
Counteracting the Ramifications of UVB Irradiation and Photoaging with Swietenia macrophylla King Seed

 ,
,  , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cytotoxicity Assessment of S. macrophylla Extract and Fractions
2.2. The Dynamic Proteomics and Genomic Dysregulation in Keratinocyte Cells Effectuated by UVB and Its Attenuation by S. macrophylla
2.2.1. Analysis of UVB-Induced Protein Modifications and the Reversal Effect of S. macrophylla
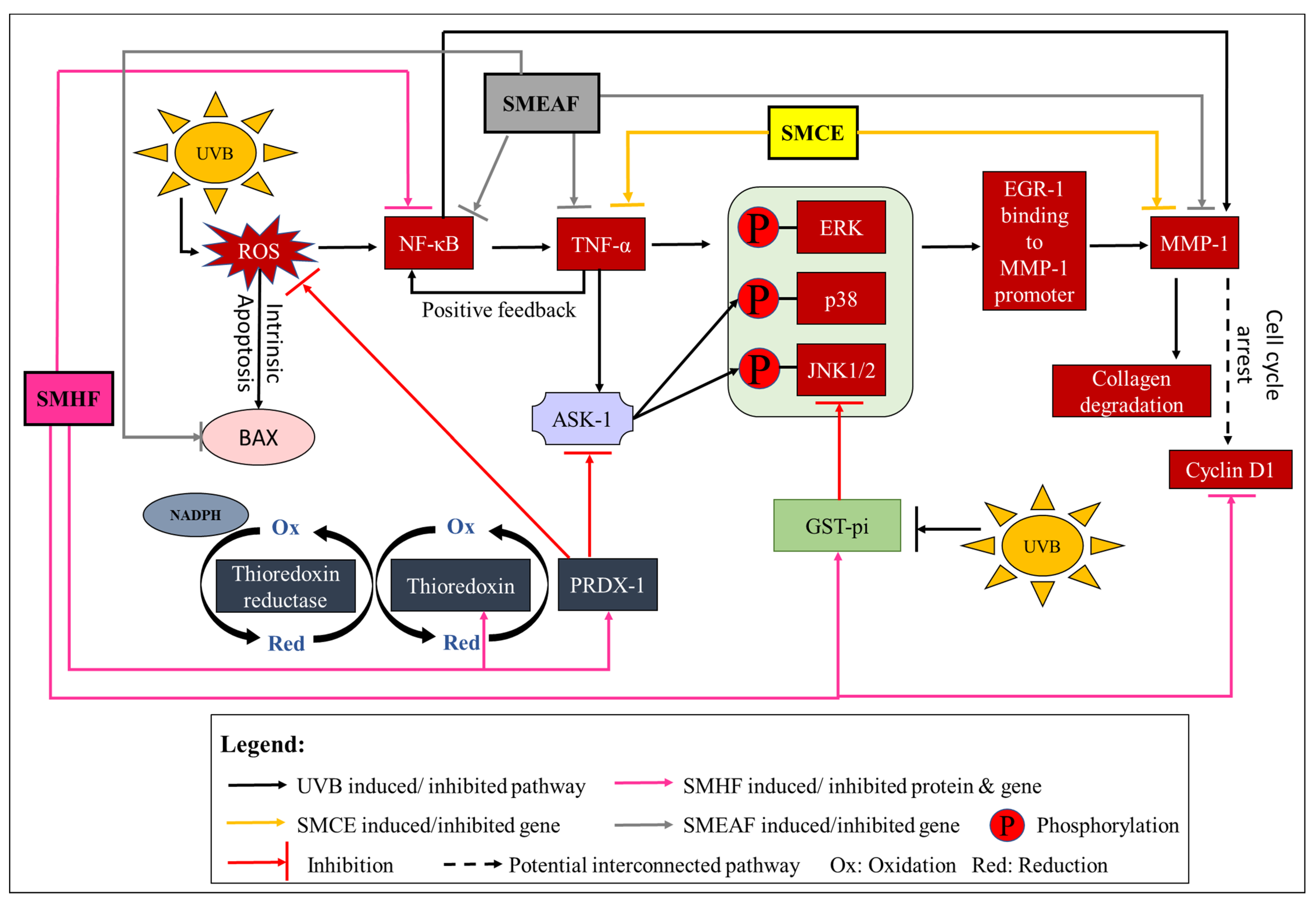
2.2.2. Oxidative Damage Induced Activation of the Redox System
2.2.3. Impact of UVB on DNA Maintenance and Repair in Keratinocyte Cells
2.2.4. Modifications of Downstream Process from RNA to Protein
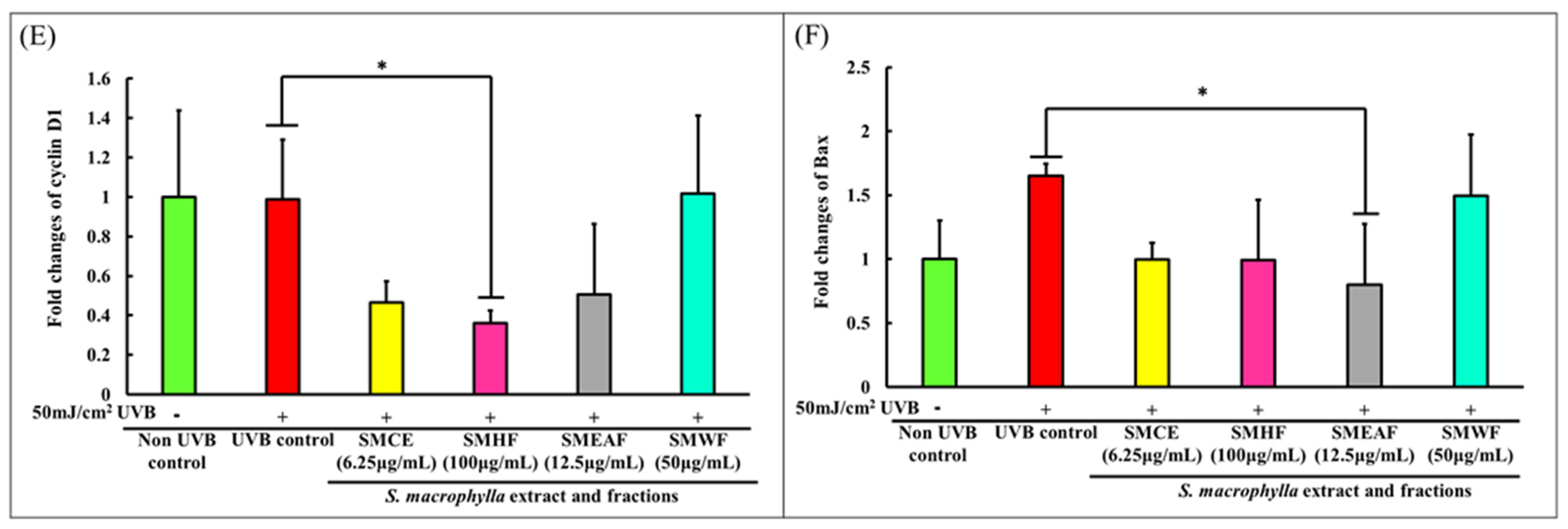
2.2.5. UVB Exposure Affects Cell Growth, Proliferation, and Migration
2.2.6. SMHF Upregulates Proteins Involved in Cell Glycolysis
2.2.7. Dysregulation of Gene Expression in HaCaT Cells after UVB Irradiation
3. Materials and Methods
3.1. Plant Material and Extraction
3.2. Cell Line and Maintenance
3.3. Cytotoxicity Assay
3.4. UV Irradiation
3.5. Protein Expression Studies
3.5.1. Preparation of Lysis Buffer
3.5.2. Protein Extraction
3.5.3. Bicinchoninic Acid (BCA) Protein Assay
3.5.4. In-Solution Tryptic Digestion
3.5.5. Protein Sample Desalting and Cleanup
3.5.6. Analysis of Protein Samples with Nanoflow-Ultra High-Performance Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
3.5.7. Protein Identification and Differential Expression Studies with PEAKS Bioinformatics Software
3.6. Quantitative Polymerase Chain Reaction (PCR) Analysis of Gene Expression Changes in UVB Irradiated Cells
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bogh, M.K.B.; Schmedes, A.V.; Philipsen, P.A.; Thieden, E.; Wulf, H.C. Vitamin D production after UVB exposure depends on baseline vitamin D and total cholesterol but not on skin pigmentation. J. Investig. Dermatol. 2010, 130, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Gathers, R.C.; Scherschun, L.; Malick, F.; Fivenson, D.P.; Lim, H.W. Narrowband UVB phototherapy for early-stage mycosis fungoides. J. Am. Acad. Dermatol. 2002, 47, 191–197. [Google Scholar] [CrossRef]
- Kanwar, A.J.; Dogra, S.; Parsad, D.; Kumar, B. Narrow-band UVB for the treatment of vitiligo: An emerging effective and well-tolerated therapy. Int. J. Dermatol. 2005, 44, 57–60. [Google Scholar] [CrossRef]
- Kirke, S.M.; Lowder, S.; Lloyd, J.J.; Diffey, B.L.; Matthews, J.N.S.; Farr, P.M. A randomized comparison of selective broadband UVB and narrowband UVB in the treatment of psoriasis. J. Investig. Dermatol. 2007, 127, 1641–1646. [Google Scholar] [CrossRef] [Green Version]
- Scharffetter–Kochanek, K.; Brenneisen, P.; Wenk, J.; Herrmann, G.; Ma, W.; Kuhr, L.; Meewes, C.; Wlaschek, M. Photoaging of the skin from phenotype to mechanisms. Exp. Gerontol. 2000, 35, 307–316. [Google Scholar] [CrossRef]
- Guinot, C.; Malvy, D.J.M.; Ambroisine, L.; Latreille, J.; Mauger, E.; Tenenhaus, M.; Morizot, F.; Lopez, S.; Le Fur, I.; Tschachler, E. Relative contribution of intrinsic vs extrinsic factors to skin aging as determined by a validated skin age score. Arch. Dermatol. 2002, 138, 1454–1460. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. DermatoEndocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addor, F.A.S.A. Beyond photoaging: Additional factors involved in the process of skin aging. Clin. Cosmet. Investig. Dermatol. 2018, 11, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Mahendra, C.K.; Tan, L.T.H.; Pusparajah, P.; Htar, T.T.; Chuah, L.-H.; Lee, V.S.; Low, L.E.; Tang, S.Y.; Chan, K.-G.; Goh, B.H. Detrimental effects of UVB on retinal pigment epithelial cells and its role in age-related macular degeneration. Oxid. Med. Cell. Longev. 2020, 2020, 1904178. [Google Scholar] [CrossRef] [PubMed]
- Berkey, C.; Biniek, K.; Dauskardt, R.H. Screening sunscreens: Protecting the biomechanical barrier function of skin from solar ultraviolet radiation damage. Int. J. Cosmet. Sci. 2017, 39, 269–274. [Google Scholar] [CrossRef] [PubMed]
- You, Y.-H.; Lee, D.-H.; Jung-Hoon, Y.; Nkajima, S.; Yasui, A.; Pfeifer, G.P. Cyclobutane pyrimidine dimers are responsible for the vast majority of mutations induced by UVB irradiation in mammalian cells. J. Biol. Chem. 2001, 276, 44688–44694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, D.; Raffa, S.; Flori, E.; Aspite, N.; Briganti, S.; Cardinali, G.; Torrisi, M.R.; Picardo, M. Keratinocyte growth factor down-regulates intracellular ROS production induced by UVB. J. Dermatol. Sci. 2009, 54, 106–113. [Google Scholar] [CrossRef]
- Filip, G.A.; Postescu, I.D.; Bolfa, P.; Catoi, C.; Muresan, A.; Clichici, S. Inhibition of UVB-induced skin phototoxicity by a grape seed extract as modulator of nitrosative stress, ERK/NF-kB signaling pathway and apoptosis, in SKH-1 mice. Food Chem. Toxicol. 2013, 57, 296–306. [Google Scholar] [CrossRef]
