
Polyphenols Extracted from Shanxi-Aged Vinegar Inhibit Inflammation in LPS-Induced RAW264.7 Macrophages and ICR Mice via the Suppression of MAPK/NF-κB Pathway Activation

 ,
,
Abstract
:1. Introduction
2. Results
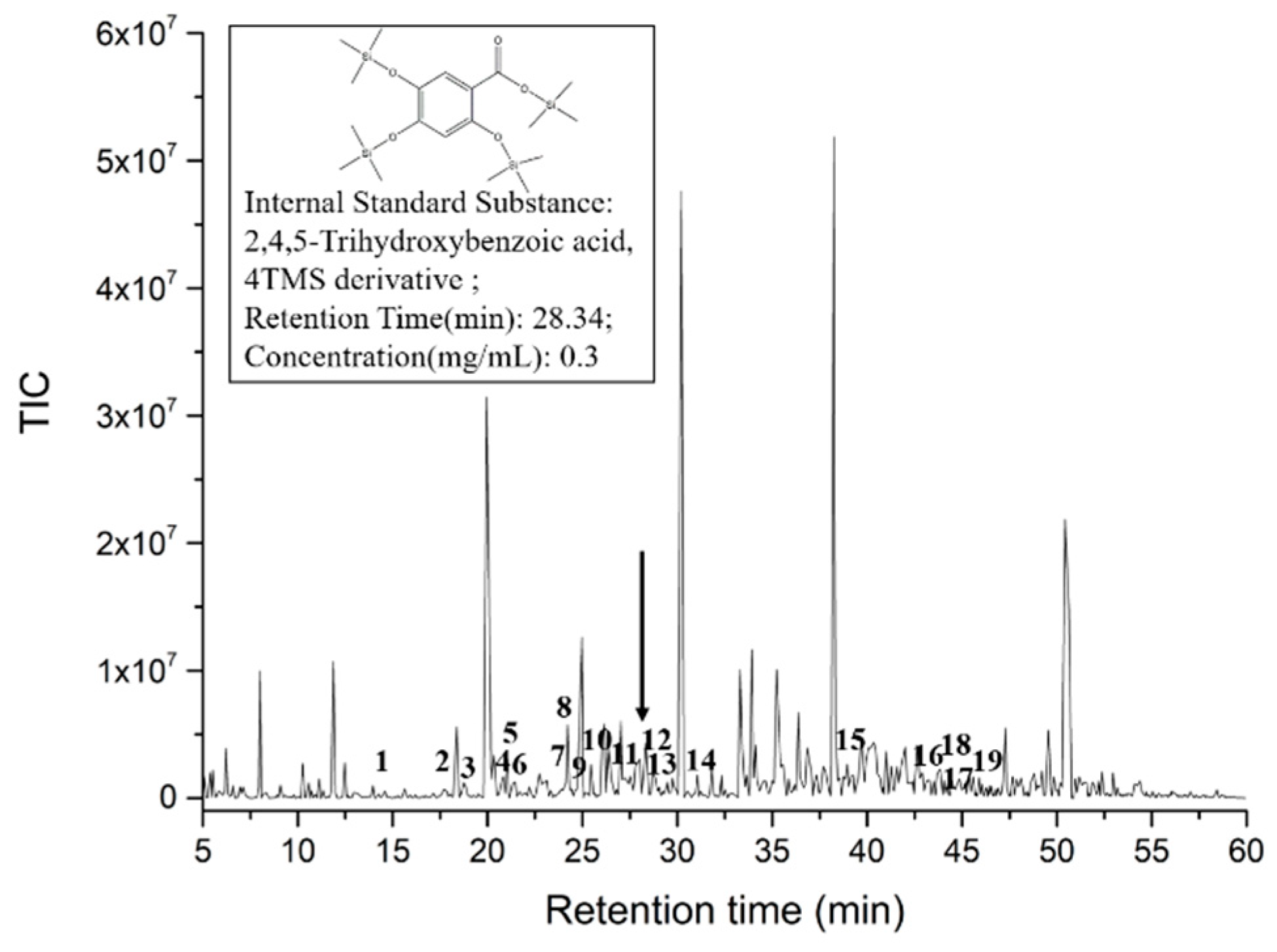
2.1. Analysis of SAVEP
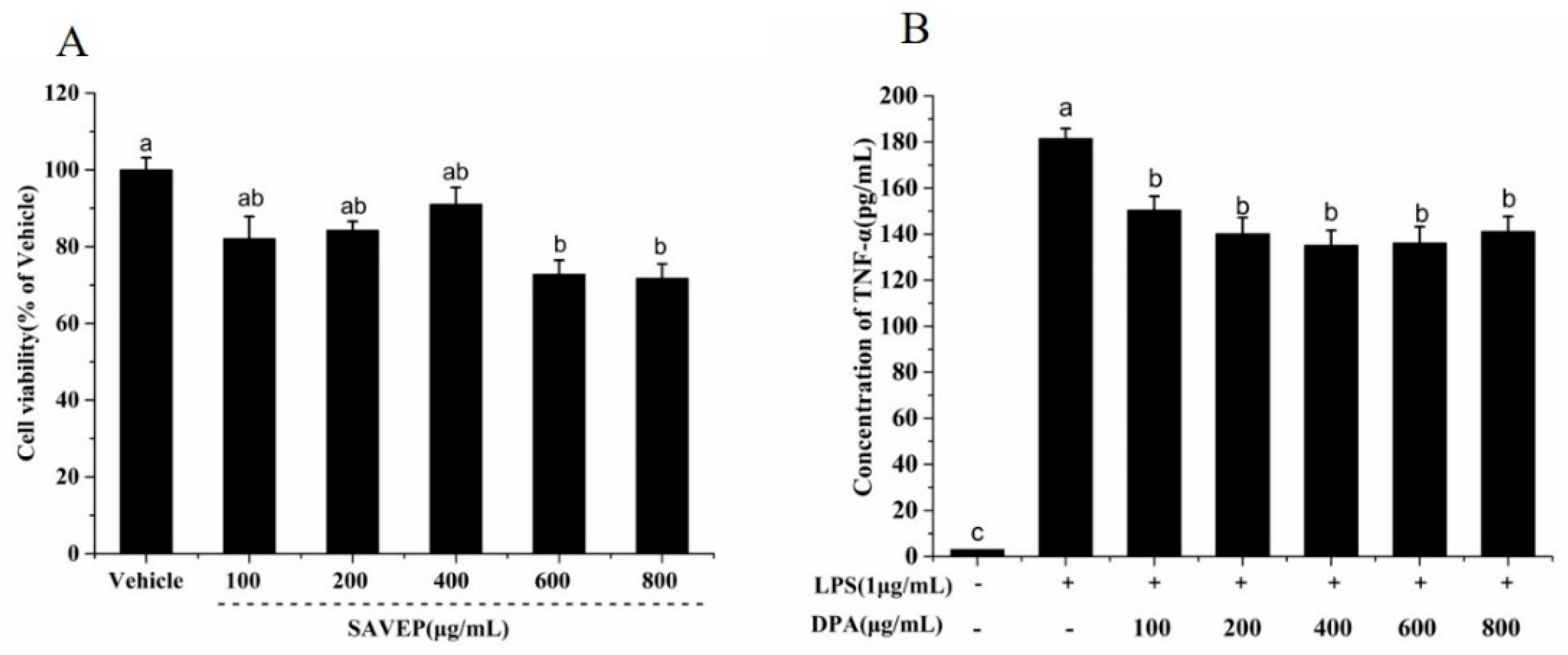
2.2. Effects of Different SAVEP Concentrations on Cell Viability and Dose Selection
2.3. Effects of SAVEP on LPS-Induced Inflammatory Cell Morphology
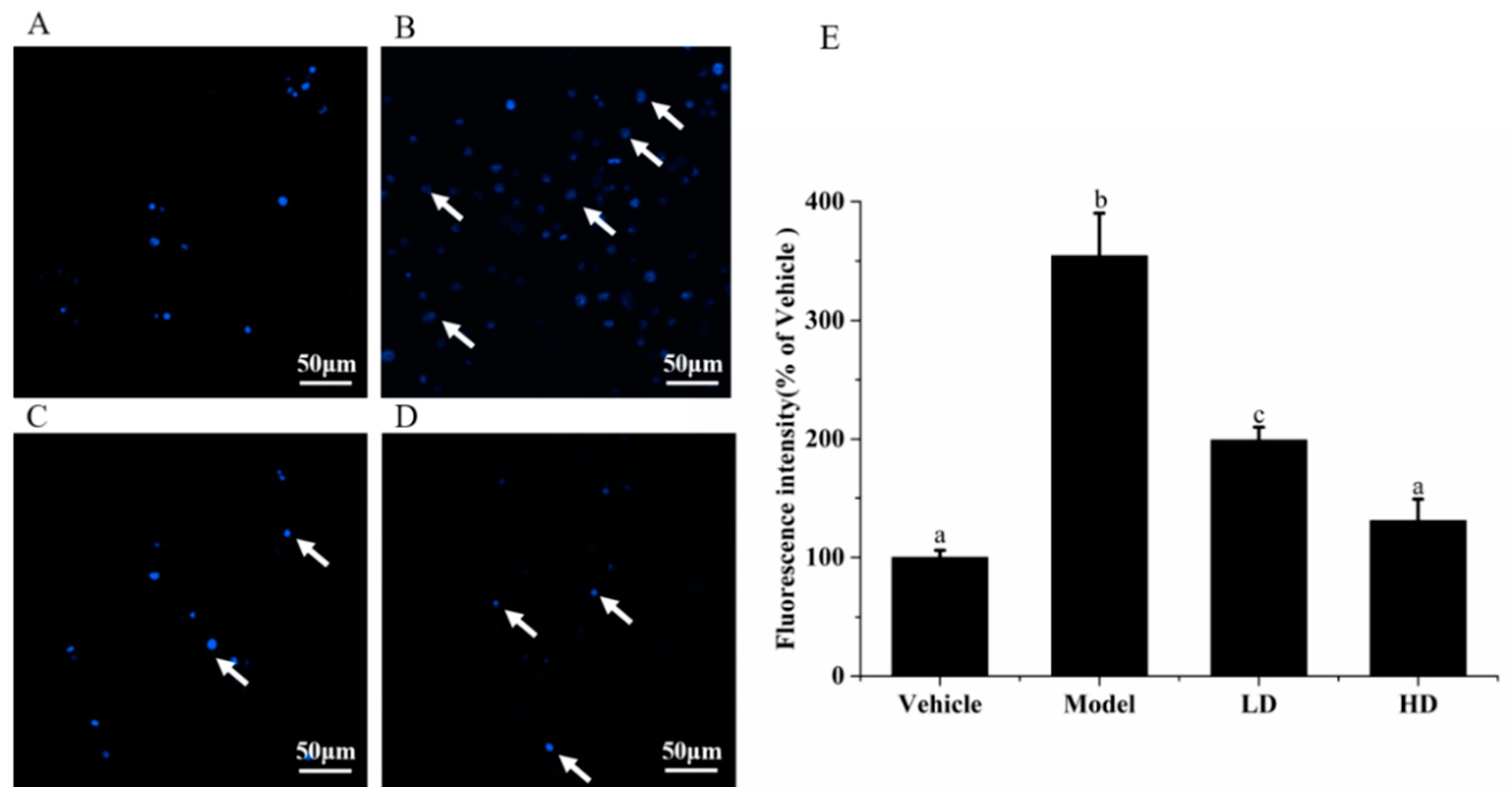
2.4. Effects of SAVEP on LPS-Induced Inflammatory Cell Nucleus Morphology
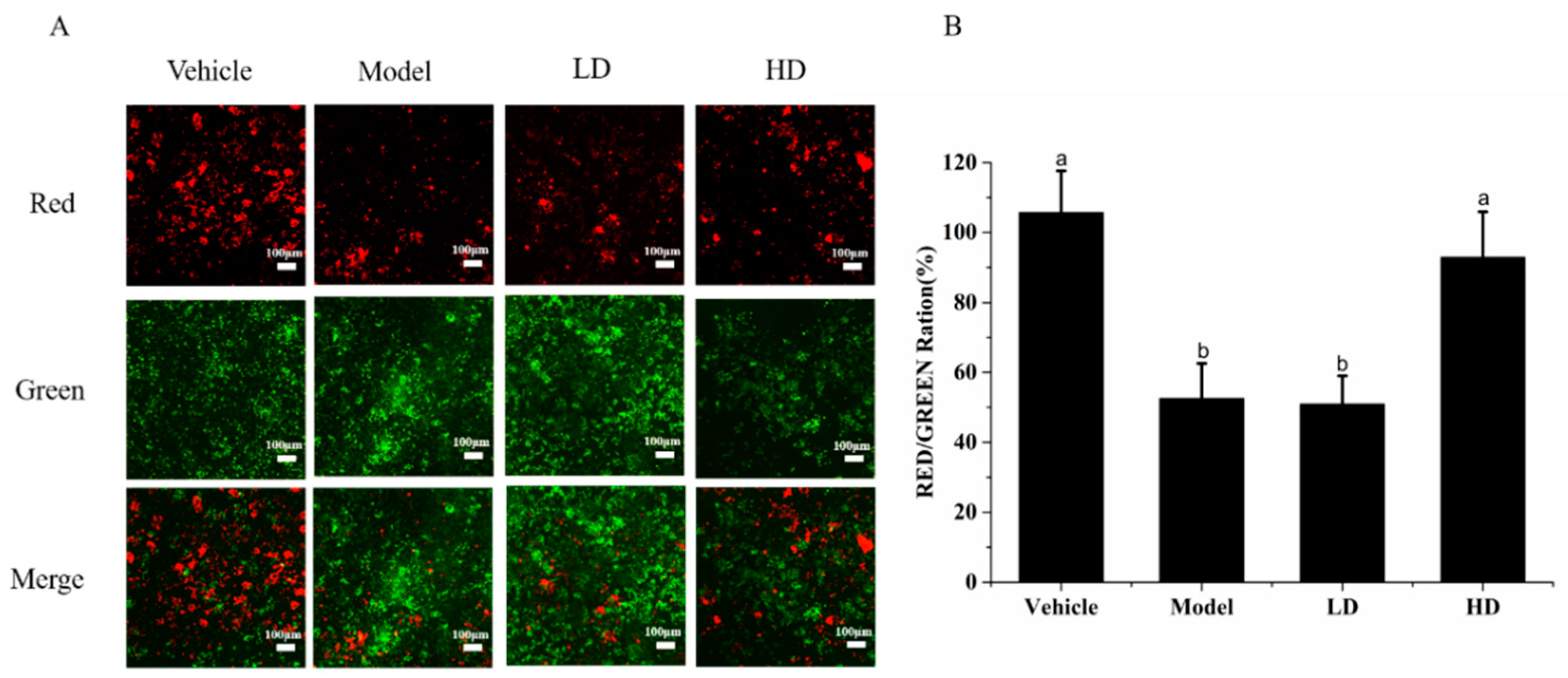
2.5. Effects of SAVEP on LPS-Induced Inflammatory Cell Mitochondrial Membrane Potential
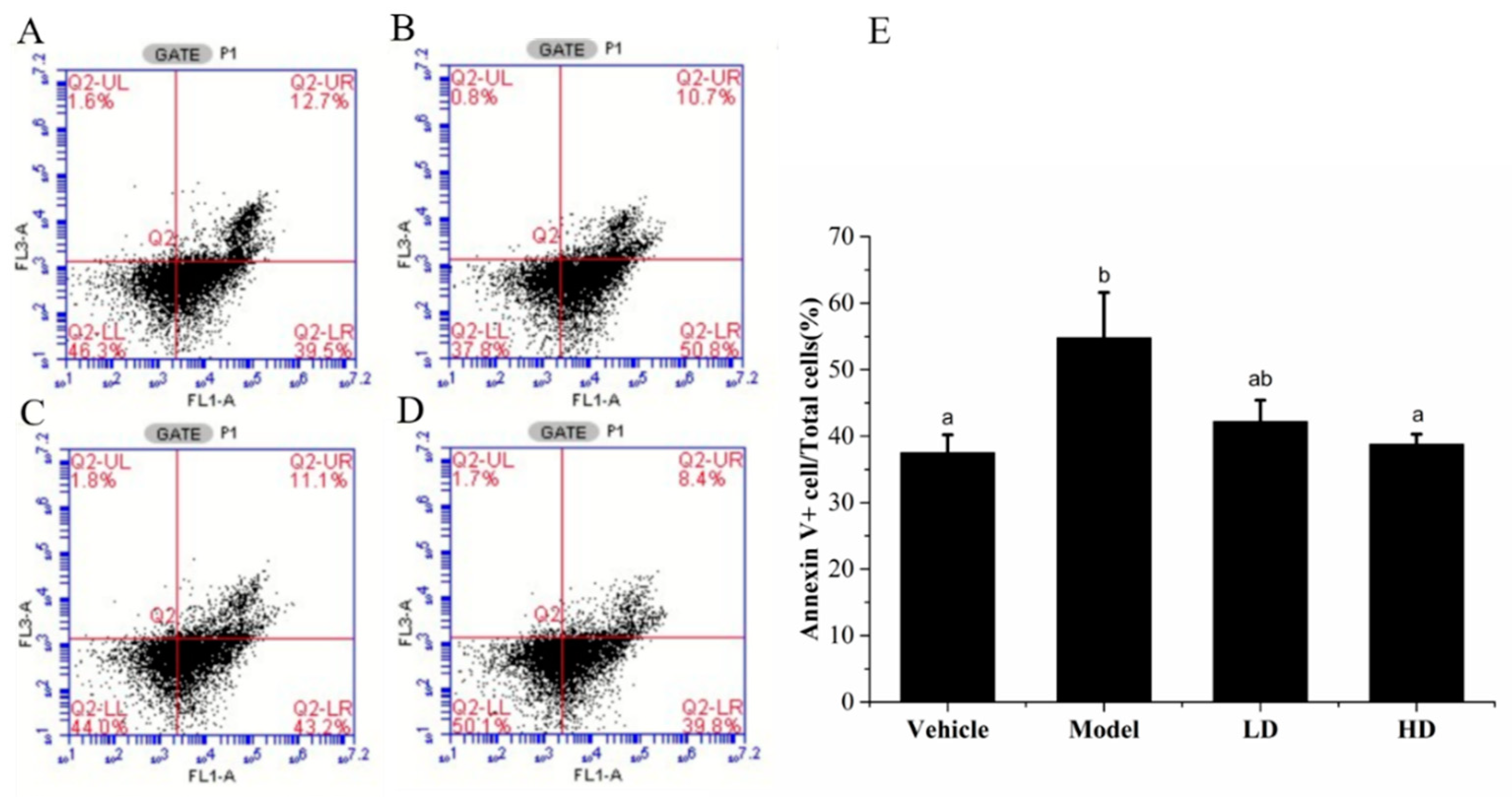
2.6. Effects of SAVEP on LPS-Induced Inflammatory Cell Apoptosis
2.7. Effects of SAVEP on LPS-Induced Inflammatory Cytokine Levels
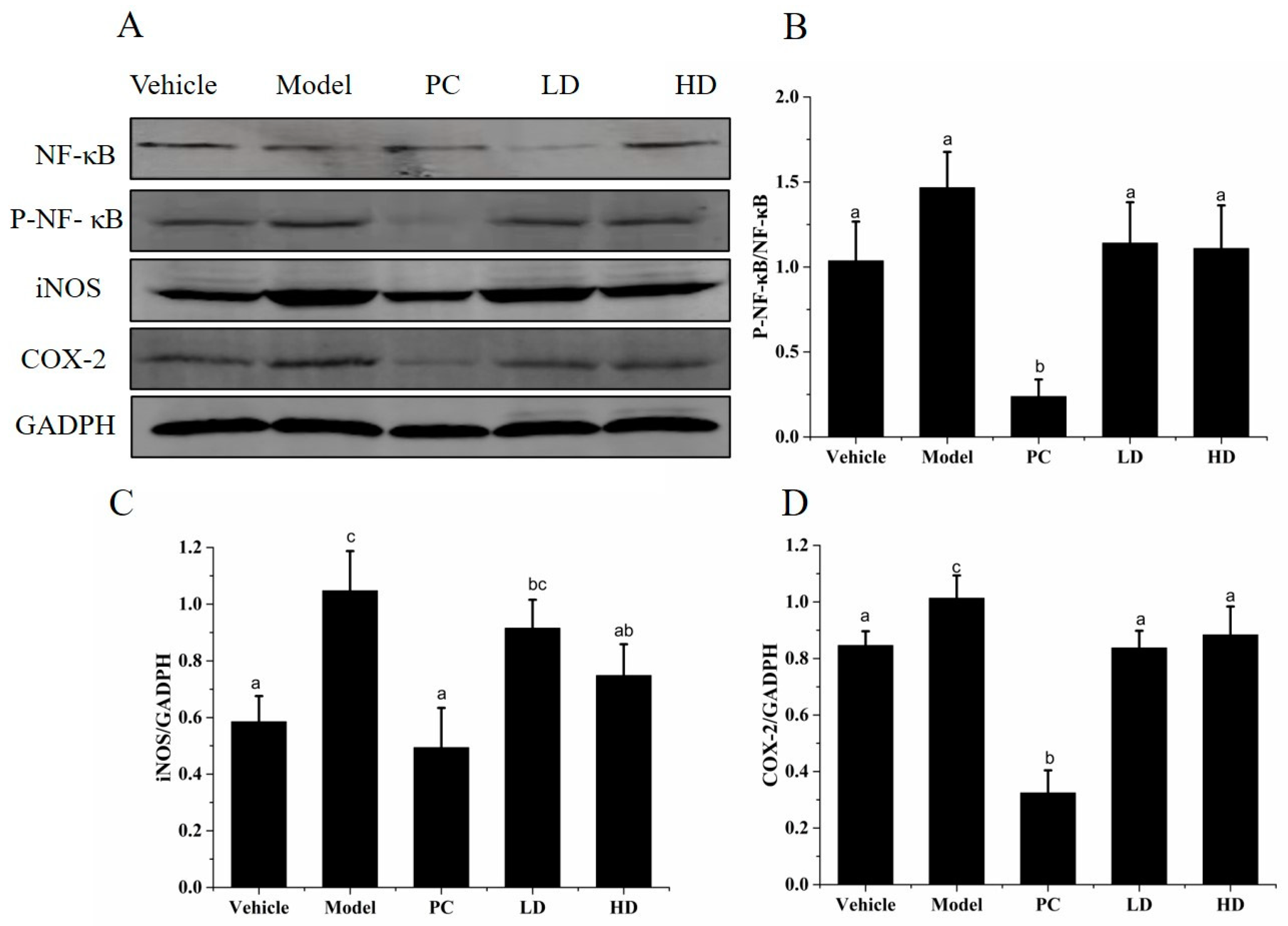
2.8. Effects of SAVEP on LPS-Induced Inflammatory Cell Inflammatory Protein Expression
2.9. Effects of SAVEP on Inflammatory Factor Levels in Mice
2.10. Effects of SAVEP on Inflammatory Protein Expression in LPS-Induced Inflammatory Mouse Liver
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Polyphenol Extraction
4.3. Analysis of Phenols of SAVEP
4.4. Cell Culture and Cell Viability Assay
4.5. Scanning Electron Microscopy (SEM)
4.6. Hoechst 33342/Propidium Iodide (PI) Staining
4.7. JC-1 Staining
4.8. Annexin V-FITC/PI Staining
4.9. Analysis of Cytokine Expression and Mouse Serum Index
4.10. Western Blot Analysis
4.11. Animals, Fodder and Treatment
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chen, H.; Chen, T.; Giudici, P.; Chen, F. Vinegar Functions on Health: Constituents, Sources, and Formation Mechanisms. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1124–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Xia, T.; Du, P.; Duan, W.; Zhang, B.; Zhang, J.; Zhu, S.; Zheng, Y.; Wang, M.; Yu, Y. Chemical Composition and Antioxidant Characteristic of Traditional and Industrial Zhenjiang Aromatic Vinegars during the Aging Process. Molecules 2018, 23, 2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, M.M.; Wang, X.Y.; Tian, C.R.; Li, X.J.; Zhang, B.S.; Song, X.Z.; Zhang, J. Characterization of Organic Acids and Phenolic Compounds of Cereal Vinegars and Fruit Vinegars in China. J. Food Process. Preserv. 2017, 41. [Google Scholar] [CrossRef]
