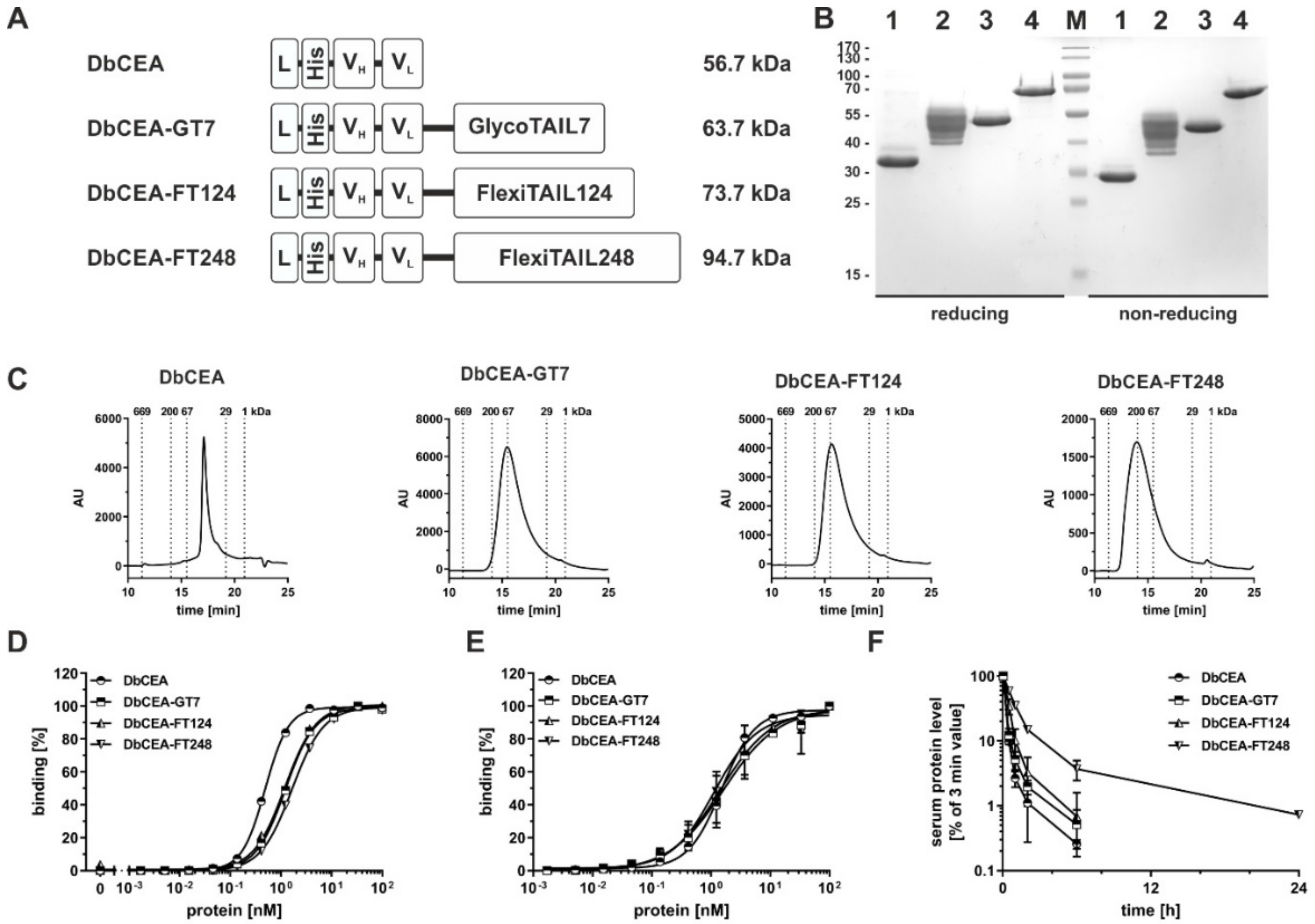
2.3. Half-Life Extension of a Bivalent Diabody
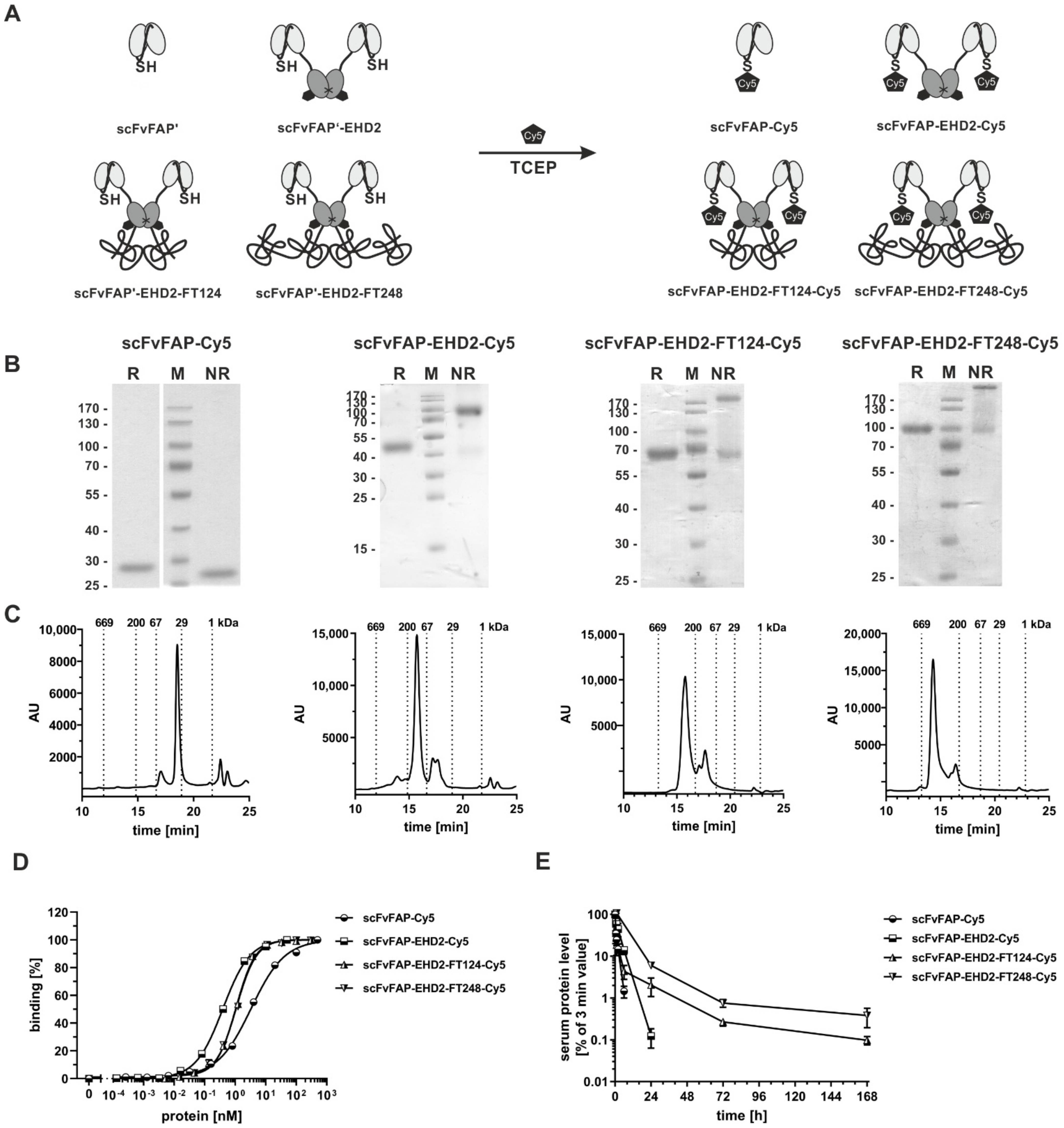
2.4. Half-Life Extension of a Bivalent scFv-EHD2 Fusion Protein
2.5. Generation of Cysteine-Modified Antibody Molecules
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Antibody Production and Purification
4.3. Antibody Characterization
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Flow Cytometry Analysis
4.6. Dynamic Light Scattering
4.7. Chemical Coupling of a Fluorophore
4.8. Pharmacokinetics
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Saeed, A.F.U.H.; Wang, R.; Ling, S.; Wang, S. Antibody Engineering for Pursuing a Healthier Future. Front. Microbiol. 2017, 8, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.L. Antibody fragments: Hope and hype. MAbs 2010, 2, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohl, W.R. Current progress in innovative engineered antibodies. Protein Cell 2018, 9, 86–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, A.; Power, C.A. David vs. Goliath: The Structure, Function, and Clinical Prospects of Antibody Fragments. Antibodies 2019, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitten, O.; Martineau, P. Les formats alternatifs aux anticorps-Fragments et nouvelles charpentes. Med. Sci. 2019, 35, 1092–1097. [Google Scholar] [CrossRef]
- Bitencourt, A.L.B.; Campos, R.M.; Cline, E.N.; Klein, W.L.; Sebollela, A. Antibody Fragments as Tools for Elucidating Structure-Toxicity Relationships and for Diagnostic/Therapeutic Targeting of Neurotoxic Amyloid Oligomers. Int. J. Mol. Sci. 2020, 21, 8920. [Google Scholar] [CrossRef]
- Freise, A.C.; Wu, A.M. In vivo imaging with antibodies and engineered fragments. Mol. Immunol. 2015, 67, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Deonarain, M.P.; Xue, Q. Tackling solid tumour therapy with small-format drug conjugates. Antib. Ther. 2020, 3, 237–245. [Google Scholar] [CrossRef]