- Bashir, M.M.; Sharma, M.R.; Werth, V.P. UVB and pro-inflammatory cytokines synergistically activate TNF-α production in keratinocytes through enhanced gene transcription. J. Investig. Dermatol. 2009, 129, 994–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagot, D.; Asselineau, D.; Bernerd, F. Direct role of human dermal fibroblasts and indirect participation of epidermal keratinocytes in MMP-1 production after UV-B irradiation. Arch. Dermatol. Res. 2002, 293, 576–583. [Google Scholar] [CrossRef]
- Niwano, T.; Terazawa, S.; Nakajima, H.; Wakabayashi, Y.; Imokawa, G. Astaxanthin and withaferin A block paracrine cytokine interactions between UVB-exposed human keratinocytes and human melanocytes via the attenuation of endothelin-1 secretion and its downstream intracellular signaling. Cytokine 2015, 73, 184–197. [Google Scholar] [CrossRef]
- Saito, M.; Tanaka, M.; Misawa, E.; Yao, R.; Nabeshima, K.; Yamauchi, K.; Abe, F.; Yamamoto, Y.; Furukawa, F. Oral administration of Aloe vera gel powder prevents UVB-induced decrease in skin elasticity via suppression of overexpression of MMPs in hairless mice. Biosci. Biotechnol. Biochem. 2016, 80, 1416–1424. [Google Scholar] [CrossRef] [Green Version]
- Ewles, M.; Goodwin, L. Bioanalytical approaches to analyzing peptides and proteins by LC–MS/MS. Bioanalysis 2011, 3, 1379–1397. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.H.; Abdul Kadir, H.; Abdul Malek, S.N.; Ng, S.W. (αR,4R,4aR,6aS,7R,8S,10R,11S)-Methyl α-acet-oxy-4-(3-furan-yl)-10-hy-droxy-4a,7,9,9-tetra-methyl-2,13-dioxo-1,4,4a,5,6,6a,7,8,9,10,11,12-dodeca-hydro-7,11-methano-2H-cyclo-octa-[f][2]benzopyran-8-acetate (6-O-acetyl-swietenolide) from the seeds of Swietenia macrophylla. Acta Crystallogr. Sect. E Struct. Rep. Online 2010, 66, o2802–o2803. [Google Scholar]
- Wu, S.F.; Lin, C.K.; Chuang, Y.S.; Chang, F.R.; Tseng, C.K.; Wu, Y.C.; Lee, J.C. Anti--hepatitis C virus activity of 3--hydroxy caruilignan C from Swietenia macrophylla stems. J. Viral Hepat. 2012, 19, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-B.; Chien, Y.-T.; Lee, J.-C.; Tseng, C.-K.; Wang, H.-C.; Lo, I.-W.; Wu, Y.-H.; Wang, S.-Y.; Wu, Y.-C.; Chang, F.-R. Limonoids from the seeds of Swietenia macrophylla with inhibitory activity against dengue virus 2. J. Nat. Prod. 2014, 77, 2367–2374. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.H.; Abdul Kadir, H.; Abdul Malek, S.N.; Ng, S.W. Swietenolide diacetate from the seeds of Swietenia macrophylla. Acta Crystallogr. Sect. E Struct. Rep. Online 2010, 66, o1396. [Google Scholar] [CrossRef] [Green Version]
- Goh, B.H.; Abdul Kadir, H. In vitro cytotoxic potential of Swietenia macrophylla King seeds against human carcinoma cell lines. J. Med. Plant Res. 2011, 5, 1395–1404. [Google Scholar]
- Hashim, M.A.; Yam, M.F.; Hor, S.Y.; Lim, C.P.; Asmawi, M.Z.; Sadikun, A. Anti-hyperglycaemic activity of Swietenia macrophylla King (Meliaceae) seed extracts in normoglycaemic rats undergoing glucose tolerance tests. Chin. Med. 2013, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadamtousi, S.Z.; Goh, B.H.; Chan, C.K.; Shabab, T.; Kadir, H.A. Biological activities and phytochemicals of Swietenia macrophylla king. Molecules 2013, 18, 10465–10483. [Google Scholar] [CrossRef] [PubMed]
- Sayyad, M.; Tiang, N.; Kumari, Y.; Goh, B.H.; Jaiswal, Y.; Rosli, R.; Williams, L.; Shaikh, M.F. Acute toxicity profiling of the ethyl acetate fraction of Swietenia macrophylla seeds and in-vitro neuroprotection studies. Saudi Pharm. J. 2017, 25, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Supriady, H.; Kamarudin, M.N.A.; Chan, C.K.; Goh, B.H.; Kadir, H.A. SMEAF attenuates the production of pro-inflammatory mediators through the inactivation of Akt-dependent NF-κB, p38 and ERK1/2 pathways in LPS-stimulated BV-2 microglial cells. J. Funct. Foods 2015, 17, 434–448. [Google Scholar] [CrossRef]
- Mak, K.-K.; Shiming, Z.; Balijepalli, M.K.; Dinkova-Kostova, A.T.; Epemolu, O.; Mohd, Z.; Pichika, M.R. Studies on the mechanism of anti-inflammatory action of swietenine, a tetranortriterpenoid isolated from Swietenia macrophylla seeds. Phytomedicine Plus 2021, 1, 100018. [Google Scholar] [CrossRef]
- Nilugal, K.C.; Fattepur, S.; Asmani, M.F.; Abdullah, I.; Vijendren, S.; Ugandar, R.E. Evaluation of wound healing acitivity of Swietenia macrophylla (Meliaceae) seed extract in albino rats. Am. J. PharmTech Res. 2017, 7, 113–124. [Google Scholar]
- Wang, X.; Phelan, S.A.; Forsman-Semb, K.; Taylor, E.F.; Petros, C.; Brown, A.; Lerner, C.P.; Paigen, B. Mice with targeted mutation of peroxiredoxin 6 develop normally but are susceptible to oxidative stress. J. Biol. Chem. 2003, 278, 25179–25190. [Google Scholar] [CrossRef] [Green Version]
- Fujii, J.; Ikeda, Y. Advances in our understanding of peroxiredoxin, a multifunctional, mammalian redox protein. Redox Rep. 2002, 7, 123–130. [Google Scholar] [CrossRef]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, glutaredoxins, and peroxiredoxins--molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef] [PubMed]