- Samad, A.; Azlan, A.; Ismail, A. Therapeutic effects of vinegar: A review. Curr. Opin. Food Sci. 2016, 8, 56–61. [Google Scholar] [CrossRef]
- Banach, M.; Wiloch, M.; Zawada, K.; Cyplik, W.; Kujawski, W. Evaluation of Antioxidant and Anti-Inflammatory Activity of Anthocyanin-Rich Water-Soluble Aronia Dry Extracts. Molecules 2020, 25, 4055. [Google Scholar] [CrossRef]
- Date, A.A.; Destache, C.J. Natural polyphenols: Potential in the prevention of sexually transmitted viral infections. Drug Discov. Today 2016, 21, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current evidence on the effect of dietary polyphenols intake on chronic diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Giglio, R.V.; Patti, A.M.; Cicero, A.F.G.; Lippi, G.; Rizzo, M.; Toth, P.P.; Banach, M. Polyphenols: Potential Use in the Prevention and Treatment of Cardiovascular Diseases. Curr. Pharm. Des. 2018, 24, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J.; Jung, H.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-κB and MAPK signaling pathways in LPS-induced RAW264.7 macrophage cells. J. Ethnopharmacol. 2019, 231, 10–18. [Google Scholar] [CrossRef]
- Li, C.-L.; Tan, L.-H.; Wang, Y.-F.; Luo, C.-D.; Chen, H.-B.; Lu, Q.; Li, Y.-C.; Yang, X.-B.; Chen, J.-N.; Liu, Y.-H.; et al. Comparison of anti-inflammatory effects of berberine, and its natural oxidative and reduced derivatives from Rhizoma Coptidis in vitro and in vivo. Phytomedicine 2019, 52, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Shimegi, S.; Fischer, Q.S.; Yang, Y.; Sato, H.; Daw, N.W. Blockade of Cyclic AMP-Dependent Protein Kinase Does Not Prevent the Reverse Ocular Dominance Shift in Kitten Visual Cortex. J. Neurophysiol. 2003, 90, 4027–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulakh, R.; Singh, S. Strategies for minimizing corticosteroid toxicity: A review. Indian J. Pediatr 2008, 75, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.-K.; Kang, K.-Y.; Jang, H.-Y.; Hwang, Y.-H.; Hong, S.-G.; Kim, S.-J.; Cho, H.-W.; Chang, D.-J.; Hur, J.-S.; Yee, S.-T. Atraric Acid Exhibits Anti-Inflammatory Effect in Lipopolysaccharide-Stimulated RAW264.7 Cells and Mouse Models. Int. J. Mol. Sci. 2020, 21, 7070. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Zoubouli, P.; Calder, P.C. Effects of polyphenols on human Th1 and Th2 cytokine production. Clin. Nutr. 2005, 24, 780–784. [Google Scholar] [CrossRef]