- Richards, D.B.; Cookson, L.M.; Barton, S.V.; Liefaard, L.; Lane, T.; Hutt, D.F.; Ritter, J.M.; Fontana, M.; Moon, J.C.; Gillmore, J.D.; et al. Repeat doses of antibody to serum amyloid P component clear amyloid deposits in patients with systemic amyloidosis. Sci. Transl. Med. 2018, 10, eaan3128. [Google Scholar] [CrossRef] [Green Version]
- Kholodenko, R.V.; Kalinovsky, D.V.; Doronin, I.I.; Ponomarev, E.D.; Kholodenko, I.V. Antibody Fragments as Potential Biopharmaceuticals for Cancer Therapy: Success and Limitations. Curr. Med. Chem. 2019, 26, 396–426. [Google Scholar] [CrossRef]
- Kontermann, R.E. Half-life extended biotherapeutics. Expert Opin. Biol. Ther. 2016, 16, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Zaman, R.; Islam, R.A.; Ibnat, N.; Othman, I.; Zaini, A.; Lee, C.Y.; Chowdhury, E.H. Current strategies in extending half-lives of therapeutic proteins. J. Control. Release 2019, 301, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Kontermann, R.E. Strategies for extended serum half-life of protein therapeutics. Curr. Opin. Biotechnol. 2011, 22, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Buscaglia, C.A.; Campetella, O.; Leguizamón, M.S.; Frasch, A.C. The repetitive domain of Trypanosoma cruzi trans-sialidase enhances the immune response against the catalytic domain. J. Infect. Dis. 1998, 177, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, P.; Buscaglia, C.A.; Campetella, O. Improving protein pharmacokinetics by genetic fusion to simple amino acid sequences. J. Biol. Chem. 2004, 279, 3375–3381. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-S.; Wen, X.-F.; Wu, Y.-L.; Wang, Y.-F.; Fan, M.; Yang, Z.-Y.; Liu, W.; Zhou, L.-F. Engineering a pharmacologically superior form of granulocyte-colony-stimulating factor by fusion with gelatin-like-protein polymer. Eur. J. Pharm. Biopharm. 2010, 74, 435–441. [Google Scholar] [CrossRef]
- Shamji, M.F.; Betre, H.; Kraus, V.B.; Chen, J.; Chilkoti, A.; Pichika, R.; Masuda, K.; Setton, L.A. Development and characterization of a fusion protein between thermally responsive elastin-like polypeptide and interleukin-1 receptor antagonist: Sustained release of a local antiinflammatory therapeutic. Arthritis Rheum. 2007, 56, 3650–3661. [Google Scholar] [CrossRef]