- Salzano, S.; Checconi, P.; Hanschmann, E.-M.; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S.; et al. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc. Natl. Acad. Sci. USA 2014, 111, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Day, A.M.; Brown, J.D.; Taylor, S.R.; Rand, J.D.; Morgan, B.A.; Veal, E.A. Inactivation of a peroxiredoxin by hydrogen peroxide is critical for thioredoxin-mediated repair of oxidized proteins and cell survival. Mol. Cell 2012, 45, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Netto, L.E.S.; Antunes, F. The roles of peroxiredoxin and thioredoxin in hydrogen peroxide sensing and in signal transduction. Mol. Cells 2016, 39, 65–71. [Google Scholar]
- Liu, S.; Guo, C.; Wu, D.; Ren, Y.; Sun, M.-Z.; Xu, P. Protein indicators for HaCaT cell damage induced by UVB irradiation. J. Photochem. Photobiol. B 2012, 114, 94–101. [Google Scholar] [CrossRef]
- Wu, C.-L.; Chou, H.-C.; Cheng, C.-S.; Li, J.-M.; Lin, S.-T.; Chen, Y.-W.; Chan, H.-L. Proteomic analysis of UVB-induced protein expression- and redox-dependent changes in skin fibroblasts using lysine- and cysteine-labeling two-dimensional difference gel electrophoresis. J. Proteomics 2012, 75, 1991–2014. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.Y.; Park, S.; Park, J.; Jang, J.Y.; Wang, S.B.; Kim, S.R.; Woo, H.A.; Lim, K.M.; Chang, T.-S. Protective role of mitochondrial peroxiredoxin III against UVB-induced apoptosis of epidermal keratinocytes. J. Investig. Dermatol. 2017, 137, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, T.J.; Lee, K.-Y. A novel function of peroxiredoxin 1 (Prx-1) in apoptosis signal-regulating kinase 1 (ASK1)-mediated signaling pathway. FEBS Lett. 2008, 582, 1913–1918. [Google Scholar] [CrossRef] [Green Version]
- Adler, V.; Yin, Z.; Fuchs, S.Y.; Benezra, M.; Rosario, L.; Tew, K.D.; Pincus, M.R.; Sardana, M.; Henderson, C.J.; Wolf, C.R.; et al. Regulation of JNK signaling by GSTp. EMBO J. 1999, 18, 1321–1334. [Google Scholar] [CrossRef]
- Verdone, L.; Agricola, E.; Caserta, M.; Di Mauro, E. Histone acetylation in gene regulation. Brief. Funct. Genomic. Protemic. 2006, 5, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Sesto, A.; Navarro, M.; Burslem, F.; Jorcano, J.L. Analysis of the ultraviolet B response in primary human keratinocytes using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 2002, 99, 2965–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dazard, J.-E.; Gal, H.; Amariglio, N.; Rechavi, G.; Domany, E.; Givol, D. Genome-wide comparison of human keratinocyte and squamous cell carcinoma responses to UVB irradiation: Implications for skin and epithelial cancer. Oncogene 2003, 22, 2993–3006. [Google Scholar] [CrossRef] [Green Version]
- Zong, W.-X. Histone 1.2, another housekeeping protein that kills. Cancer Biol. Ther. 2004, 3, 42–43. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Vela, A.; Korsmeyer, S.J. Proapoptotic histone H1.2 induces CASP-3 and -7 activation by forming a protein complex with CYT c, APAF-1 and CASP-9. FEBS Lett. 2007, 581, 3422–3428. [Google Scholar] [CrossRef] [Green Version]
- Colombo, E.; Bonetti, P.; Lazzerini Denchi, E.; Martinelli, P.; Zamponi, R.; Marine, J.-C.; Helin, K.; Falini, B.; Pelicci, P.G. Nucleophosmin is required for DNA integrity and p19Arf protein stability. Mol. Cell. Biol. 2005, 25, 8874–8886. [Google Scholar] [CrossRef] [Green Version]
- Maiguel, D.A.; Jones, L.; Chakravarty, D.; Yang, C.; Carrier, F. Nucleophosmin sets a threshold for p53 response to UV radiation. Mol. Cell. Biol. 2004, 24, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.H.; Chang, J.H.; Yung, B.Y.M. Resistance to UV-induced cell-killing in nucleophosmin/B23 over-expressed NIH 3T3 fibroblasts: Enhancement of DNA repair and up-regulation of PCNA in association with nucleophosmin/B23 over-expression. Carcinogenesis 2002, 23, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamore, S.D.; Qiao, S.; Horn, D.; Wondrak, G.T. Proteomic identification of cathepsin B and nucleophosmin as novel UVA-targets in human skin fibroblasts. Photochem. Photobiol. 2010, 86, 1307–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosammaparast, N.; Ewart, C.S.; Pemberton, L.F. A role for nucleosome assembly protein 1 in the nuclear transport of histones H2A and H2B. EMBO J. 2002, 21, 6527–6538. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Chodaparambil, J.V.; Bao, Y.; McBryant, S.J.; Luger, K. Nucleosome assembly protein 1 exchanges histone H2A-H2B dimers and assists nucleosome sliding. J. Biol. Chem. 2005, 280, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, H.; Sakai, H. A porcine brain protein (35 K protein) which bundles microtubules and its identification as glyceraldehyde 3-phosphate dehydrogenase. J. Biochem. 1983, 93, 1259–1269. [Google Scholar] [CrossRef]