- Biesalski, H.K. Polyphenols and inflammation: Basic interactions. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Konukoglu, D.; Turhan, M.S.; Ercan, M.; Serin, O. Relationship between plasma leptin and zinc levels and the effect of insulin and oxidative stress on leptin levels in obese diabetic patients. J. Nutr. Biochem. 2004, 15, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Na, H.K.; Chun, K.S.; Kim, Y.K.; Lee, S.J.; Lee, S.S.; Lee, O.S.; Sim, Y.C.; Surh, Y.J. Inhibition of phorbol ester-induced COX-2 expression by epigallocatechin gallate in mouse skin and cultured human mammary epithelial cells. J. Nutr. 2003, 133, 3805S–3810S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doster, M.A.; Michailides, T.J. Production of Bright Greenish Yellow Fluorescence in Figs Infected by Aspergillus Species in California Orchards. Plant Dis. 1998, 82, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santangelo, C.; Varì, R.; Scazzocchio, B.; Di Benedetto, R.; Filesi, C.; Masella, R. Polyphenols, intracellular signalling and inflammation. Annali dell’Istituto Superiore di Sanità 2007, 43, 394–405. [Google Scholar]
- Du, P.; Zhou, J.; Zhang, L.; Zhang, J.; Li, N.; Zhao, C.; Tu, L.; Zheng, Y.; Xia, T.; Luo, J.; et al. GC x GC-MS analysis and hypolipidemic effects of polyphenol extracts from Shanxi-aged vinegar in rats under a high fat diet. Food Funct. 2020, 11, 7468–7480. [Google Scholar] [CrossRef]
- Rabah, S.; Kouachi, K.; Ramos, P.A.B.; Gomes, A.P.; Almeida, A.; Haddadi-Guemghar, H.; Madani, K.; Silvestre, A.J.D.; Santos, S.A.O. Unveiling the bioactivity of Allium triquetrum L. lipophilic fractions: Chemical characterization and in vitro antibacterial activity against methicillin-resistant Staphylococcus aureus. Food Funct. 2020, 11, 5257–5265. [Google Scholar] [CrossRef]
- Zong, L.; Zhang, J.; Dai, L.; Liu, J.; Yang, Y.; Xie, J.; Luo, X. The Anti-Inflammatory Properties of Rhododendron molle Leaf Extract in LPS-Induced RAW264.7. Chem. Biodivers. 2020. [Google Scholar] [CrossRef]
- Zamzami, N.; Hirsch, T.; Dallaporta, B.; Petit, P.X.; Kroemer, G. Mitochondrial implication in accidental and programmed cell death: Apoptosis and necrosis. J. Bioenerg. Biomembr. 1997, 29, 185–193. [Google Scholar] [CrossRef]
- Ferri, K.F.; Kroemer, G. Organelle-specific initiation of cell death pathways. Nat. Cell Biol. 2001, 11, E255–E263. [Google Scholar] [CrossRef]