- Schellenberger, V.; Wang, C.-W.; Geething, N.C.; Spink, B.J.; Campbell, A.; To, W.; Scholle, M.D.; Yin, Y.; Yao, Y.; Bogin, O.; et al. A recombinant polypeptide extends the in vivo half-life of peptides and proteins in a tunable manner. Nat. Biotechnol. 2009, 27, 1186–1190. [Google Scholar] [CrossRef]
- Schlapschy, M.; Binder, U.; Börger, C.; Theobald, I.; Wachinger, K.; Kisling, S.; Haller, D.; Skerra, A. PASylation: A biological alternative to PEGylation for extending the plasma half-life of pharmaceutically active proteins. Protein Eng. Des. Sel. 2013, 26, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Khodabakhsh, F.; Salimian, M.; Hedayati, M.H.; Ahangari Cohan, R.; Norouzian, D. Challenges and advancements in the pharmacokinetic enhancement of therapeutic proteins. Prep. Biochem. Biotechnol. 2021, 51, 519–529. [Google Scholar] [CrossRef]
- Stork, R.; Zettlitz, K.A.; Müller, D.; Rether, M.; Hanisch, F.-G.; Kontermann, R.E. N-glycosylation as novel strategy to improve pharmacokinetic properties of bispecific single-chain diabodies. J. Biol. Chem. 2008, 283, 7804–7812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stork, R.; Campigna, E.; Robert, B.; Müller, D.; Kontermann, R.E. Biodistribution of a bispecific single-chain diabody and its half-life extended derivatives. J. Biol. Chem. 2009, 284, 25612–25619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, O.; Plappert, A.; Fellermeier, S.; Siegemund, M.; Pfizenmaier, K.; Kontermann, R.E. Tetravalent antibody-scTRAIL fusion proteins with improved properties. Mol. Cancer Ther. 2014, 13, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, O.; Rau, A.; Beha, N.; Richter, F.; Kontermann, R.E. Diabody-Ig: A novel platform for the generation of multivalent and multispecific antibody molecules. MAbs 2019, 11, 919–929. [Google Scholar] [CrossRef]
- Fabre, M.; Ferrer, C.; Domínguez-Hormaetxe, S.; Bockorny, B.; Murias, L.; Seifert, O.; Eisler, S.A.; Kontermann, R.E.; Pfizenmaier, K.; Lee, S.Y.; et al. OMTX705, a Novel FAP-Targeting ADC Demonstrates Activity in Chemotherapy and Pembrolizumab-Resistant Solid Tumor Models. Clin. Cancer Res. 2020, 26, 3420–3430. [Google Scholar] [CrossRef] [Green Version]