- Singh, R.; Green, M.R. Sequence-specific binding of transfer RNA by glyceraldehyde-3-phosphate dehydrogenase. Science 1993, 259, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, E.J. Glyceraldehyde-3-phosphate dehydrogenase is required for vesicular transport in the early secretory pathway. J. Biol. Chem. 2001, 276, 2480–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.R.; Agrawal, N.; Kim, S.F.; Cascio, M.B.; Fujimuro, M.; Ozeki, Y.; Takahashi, M.; Cheah, J.H.; Tankou, S.K.; Hester, L.D.; et al. S-nitrosylated GAPDH initiates apoptotic cell death by nuclear translocation following Siah1 binding. Nat. Cell Biol. 2005, 7, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Jouvet, N.; Jilani, A.; Vongsamphanh, R.; Yang, X.; Yang, S.; Ramotar, D. Human glyceraldehyde-3-phosphate dehydrogenase plays a direct role in reactivating oxidized forms of the DNA repair enzyme APE1. J. Biol. Chem. 2008, 283, 30632–30641. [Google Scholar] [CrossRef] [Green Version]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell. Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susorov, D.; Zakharov, N.; Shuvalova, E.; Ivanov, A.; Egorova, T.; Shuvalov, A.; Shatsky, I.N.; Alkalaeva, E. Eukaryotic translation elongation factor 2 (eEF2) catalyzes reverse translocation of the eukaryotic ribosome. J. Biol. Chem. 2018, 293, 5220–5229. [Google Scholar] [CrossRef] [Green Version]
- Plafker, S.M.; Macara, I.G. Ribosomal protein L12 uses a distinct nuclear import pathway mediated by importin 11. Mol. Cell. Biol. 2002, 22, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Takao, J.; Ariizumi, K.; Dougherty, I.I.; Cruz, P.D., Jr. Genomic scale analysis of the human keratinocyte response to broad-band ultraviolet-B irradiation. Photodermatol. Photoimmunol. Photomed. 2002, 18, 5–13. [Google Scholar] [CrossRef]
- Kajita, Y.; Nakayama, J.; Aizawa, M.; Ishikawa, F. The UUAG-specific RNA binding protein, heterogeneous nuclear ribonucleoprotein D0. Common modular structure and binding properties of the 2xRBD-Gly family. J. Biol. Chem. 1995, 270, 22167–22175. [Google Scholar] [CrossRef] [Green Version]
- Michelotti, E.F.; Michelotti, G.A.; Aronsohn, A.I.; Levens, D. Heterogeneous nuclear ribonucleoprotein K is a transcription factor. Mol. Cell. Biol. 1996, 16, 2350–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Jia, R. Splicing factor poly(rC)-binding protein 1 is a novel and distinctive tumor suppressor. J. Cell. Physiol. 2018, 234, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Farrukh, M.R.; Nissar, U.A.; Afnan, Q.; Rafiq, R.A.; Sharma, L.; Amin, S.; Kaiser, P.; Sharma, P.R.; Tasduq, S.A. Oxidative stress mediated Ca2+ release manifests endoplasmic reticulum stress leading to unfolded protein response in UV-B irradiated human skin cells. J. Dermatol. Sci. 2014, 75, 24–35. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L. Induction of heat shock proteins for protection against oxidative stress. Ad. Drug Deliv. Rev. 2009, 61, 310–318. [Google Scholar] [CrossRef]
- Edkins, A.L.; Price, J.T.; Pockley, A.G.; Blatch, G.L. Heat shock proteins as modulators and therapeutic targets of chronic disease: An integrated perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160521. [Google Scholar] [CrossRef] [PubMed]
- Jee, H. Size dependent classification of heat shock proteins: A mini-review. J. Exerc. Rehabil. 2016, 12, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Beere, H.M. Death versus survival: Functional interaction between the apoptotic and stress-inducible heat shock protein pathways. J. Clin. Investig. 2005, 115, 2633–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merwald, H.; Kokesch, C.; Klosner, G.; Matsui, M.; Trautinger, F. Induction of the 72-kilodalton heat shock protein and protection from ultraviolet B-induced cell death in human keratinocytes by repetitive exposure to heat shock or 15-deoxy-delta(12,14)-prostaglandin J2. Cell Stress Chaperones 2006, 11, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perluigi, M.; Di Domenico, F.; Blarzino, C.; Foppoli, C.; Cini, C.; Giorgi, A.; Grillo, C.; De Marco, F.; Butterfield, D.A.; Schininà, M.E.; et al. Effects of UVB-induced oxidative stress on protein expression and specific protein oxidation in normal human epithelial keratinocytes: A proteomic approach. Proteome Sci. 2010, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, B.G.; Wang, B.; Freed, I.; Mamelak, A.J.; Watanabe, H.; Sauder, D.N. Microarray analysis of UVB-regulated genes in keratinocytes: Downregulation of angiogenesis inhibitor thrombospondin-1. J. Dermatol. Sci. 2004, 34, 185–194. [Google Scholar] [CrossRef]
- Shan, Y.-X.; Liu, T.-J.; Su, H.-F.; Samsamshariat, A.; Mestril, R.; Wang, P.H. Hsp10 and Hsp60 modulate Bcl-2 family and mitochondria apoptosis signaling induced by doxorubicin in cardiac muscle cells. J. Mol. Cell. Cardiol. 2003, 35, 1135–1143. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Nishida, E.; Koyasu, S.; Sakai, H.; Yahara, I. Calmodulin-regulated binding of the 90-kDa heat shock protein to actin filaments. J. Biol. Chem. 1986, 261, 16033–16036. [Google Scholar] [CrossRef]