- Kim, R.; Emi, M.; Tanabe, K. Role of mitochondria as the gardens of cell death. Cancer Chemother. Pharm. 2006, 57, 545–553. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Gao, H.; Yuan, R.; Han, S.; Li, X.X.; Tang, M.; Dong, B.; Li, J.X.; Zhao, L.C.; Feng, J.; et al. Procyanidin A2, a polyphenolic compound, exerts anti-inflammatory and anti-oxidative activity in lipopolysaccharide-stimulated RAW264.7 cells. PLoS ONE 2020, 15, e0237017. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Zhou, J.; Lv, H.; Yu, Q.; Peng, L.; Hua, S. Betulin exhibits anti-inflammatory activity in LPS-stimulated macrophages and endotoxin-shocked mice through an AMPK/AKT/Nrf2-dependent mechanism. Cell Death Dis. 2017, 8, e2798. [Google Scholar] [CrossRef] [Green Version]
- Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.; Tzur, A. JC-1: Alternative excitation wavelengths facilitate mitochondrial membrane potential cytometry. Cell Death Dis. 2012, 3, e430. [Google Scholar] [CrossRef] [Green Version]
- Lizard, G.; Miguet, C.; Besséde, G.; Monier, S.; Gueldry, S.; Neel, D.; Gambert, P. Impairment with various antioxidants of the loss of mitochondrial transmembrane potential and of the cytosolic release of cytochrome c occuring during 7-ketocholesterol-induced apoptosis. Free Radic. Biol. Med. 2000, 28, 743–753. [Google Scholar] [CrossRef]
- Hu, Q.; Yuan, B.; Xiao, H.; Zhao, L.; Wu, X.; Rakariyatham, K.; Zhong, L.; Han, Y.; Kimatu, M.B.; Yang, W. Polyphenols-rich extract from Pleurotus eryngii with growth inhibitory of HCT116 colon cancer cells and anti-inflammatory function in RAW264.7 cells. Food Funct. 2018, 9, 1601–1611. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, W.S.; Shin, J.S.; Jang, D.S.; Lee, K.T. Xanthotoxin suppresses LPS-induced expression of iNOS, COX-2, TNF-α, and IL-6 via AP-1, NF-κB, and JAK-STAT inactivation in RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef]
- Kim, H.-J.; Joe, H.-I.; Zhang, Z.; Lee, W.S.; Lee, K.-Y.; Kook, Y.-B.; An, H.-J. Anti-inflammatory effect of Acalypha australis L. via suppression of NF-κB signaling in LPS-stimulated RAW 264.7 macrophages and LPS-induced septic mice. Mol. Immunol. 2020, 119, 123–131. [Google Scholar] [CrossRef]
- Cho, E.J.; An, H.J.; Shin, J.S.; Choi, H.E.; Ko, J.; Cho, Y.W.; Kim, H.M.; Choi, J.H.; Lee, K.T. Roxatidine suppresses inflammatory responses via inhibition of NF-κB and p38 MAPK activation in LPS-induced RAW 264.7 macrophages. J. Cell. Biochem. 2011, 112, 3648–3659. [Google Scholar] [CrossRef]