- Kubetzko, S.; Sarkar, C.A.; Plückthun, A. Protein PEGylation decreases observed target association rates via a dual blocking mechanism. Mol. Pharmacol. 2005, 68, 1439–1454. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.S.; Conover, C.; Shi, C.; Whitlow, M.; Filpula, D. Prolonged circulating lives of single-chain Fv proteins conjugated with polyethylene glycol: A comparison of conjugation chemistries and compounds. Bioconjug. Chem. 1999, 10, 973–981. [Google Scholar] [CrossRef]
- Geething, N.C.; To, W.; Spink, B.J.; Scholle, M.D.; Wang, C.-W.; Yin, Y.; Yao, Y.; Schellenberger, V.; Cleland, J.L.; Stemmer, W.P.C.; et al. Gcg-XTEN: An improved glucagon capable of preventing hypoglycemia without increasing baseline blood glucose. PLoS ONE 2010, 5, e10175. [Google Scholar] [CrossRef] [Green Version]
- Cleland, J.L.; Geething, N.C.; Moore, J.A.; Rogers, B.C.; Spink, B.J.; Wang, C.-W.; Alters, S.E.; Stemmer, W.P.C.; Schellenberger, V. A novel long-acting human growth hormone fusion protein (VRS-317): Enhanced in vivo potency and half-life. J. Pharm. Sci. 2012, 101, 2744–2754. [Google Scholar] [CrossRef] [Green Version]
- Binder, U.; Skerra, A. Current Strategies for Pharmacokinetic Optimization. In Biobetters; Rosenberg, A., Demeule, B., Eds.; Springer: New York, NY, USA, 2015; pp. 269–311. ISBN 978-1-4939-2542-1. [Google Scholar]
- Schönfeld, D.; Matschiner, G.; Chatwell, L.; Trentmann, S.; Gille, H.; Hülsmeyer, M.; Brown, N.; Kaye, P.M.; Schlehuber, S.; Hohlbaum, A.M.; et al. An engineered lipocalin specific for CTLA-4 reveals a combining site with structural and conformational features similar to antibodies. Proc. Natl. Acad. Sci. USA 2009, 106, 8198–8203. [Google Scholar] [CrossRef] [Green Version]