- Taiyab, A.; Rao, C.M. HSP90 modulates actin dynamics: Inhibition of HSP90 leads to decreased cell motility and impairs invasion. Biochim. Biophys. Acta 2011, 1813, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Y.; Guan, S.; Fan, J.; Cheng, C.-F.; Bright, A.M.; Chinn, C.; Chen, M.; Woodley, D.T. Extracellular heat shock protein-90α: Linking hypoxia to skin cell motility and wound healing. EMBO J. 2007, 26, 1221–1233. [Google Scholar] [CrossRef]
- Wilkinson, B.; Gilbert, H.F. Protein disulfide isomerase. Biochim. Biophys. Acta 2004, 1699, 35–44. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Wang, C.-C. Protein disulfide–isomerase, a folding catalyst and a redox-regulated chaperone. Free Radic. Biol. Med. 2015, 83, 305–313. [Google Scholar] [CrossRef]
- Kuo, T.-F.; Chen, T.-Y.; Jiang, S.-T.; Chen, K.-W.; Chiang, Y.-M.; Hsu, Y.-J.; Liu, Y.-J.; Chen, H.-M.; Yokoyama, K.K.; Tsai, K.-C.; et al. Protein disulfide isomerase a4 acts as a novel regulator of cancer growth through the procaspase pathway. Oncogene 2017, 36, 5484–5496. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Hu, C.-T.; Cheng, C.-C.; Lee, M.-C.; Pan, S.-M.; Lin, T.-Y.; Wu, W.-S. Oxidation of heat shock protein 60 and protein disulfide isomerase activates ERK and migration of human hepatocellular carcinoma HepG2. Oncotarget 2016, 7, 11067–11082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Synergistic cytoprotective effects of rutin and ascorbic acid on the proteomic profile of 3D-cultured keratinocytes exposed to UVA or UVB radiation. Nutrients 2019, 11, 2672. [Google Scholar] [CrossRef] [Green Version]
- Yokota, S.; Yanagi, H.; Yura, T.; Kubota, H. Cytosolic chaperonin-containing t-complex polypeptide 1 changes the content of a particular subunit species concomitant with substrate binding and folding activities during the cell cycle. Eur. J. Biochem. 2001, 268, 4664–4673. [Google Scholar] [CrossRef]
- Vallin, J.; Grantham, J. The role of the molecular chaperone CCT in protein folding and mediation of cytoskeleton-associated processes: Implications for cancer cell biology. Cell Stress Chaperones 2019, 24, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balcer, H.I.; Goodman, A.L.; Rodal, A.A.; Smith, E.; Kugler, J.; Heuser, J.E.; Goode, B.L. Coordinated regulation of actin filament turnover by a high-molecular-weight Srv2/CAP complex, cofilin, profilin, and Aip1. Curr. Biol. 2003, 13, 2159–2169. [Google Scholar] [CrossRef]
- Zhang, H.; Ghai, P.; Wu, H.; Wang, C.; Field, J.; Zhou, G.-L. Mammalian adenylyl cyclase-associated protein 1 (CAP1) regulates cofilin function, the actin cytoskeleton, and cell adhesion. J. Biol. Chem. 2013, 288, 20966–20977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z.; Lambrechts, A.; Parepally, M.; Roy, P. Silencing profilin-1 inhibits endothelial cell proliferation, migration and cord morphogenesis. J. Cell. Sci. 2006, 119, 4127–4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.P.; Ryu, J.M.; Kim, M.O.; Park, J.H.; Han, H.J. Rapid actions of plasma membrane estrogen receptors regulate motility of mouse embryonic stem cells through a profilin-1/cofilin-1-directed kinase signaling pathway. Mol. Endocrinol. 2012, 26, 1291–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vié, N.; Copois, V.; Bascoul-Mollevi, C.; Denis, V.; Bec, N.; Robert, B.; Fraslon, C.; Conseiller, E.; Molina, F.; Larroque, C.; et al. Overexpression of phosphoserine aminotransferase PSAT1 stimulates cell growth and increases chemoresistance of colon cancer cells. Mol. Cancer 2008, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalf, S.; Dougherty, S.; Kruer, T.; Hasan, N.; Biyik-Sit, R.; Reynolds, L.; Clem, B.F. Selective loss of phosphoserine aminotransferase 1 (PSAT1) suppresses migration, invasion, and experimental metastasis in triple negative breast cancer. Clin. Exp. Metastasis 2020, 37, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, W.; Sun, X.; Xie, S.; Xu, X.; Liu, M.; Yang, C.; Li, M.; Zhang, W.; Liu, W.; et al. NudCL2 regulates cell migration by stabilizing both myosin-9 and LIS1 with Hsp90. Cell Death Dis. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Sarkar, S.; Egelhoff, T.; Baskaran, H. Insights into the roles of non-muscle myosin IIA in human keratinocyte migration. Cell. Mol. Bioeng. 2009, 2, 486–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Wu, N.; Cui, L.; Wu, Z.; Qiu, G. Filamin B: The next hotspot in skeletal research? J. Genet. Genom. 2017, 44, 335–342. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, M.H.; Moskowitz, I.P.; Mendonza, A.M.; Vidali, L.; Nakamura, F.; Kwiatkowski, D.J.; Walsh, C.A. Filamin A (FLNA) is required for cell–cell contact in vascular development and cardiac morphogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19836–19841. [Google Scholar] [CrossRef] [Green Version]