- Liu, T.; Xiao, N.; Liu, Y.; Wilson, B.; Li, J.; Wang, Y.; Cao, X.; Liu, Z. The anti-inflammatory effects of ethyl acetate on Lipopolysaccharide/D-galactosamine challenged mice and Lipopolysaccharide activated RAW264.7 cells. Biochem. Biophys. Res. Commun. 2012, 427, 518–524. [Google Scholar] [CrossRef]
- Tang, C.; Sun, J.; Liu, J.; Jin, C.; Wu, X.; Zhang, X.; Chen, H.; Gou, Y.; Kan, J.; Qian, C.; et al. Immune-enhancing effects of polysaccharides from purple sweet potato. Int. J. Biol. Macromol. 2019, 123, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Heeg, A.; Kuchler, L.; Eifler, L.K.; Knape, T.; Heide, H.; Brüne, B.; von Knethen, A. LPS-induced Pellino3 degradation is mediated by p62-dependent autophagy. Crit. Care 2012, 16, P50. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Kitano, H. A comprehensive map of the toll-like receptor signaling network. Mol. Syst. Biol. 2006, 2, 2006.0015. [Google Scholar] [CrossRef]
- An, H.; Xu, H.; Yu, Y.; Zhang, M.; Qi, R.; Yan, X.; Liu, S.; Wang, W.; Guo, Z.; Qin, Z.; et al. Up-regulation of TLR9 gene expression by LPS in mouse macrophages via activation of NF-κB, ERK and p38 MAPK signal pathways. Immunol. Lett. 2002, 81, 165–169. [Google Scholar] [CrossRef]
- Guimaraes, M.R.; Leite, F.R.; Spolidorio, L.C.; Kirkwood, K.L.; Rossa, C., Jr. Curcumin abrogates LPS-induced pro-inflammatory cytokines in RAW 264.7 macrophages. Evidence for novel mechanisms involving SOCS-1, -3 and p38 MAPK. Arch. Oral. Biol. 2013, 58, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Neuder, L.E.; Keener, J.M.; Eckert, R.E.; Trujillo, J.C.; Jones, S.L. Role of p38 MAPK in LPS induced pro-inflammatory cytokine and chemokine gene expression in equine leukocytes. Vet. Immunol. Immunopathol. 2009, 129, 192–199. [Google Scholar] [CrossRef]
- Somensi, N.; Rabelo, T.K.; Guimaraes, A.G.; Quintans-Junior, L.J.; de Souza Araujo, A.A.; Moreira, J.C.F.; Gelain, D.P. Carvacrol suppresses LPS-induced pro-inflammatory activation in RAW 264.7 macrophages through ERK1/2 and NF-kB pathway. Int. Immunopharmacol. 2019, 75, 105743. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Patel, G.; Xue, Q.; Singor Njateng, G.S.; Cai, S.; Cheng, G.; Kai, G. In vitro and in vivo anti-inflammatory effects of different extracts from Epigynum auritum through down-regulation of NF-κB and MAPK signaling pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef]