- Amstutz, P.; Koch, H.; Binz, H.K.; Deuber, S.A.; Plückthun, A. Rapid selection of specific MAP kinase-binders from designed ankyrin repeat protein libraries. Protein Eng. Des. Sel. 2006, 19, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, F.; Merten, H.; Zimmermann, M.; Béhé, M.; Zangemeister-Wittke, U.; Plückthun, A. Influence of size and charge of unstructured polypeptides on pharmacokinetics and biodistribution of targeted fusion proteins. J. Control. Release 2019, 307, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Brandl, F.; Busslinger, S.; Zangemeister-Wittke, U.; Plückthun, A. Optimizing the anti-tumor efficacy of protein-drug conjugates by engineering the molecular size and half-life. J. Control. Release 2020, 327, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Olafsen, T.; Kenanova, V.E.; Wu, A.M. Tunable pharmacokinetics: Modifying the in vivo half-life of antibodies by directed mutagenesis of the Fc fragment. Nat. Protoc. 2006, 1, 2048–2060. [Google Scholar] [CrossRef] [PubMed]
- Mendler, C.T.; Friedrich, L.; Laitinen, I.; Schlapschy, M.; Schwaiger, M.; Wester, H.-J.; Skerra, A. High contrast tumor imaging with radio-labeled antibody Fab fragments tailored for optimized pharmacokinetics via PASylation. MAbs 2015, 7, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Bumbaca, B.; Li, Z.; Shah, D.K. Pharmacokinetics of protein and peptide conjugates. Drug Metab. Pharmacokinet. 2019, 34, 42–54. [Google Scholar] [CrossRef]
- Tsai, W.-T.K.; Wu, A.M. Aligning physics and physiology: Engineering antibodies for radionuclide delivery. J. Labelled Comp. Radiopharm. 2018, 61, 693–714. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| scFvCEA | scFvCEA-GT7 | scFvCEA-FT124 | scFvCEA-FT248 | |
|---|---|---|---|---|
| calc. MW [kDa] | 26.9 | 32.2 | 37.5 | 48.0 |
| app. MW [kDa] | 27.1 | 53.6–87.0 | 91.9 | 178.9 |
| agg. temp. [°C] | 46 | n.d. | 47 | 47 |
| ELISA [nM] | 0.5 ± 0.04 | 0.5 ± 0.05 | 0.5 ± 0.05 | 0.8 ± 0.14 |
| FACS [nM] | 1.3 ± 0.07 | 1.7 ± 0.10 | 1.7 ± 0.95 | 1.1 ± 0.18 |
| t½α [h] | 0.1 ± 0.01 | 0.2 ± 0.01 | 0.4 ± 0.02 | 0.8 ± 0.09 |
| t½β [h] | 0.7 ± 0.30 | 1.4 ± 0.09 | 5.7 ± 0.62 | 11.5 ± 0.64 |
| AUC [%×h] | 16 ± 4 | 42 ± 3 | 87 ± 4 | 275 ± 31 |
| DbCEA | DbCEA-GT7 | DbCEA-FT124 | DbCEA-FT248 | |
|---|---|---|---|---|