- Hamill, K.J.; Hiroyasu, S.; Colburn, Z.T.; Ventrella, R.V.; Hopkinson, S.B.; Skalli, O.; Jones, J.C.R. Alpha actinin-1 regulates cell-matrix adhesion organization in keratinocytes: Consequences for skin cell motility. J. Investig. Dermatol. 2015, 135, 1043–1052. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Park, Z.-Y.; Lin, D.; Brahmbhatt, A.A.; Rio, M.-C.; Yates, J.R., III; Klemke, R.L. Regulation of cell migration and survival by focal adhesion targeting of Lasp-1. J. Cell. Biol. 2004, 165, 421–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizzarro, V.; Fontanella, B.; Carratù, A.; Belvedere, R.; Marfella, R.; Parente, L.; Petrella, A. Annexin A1 N-terminal derived peptide Ac2-26 stimulates fibroblast migration in high glucose conditions. PLoS ONE 2012, 7, e45639. [Google Scholar]
- Tsukita, S.; Yonemura, S.; Tsukita, S. ERM (ezrin/radixin/moesin) family: From cytoskeleton to signal transduction. Curr. Opin. Cell Biol. 1997, 9, 70–75. [Google Scholar] [CrossRef]
- Jacob, J.T.; Coulombe, P.A.; Kwan, R.; Omary, M.B. Types I and II Keratin Intermediate Filaments. Cold Spring Harb. Perspect. Biol. 2018, 10, a018275. [Google Scholar] [CrossRef] [Green Version]
- Machesky, L.M.; Li, A. Fascin. Commun. Integr. Biol. 2010, 3, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Muneyuki, E.; Hisabori, T. ATP synthase—A marvellous rotary engine of the cell. Nat. Rev. Mol. Cell Biol. 2001, 2, 669–677. [Google Scholar] [CrossRef]
- Grüning, N.-M.; Du, D.; Keller, M.A.; Luisi, B.F.; Ralser, M. Inhibition of triosephosphate isomerase by phosphoenolpyruvate in the feedback-regulation of glycolysis. Open Biol. 2014, 4, 130232. [Google Scholar] [CrossRef] [Green Version]
- de Padua, M.C.; Delodi, G.; Vučetić, M.; Durivault, J.; Vial, V.; Bayer, P.; Noleto, G.R.; Mazure, N.M.; Ždralević, M.; Pouysségur, J. Disrupting glucose-6-phosphate isomerase fully suppresses the “Warburg effect” and activates OXPHOS with minimal impact on tumor growth except in hypoxia. Oncotarget 2017, 8, 87623–87637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, M.; Lu, T.; Fan, S.; Zhang, H.; Gong, R.; Sun, L.; Fu, Z.; Fan, L. Glucose-6-phosphate isomerase promotes the proliferation and inhibits the apoptosis in fibroblast-like synoviocytes in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 100. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Zhu, X.; Wu, H.; Wang, Y.; Hu, X. Perturbation of phosphoglycerate kinase 1 (PGK1) only marginally affects glycolysis in cancer cells. J. Biol. Chem. 2020, 295, 6425–6446. [Google Scholar] [CrossRef] [Green Version]
- Hanse, E.A.; Ruan, C.; Kachman, M.; Wang, D.; Lowman, X.H.; Kelekar, A. Cytosolic malate dehydrogenase activity helps support glycolysis in actively proliferating cells and cancer. Oncogene 2017, 36, 3915–3924. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Li, J.; Leonard, S.S.; Shi, X.; Costa, M.; Castranova, V.; Vallyathan, V.; Huang, C. Differential role of hydrogen peroxide in UV-induced signal transduction. Mol. Cell. Biochem. 2002, 234–235, 81–90. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. IκB kinases: Key regulators of the NF-κB pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef]
- Yeo, H.; Lee, J.Y.; Kim, J.; Ahn, S.S.; Jeong, J.Y.; Choi, J.H.; Lee, Y.H.; Shin, S.Y. Transcription factor EGR-1 transactivates the MMP1 gene promoter in response to TNFα in HaCaT keratinocytes. BMB Rep. 2020, 53, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Tobiume, K.; Matsuzawa, A.; Takahashi, T.; Nishitoh, H.; Morita, K.-I.; Takeda, K.; Minowa, O.; Miyazono, K.; Noda, T.; Ichijo, H. ASK1 is required for sustained activations of JNK/p38 MAP kinases and apoptosis. EMBO Rep. 2001, 2, 222–228. [Google Scholar] [CrossRef]
- Elliott, S.F.; Coon, C.I.; Hays, E.; Stadheim, T.A.; Vincenti, M.P. Bcl-3 is an interleukin-1–responsive gene in chondrocytes and synovial fibroblasts that activates transcription of the matrix metalloproteinase 1 gene. Arthritis Rheum. 2002, 46, 3230–3239. [Google Scholar] [CrossRef]
- Fanjul-Fernández, M.; Folgueras, A.R.; Cabrera, S.; López-Otín, C. Matrix metalloproteinases: Evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. Acta 2010, 1803, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Buckman, S.Y.; Gresham, A.; Hale, P.; Hruza, G.; Anast, J.; Masferrer, J.; Pentland, A.P. COX-2 expression is induced by UVB exposure in human skin: Implications for the development of skin cancer. Carcinogenesis 1998, 19, 723–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Hu, Y.; Jiang, X.; Chen, T.; Ma, Y.; Wu, S.; Sun, J.; Jiao, R.; Li, X.; Deng, L.; et al. Cyanidin-3-O-glucoside inhibits the UVB-induced ROS/COX-2 pathway in HaCaT cells. J. Photochem. Photobiol. B 2017, 177, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Tyagi, N.; Bhardwaj, A.; Rusu, L.; Palanki, R.; Vig, K.; Singh, S.R.; Singh, A.P.; Palanki, S.; Miller, M.E.; et al. Silver nanoparticles protect human keratinocytes against UVB radiation-induced DNA damage and apoptosis: Potential for prevention of skin carcinogenesis. Nanomedicine 2015, 11, 1265–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyo, C.-W.; Choi, J.H.; Oh, S.-M.; Choi, S.-Y. Oxidative stress-induced cyclin D1 depletion and its role in cell cycle processing. Biochim. Biophys. Acta 2013, 1830, 5316–5325. [Google Scholar] [CrossRef]