- Tangney, C.C.; Rasmussen, H.E. Polyphenols, inflammation, and cardiovascular disease. Curr. Atheroscler. Rep. 2013, 15, 324. [Google Scholar] [CrossRef]
- Im, J.Y.; Kim, D.; Lee, K.W.; Kim, J.B.; Lee, J.K.; Kim, D.S.; Lee, Y.I.; Ha, K.S.; Joe, C.O.; Han, P.L. COX-2 Regulates the Insulin-Like Growth Factor I-Induced Potentiation of Zn(2+)-Toxicity in Primary Cortical Culture. Mol. Pharmacol. 2004, 66, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tucsek, Z.; Radnai, B.; Racz, B.; Debreceni, B.; Priber, J.K.; Dolowschiak, T.; Palkovics, T.; Gallyas, F., Jr.; Sumegi, B.; Veres, B. Suppressing LPS-induced early signal transduction in macrophages by a polyphenol degradation product: A critical role of MKP-1. J. Leukoc. Biol. 2011, 89, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.X.; Luo, D.; Tanigawa, S.; Hashimoto, F.; Uto, T.; Masuzaki, S.; Fujii, M.; Sakata, Y. Prodelphinidin B-4 3′-O-gallate, a tea polyphenol, is involved in the inhibition of COX-2 and iNOS via the downregulation of TAK1-NF-κB pathway. Biochem. Pharmacol. 2007, 74, 742–751. [Google Scholar] [CrossRef]
- Hou, D.X.; Masuzaki, S.; Hashimoto, F.; Uto, T.; Tanigawa, S.; Fujii, M.; Sakata, Y. Green tea proanthocyanidins inhibit cyclooxygenase-2 expression in LPS-activated mouse macrophages: Molecular mechanisms and structure-activity relationship. Arch. Biochem. Biophys. 2007, 460, 67–74. [Google Scholar] [CrossRef]
- Karin, M.; Yamamoto, Y.; Wang, Q.M. The IKK NF-κB system: A treasure trove for drug development. Nat. Rev. Drug Discov. 2004, 3, 17–26. [Google Scholar] [CrossRef]
- Lee, T.H.; Tai, D.I.; Cheng, C.J.; Sun, C.S.; Lin, C.Y.; Sheu, M.J.; Lee, W.P.; Peng, C.Y.; Wang, A.H.; Tsai, S.L. Enhanced nuclear factor-kappa B-associated Wnt-1 expression in hepatitis B- and C-related hepatocarcinogenesis: Identification by functional proteomics. J. Biomed. Sci. 2006, 13, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yuan, X.; Chen, L.; Wang, T.; Wang, Z.; Sun, G.; Li, X.; Li, X.; Liu, G. Histamine Induces Bovine Rumen Epithelial Cell Inflammatory Response via NF-κB Pathway. Cell. Physiol. Biochem. 2017, 42, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Lin, J.-K. (−)-Epigallocatechin-3-gallate Blocks the Induction of Nitric Oxide Synthase by Down-Regulating Lipopolysaccharide-Induced Activity of Transcription Factor Nuclear Factor-κB. Mol. Pharmacol. 1997, 52, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Lau, F.C.; Joseph, J.A.; McDonald, J.E.; Kalt, W. Attenuation of iNOS and COX2 by blueberry polyphenols is mediated through the suppression of NF-κB activation. J. Funct. Foods 2009, 1, 274–283. [Google Scholar] [CrossRef]