| calc. MW [kDa] | 56.7 | 63.7 | 73.7 | 94.7 |
| app. MW [kDa] | 56.8 | 98.7 | 93.3 | 156.5 |
| agg. temp. [°C] | 47 | 48 | 50 | 49 |
| ELISA [nM] | 0.6 ± 0.07 | 1.4 ± 0.19 | 1.3 ± 0.13 | 1.7 ± 0.11 |
| FACS [nM] | 1.5 ± 0.06 | 1.5 ± 0.26 | 1.3 ± 0.33 | 1.5 ± 0.65 |
| t½α [h] | 0.2 ± 0.01 | 0.2 ± 0.02 | 0.3 ± 0.06 | 0.6 ± 0.05 |
| t½β [h] | 1.7 ± 0.30 | 1.6 ± 0.50 | 1.2 ± 0.50 | 5.7 ± 0.50 |
| AUC [%×h] | 32 ± 2 | 38 ± 4 | 56 ± 15 | 162 ± 16 |
| scFvCEA-EHD2 | scFvCEA-EHD2-GT7 | scFvCEA-EHD2-FT124 | scFvCEA-EHD2-FT248 | |
|---|---|---|---|---|
| calc. MW [kDa] | 83.0 | 90.0 | 100.6 | 121.6 |
| app. MW [kDa] | 109.5 | 244.4 | 255.4 | 420.5 |
| agg. temp. [°C] | 46 | 47 | 48 | 47 |
| ELISA [nM] | 0.3 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.03 | 0.2 ± 0.03 |
| t½α [h] | 0.8 ± 0.18 | 0.4 ± 0.02 | 0.4 ± 0.03 | 2.4 ± 0.06 |
| t½β [h] | 5.0 ± 0.4 | 13.4 ± 1.4 | 17.0 ± 4.0 | 24.9 ± 2.7 |
| AUC [%×h] | 337 ± 45 | 290 ± 80 | 453 ± 54 | 1625 ± 159 |
| scFvFAP-Cy5 | scFvFAP-EHD2-Cy5 | scFvFAP-EHD2-FT124-Cy5 | scFvFAP-EHD2-FT248-Cy5 | |
|---|---|---|---|---|
| calc. MW [kDa] | 28.9 | 84.4 | 101.8 | 122.8 |
| app. MW [kDa] | 32.9 | 111.5 | 265.0 | 431.0 |
| FACS [nM] | 3.3 ± 0.03 | 0.4 ± 0.02 | 1.1 ± 0.05 | 1.1 ± 0.01 |
| t½α [h] | 0.5 ± 0.01 | 1.5 ± 1.0 | 0.4 ± 0.11 | n.d. |
| t½β [h] | 1.2 ± 0.03 | 2.6 ± 0.3 | 36.6 ± 9.4 | 39.3 ± 6.5 |
| AUC [%×h] | 98 ± 14 | 363 ± 69 | 1613 ± 234 | 2016 ± 142 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seifert, O.; Kontermann, R.E. GlycoTAIL and FlexiTAIL as Half-Life Extension Modules for Recombinant Antibody Fragments. Molecules 2022, 27, 3272. https://doi.org/10.3390/molecules27103272
Seifert O, Kontermann RE. GlycoTAIL and FlexiTAIL as Half-Life Extension Modules for Recombinant Antibody Fragments. Molecules. 2022; 27(10):3272. https://doi.org/10.3390/molecules27103272
Chicago/Turabian StyleSeifert, Oliver, and Roland E. Kontermann. 2022. "GlycoTAIL and FlexiTAIL as Half-Life Extension Modules for Recombinant Antibody Fragments" Molecules 27, no. 10: 3272. https://doi.org/10.3390/molecules27103272