- Han, W.; He, Y.Y. Requirement for metalloproteinase-dependent ERK and AKT activation in UVB-induced G1-S cell cycle progression of human keratinocytes. Photochem. Photobiol. 2009, 85, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Hitomi, M.; Stacey, D.W. Variations in cyclin D1 levels through the cell cycle determine the proliferative fate of a cell. Cell Div. 2006, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.; Ahn, B.-N.; Kang, K.-H.; Kim, Y.-S.; Li, Y.-X.; Kong, C.-S.; Kim, S.-K.; Kim, D.G. Dioxinodehydroeckol protects human keratinocyte cells from UVB-induced apoptosis modulated by related genes Bax/Bcl-2 and caspase pathway. J. Photochem. Photobiol. B 2015, 153, 352–357. [Google Scholar] [CrossRef]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J. Photochem. Photobiol. B. 2019, 197, 111518. [Google Scholar] [CrossRef]
- Tian, Y.; Hoshino, T.; Chen, C.J.; E, Y.; Yabe, S.; Liu, W. The evaluation of whitening efficacy of cosmetic products using a human skin pigmentation spot model. Skin Res. Technol. 2009, 15, 218–223. [Google Scholar] [CrossRef]
- Goh, J.X.H.; Tan, L.T.-H.; Yew, H.C.; Pusparajah, P.; Lingham, P.; Long, C.M.; Lee, L.-H.; Goh, B.-H. Hydration effects of moisturizing gel on normal skin: A pilot study. Prog. Drug Discov. Biomed. Sci. 2019, 2, a0000023. [Google Scholar] [CrossRef]
- Adejokun, D.A.; Dodou, K. A novel method for the evaluation of the long-term stability of cream formulations containing natural oils. Cosmetics 2020, 7, 86. [Google Scholar] [CrossRef]
- Mosquera, T.; Peña, S.; Álvarez, P.; Noriega, P. Changes in skin elasticity and firmness caused by cosmetic formulas elaborated with essential oils of Aristeguietia glutinosa (matico) and Ocotea quixos (ishpingo). A statistical analysis. Cosmetics 2020, 7, 95. [Google Scholar] [CrossRef]
- Goh, B.H.; Chan, C.K.; Kamarudin, M.N.A.; Abdul Kadir, H. Swietenia macrophylla King induces mitochondrial-mediated apoptosis through p53 upregulation in HCT116 colorectal carcinoma cells. J. Ethnopharmacol. 2014, 153, 375–385. [Google Scholar] [CrossRef]
- Tan, L.T.-H.; Mahendra, C.K.; Yow, Y.-Y.; Chan, K.-G.; Khan, T.M.; Lee, L.-H.; Goh, B.-H. Streptomyces sp. MUM273b: A mangrove-derived potential source for antioxidant and UVB radiation protectants. Microbiol. Open 2019, 8, e859. [Google Scholar] [CrossRef] [Green Version]
- Mahendra, C.K.; Tan, L.T.-H.; Yap, W.H.; Chan, C.K.; Pusparajah, P.; Goh, B.H. An optimized cosmetic screening assay for ultraviolet B (UVB) protective property of natural products. Prog. Drug Discov. Biomed. Sci. 2019, 2, 1–6. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Kumari, Y.; Abidin, S.A.Z.; Othman, I.; Shaikh, M.F. Pilocarpine induced behavioral and biochemical alterations in chronic seizure-like condition in adult zebrafish. Int. J. Mol. Sci. 2020, 21, 2492. [Google Scholar] [CrossRef] [Green Version]
- Garcin, G.; Le Gallic, L.; Stoebner, P.-E.; Guezennec, A.; Guesnet, J.; Lavabre-Bertrand, T.; Martinez, J.; Meunier, L. Constitutive expression of MC1R in HaCaT keratinocytes inhibits basal and UVB-induced TNF-alpha production. Photochem. Photobiol. 2009, 85, 1440–1450. [Google Scholar] [CrossRef]
- Adachi, H.; Murakami, Y.; Tanaka, H.; Nakata, S. Increase of stratifin triggered by ultraviolet irradiation is possibly related to premature aging of human skin. Exp. Dermatol. 2014, 23, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.-H.; Chan, C.-K.; Chan, K.-G.; Pusparajah, P.; Khan, T.M.; Ser, H.-L.; Lee, L.-H.; Goh, B.-H. Streptomyces sp. MUM256: A Source for Apoptosis Inducing and Cell Cycle-Arresting Bioactive Compounds against Colon Cancer Cells. Cancers 2019, 11, 1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.-P.; Zhou, F.; Li, J.; Duan, S.-J. NEAT1 regulates MPP+-induced neuronal injury by targeting miR-124 in neuroblastoma cells. Neurosci. Lett. 2019, 708, 134340. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name of Protein (Uniprot ID) | Comparison Ratio of Untreated Controls | Comparison Ratio of UVB Control vs. S. macrophylla Treatment + UVB Samples | |||
|---|---|---|---|---|---|---|
| Non-UVB: UVB | UVB: SMCE + UVB | UVB: SMHF + UVB | UVB: SMEAF + UVB | UVB: SMWF + UVB | ||
| 1 | Chaperonin containing TCP1 subunit 2 β isoform CRA_b (CCT-β) (V9HW96) | 1.00:0.49 (↓) | N/S 1 | 1.00:1.41 (↑) | N/S | N/S |
| 2 | T-complex protein 1 subunit delta (CCT-δ) (A8K3C3) | 1.00:0.47 (↓) | N/S | 1.00:1.71 (↑) | N/S | 1.00:0.52 (↓) |