- Hou, J.; Hu, M.; Zhang, L.; Gao, Y.; Ma, L.; Xu, Q. Dietary Taxifolin Protects Against Dextran Sulfate Sodium-Induced Colitis via NF-κB Signaling, Enhancing Intestinal Barrier and Modulating Gut Microbiota. Front. Immunol. 2020, 11, 631809. [Google Scholar] [CrossRef] [PubMed]
- Famurewa, A.C.; Ejezie, A.J.; Ugwu-Ejezie, C.S.; Ikekpeazu, E.J.; Ejezie, F.E. Antioxidant and anti-inflammatory mechanisms of polyphenols isolated from virgin coconut oil attenuate cadmium-induced oxidative stress-mediated nephrotoxicity and inflammation in rats. J. Appl. Biomed. 2018, 16, 281–288. [Google Scholar] [CrossRef]
- Yeo, S.C.M.; Fenwick, P.S.; Barnes, P.J.; Lin, H.S.; Donnelly, L.E. Isorhapontigenin, a bioavailable dietary polyphenol, suppresses airway epithelial cell inflammation through a corticosteroid-independent mechanism. Br. J. Pharmacol. 2017, 174, 2043–2059. [Google Scholar] [CrossRef]
- Grun, C.H.; van Dorsten, F.A.; Jacobs, D.M.; Le Belleguic, M.; van Velzen, E.J.; Bingham, M.O.; Janssen, H.G.; van Duynhoven, J.P. GC-MS methods for metabolic profiling of microbial fermentation products of dietary polyphenols in human and in vitro intervention studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 871, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Soleha, M.; Pratiwi, D.E.; Sari, I.D.; Hermiyanti, E.; Yunarto, N.; Setyorini, H.A. Antioxidant Activity of Methanol Extract Tetracera scanden L Merr Predicted Active Compound of Methanol Extract with GCMS NIST Library. J. Phys. Conf. Ser. 2020, 1665, 012028. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Lee, H.G.; Kim, H.S.; Vaas, A.; De Silva, H.I.C.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, D.S.; Lee, J.S.; et al. Fucoidan Purified from Sargassum polycystum Induces Apoptosis through Mitochondria-Mediated Pathway in HL-60 and MCF-7 Cells. Mar. Drugs 2020, 18, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamada, T.; Kang, M.C.; Phan, C.S.; Zanil, I.I.; Jeon, Y.J.; Vairappan, C.S. Bioactive Cembranoids from the Soft Coral Genus Sinularia sp. in Borneo. Mar. Drugs 2018, 16, 99. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Du, L.B.; Kumar, A.; Jia, H.Y.; Liang, X.J.; Tian, Q.; Nie, G.J.; Liu, Y. Inhibitory effects of trolox-encapsulated chitosan nanoparticles on tert-butylhydroperoxide induced RAW264.7 apoptosis. Biomaterials 2012, 33, 8517–8528. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.M.; Gauley, J.; Pisetsky, D.S. The properties of microparticles from RAW 264.7 macrophage cells undergoing in vitro activation or apoptosis. Innate Immun. 2014, 20, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Zhang, J.; Su, Y.; Zhang, X.; Li, J.; Tu, L.; Yu, J.; Zheng, Y.; Wang, M. Monascus vinegar-mediated alternation of gut microbiota and its correlation with lipid metabolism and inflammation in hyperlipidemic rats. J. Funct. Foods 2020, 74, 104152. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vehicle | Model | LD | HD | p-Value | |
|---|---|---|---|---|---|
| IL-1β(pg/mL) | 2.31 ± 0.20 a | 4.14 ± 0.64 b | 2.85 ± 0.35 a | 2.53 ± 0.44 a | 0.004 |
| IL-6(pg/mL) | 2.46 ± 0.13 ab | 2.78 ± 0.20 b | 2.51 ± 0.11 ab | 2.26 ± 0.26 a | 0.049 |
| IL-18(pg/mL) | 17.80 ± 0.51 a | 21.73 ± 0.60 b | 20.41 ± 0.49 b | 18.46 ± 1.21 a | 0.008 |
| MCP-1(pg/mL) | 2.21 ± 0.20 a | 3.06 ± 0.20 b | 2.87 ± 0.35 b | 2.75 ± 0.25 ab | 0.019 |
| NO(μmol/L) | 4.60 × 10−3 ± 2.00 × 10−4 a | 5.30 × 10−3 ± 1.34 × 10−4 b | 5.39 × 10−3 ± 5.06 × 10−4 b | 5.20 × 10−3 ± 1.38 × 10−4 ab | 0.033 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, P.; Song, J.; Qiu, H.; Liu, H.; Zhang, L.; Zhou, J.; Jiang, S.; Liu, J.; Zheng, Y.; Wang, M. Polyphenols Extracted from Shanxi-Aged Vinegar Inhibit Inflammation in LPS-Induced RAW264.7 Macrophages and ICR Mice via the Suppression of MAPK/NF-κB Pathway Activation. Molecules 2021, 26, 2745. https://doi.org/10.3390/molecules26092745
Du P, Song J, Qiu H, Liu H, Zhang L, Zhou J, Jiang S, Liu J, Zheng Y, Wang M. Polyphenols Extracted from Shanxi-Aged Vinegar Inhibit Inflammation in LPS-Induced RAW264.7 Macrophages and ICR Mice via the Suppression of MAPK/NF-κB Pathway Activation. Molecules. 2021; 26(9):2745. https://doi.org/10.3390/molecules26092745
Chicago/Turabian StyleDu, Peng, Jia Song, Huirui Qiu, Haorui Liu, Li Zhang, Junhan Zhou, Shengping Jiang, Jinyu Liu, Yu Zheng, and Min Wang. 2021. "Polyphenols Extracted from Shanxi-Aged Vinegar Inhibit Inflammation in LPS-Induced RAW264.7 Macrophages and ICR Mice via the Suppression of MAPK/NF-κB Pathway Activation" Molecules 26, no. 9: 2745. https://doi.org/10.3390/molecules26092745