| 3 | T-complex protein 1 subunit gamma (CCT-γ) (B3KX11) | N/S | N/S | 1.00:1.58 (↑) | N/S | N/S |
| 4 | T-complex protein 1 subunit eta (CCT-η) (Q99832) | N/S | N/S | 1.00:1.92 (↑) | N/S | N/S |
| 5 | T-complex protein 1 subunit epsilon (CCT-ε) (P48643) | N/S | N/S | 1.00:2.39 (↑) | N/S | N/S |
| 6 | Ribosomal protein L12 variant (Q59FI9) | 1.00:0.34 (↓) | N/S | N/S | N/S | N/S |
| 7 | Ribosomal protein S8 (Q9BS10) | N/S | N/S | 1.00:1.90 (↑) | N/S | N/S |
| 8 | 40S ribosomal protein SA (C9J9K3) | N/S | N/S | 1.00:1.87 (↑) | N/S | N/S |
| 9 | 40S ribosomal protein S5 (M0QZN2) | 1.00:0.32 (↓) | N/S | N/S | N/S | 1.00:0.32 (↓) |
| 10 | 40S ribosomal protein S7 (P62081) | 1.00:0.45 (↓) | N/S | N/S | N/S | 1.00:0.29 (↓) |
| 11 | 40S ribosomal protein S10 (P46783) | 1.00:0.33 (↓) | N/S | N/S | N/S | 1.00:0.31 (↓) |
| 12 | 40S ribosomal protein S16 (M0R210 or M0R3H0) | 1.00:0.36 (↓) | N/S | N/S | N/S | 1.00:0.51 (↓) |
| 13 | 40S ribosomal protein S25 (P62851) | 1.00:0.48 (↓) | N/S | N/S | N/S | 1.00:0.50 (↓) |
| 14 | 60S ribosomal protein L6 (Q8N5Z7) | 1.00:0.31 (↓) | N/S | N/S | N/S | N/S |
| 15 | 60S ribosomal protein L18 (F8VUA6) | 1.00:0.51 (↓) | 1.00:0.22 (↓) | 1.00:0.68 (↓) | N/S | 1.00:0.44 (↓) |
| 16 | 60S ribosomal protein L22 (K7EJT5) | 1.00:0.37 (↓) | N/S | 1.00:1.71 (↑) | N/S | N/S |
| 17 | 60S ribosomal protein L24 (C9JXB8) | 1.00:0.48 (↓) | N/S | N/S | N/S | N/S |
| 18 | 60S ribosomal protein L29 (A0A024R326) | 1.00:0.50 (↓) | N/S | N/S | N/S | 1.00:0.39 (↓) |
| 19 | 60s acidic ribosomal protein P0 (Q53HW2) | N/S | N/S | N/S | N/S | 1.00:0.40 (↓) |
| 20 | 60s acidic ribosomal protein P2 (P05387) | N/S | N/S | 1.00:1.39 (↑) | N/S | 1.00:0.55 (↓) |
| 21 | Albumin (F6KPG5) | 1.00:2.66 (↑) | N/S | N/S | N/S | 1.00:0.41 (↓) |
| 22 | chloride intracellular channel (A0A1U9 × 8Y4) | N/S | N/S | N/S | N/S | 1.00:0.63 (↓) |
| 23 | chloride intracellular channel 1 (CLIC1) (O00299) | 1.00:0.56 (↓) | N/S | 1.00:1.55 (↑) | N/S | N/S |
| 24 | Cofilin 1 (V9HWI5) | 1.00:0.61 (↓) | N/S | 1.00:1.38 (↑) | N/S | 1.00:0.63 (↓) |
| 25 | Elongation factor 1-alpha (Q6IPN6 or Q53HM9) | 1.00:0.36 (↓) | N/S | 1.00:1.88 (↑) | N/S | N/S |
| 26 | Elongation factor 1-delta (A0A087X1X7 or E9PN91) | 1.00:0.33 (↓) | N/S | 1.00:1.51 (↑) | 1.00:1.49 (↑) | 1.00:0.66 (↓) |
| 27 | Eukaryotic translation elongation factor 2 (eEF2) (Epididymis secretory sperm binding protein) (A0A384N6H1) | 1.00:0.55 (↓) | N/S | 1.00:1.67 (↑) | N/S | 1.00:0.41 (↓) |
| 28 | Glycine-tRNA ligase (P41250) | 1.00:0.28 (↓) | N/S | 1.00:7.22 (↑) | N/S | N/S |
| 29 | Heat shock protein (HSP)-10 kDa (Chaperonin 10) (Epididymis secretory sperm binding protein) (A0A384N6A4) | N/S | N/S | 1.00:1.48 (↑) | N/S | 1.00:0.53 (↓) |
| 30 | HSP-60 kDa mitochondrial (P10809) | N/S | N/S | N/S | N/S | 1.00:0.52 (↓) |
| 31 | HSP-70 kDa protein 1A variant (Q59EJ3) | N/S | N/S | N/S | N/S | 1.00:0.54 (↓) |
| 32 | HSP-70 kDa protein 4 (Q59GF8) | N/S | N/S | 1.00:1.36 (↑) | N/S | N/S |
| 33 | HSP-70 family protein 5 (Epididymis secretory sperm binding protein Li 89n) (78 kDa glucose-regulated protein) (V9HWB4) | N/S | N/S | N/S | N/S | 1.00:0.54 (↓) |
| 34 | HSP-70 kDa protein 8, isoform CRA_a (Epididymis luminal protein 33) (V9HW22) | 1.00:0.52 (↓) | N/S | 1.00:1.50 (↑) | N/S | N/S |
| 35 | HSP-70 kDa protein 9 (75 kDa glucose-regulated protein) (B7Z4V2) | N/S | N/S | 1.00:1.35 (↑) | N/S | N/S |
| 36 | HSP 90α (P07900) | 1.00:0.60 (↓) | N/S | 1.00:1.51 (↑) | N/S | 1.00:0.51 (↓) |
| 37 | Gluthathione S-transferase (GST)-pi (Epididymis secretory protein Li 22) (V9HWE9) | 1.00:0.63 (↓) | N/S | 1.00:1.45 (↑) | N/S | 1.00:0.58 (↓) |
| 38 | Poly (RC) binding protein 1 (Epididymis secretory protein Li 85) (Q53SS8) | 1.00:0.32 (↓) | N/S | 1.00:2.10 (↑) | N/S | 1.00:0.56 (↓) |
| 39 | Histone H2A type 1-A (Q96QV6) | N/S | N/S | N/S | 1.00:1.41 (↑) | N/S |
| 40 | Histone H2A type 2-B (Q8IUE6) | 1.00:2.75 (↑) | N/S | 1.00:1.66 (↑) | N/S | 1.00:0.56 (↓) |
| 41 | Histone H2B (A8K9J7 or B4DR52) | 1.00:2.86 (↑) | N/S | N/S | N/S | 1.00:0.40 (↓) |
| 42 | Histone H1.2 (P16403) | N/S | N/S | 1.00:1.69 (↑) | 1.00:1.41 (↑) | 1.00:0.62 (↓) |
| 43 | Histone H1.5 (P16401) | N/S | N/S | 1.00:2.01 (↑) | N/S | N/S |
| 44 | Histone H4 (B2R4R0) | N/S | N/S | N/S | N/S | 1.00:0.50 (↓) |
| 45 | Myosin light polypeptide 6 (F8W1R7) | 1.00:0.43 (↓) | N/S | N/S | N/S | N/S |
| 46 | Myosin 9 (P35579) | 1.00:0.48 (↓) | N/S | N/S | N/S | 1.00:0.52 (↓) |
| 47 | Nucleophosmin (Nucleolar phosphoprotein B23 numatrin) isoform CRA_f (A0A0S2Z4G7) | 1.00:0.71 (↓) | N/S | 1.00:1.34 (↑) | N/S | 1.00:0.43 (↓) |
| 48 | Peptidyl-prolyl cis-trans isomerase (V9HWF5) | 1.00:0.56 (↓) | N/S | N/S | N/S | 1.00:0.43 (↓) |
| 49 | PRDX-1 (A0A384NPQ2 or A0A0A0MSI0) | 1.00:0.57 (↓) | N/S | 1.00:1.67 (↑) | N/S | 1.00:0.43 (↓) |
| 50 | PRDX-2 (B4DF70) | N/S | N/S | N/S | N/S | 1.00:0.51 (↓) |
| 51 | PRDX-3 (Thioredoxin-dependent peroxide reductase mitochondrial) (P30048) | N/S | N/S | 1.00:0.58 (↓) | 1.00:0.73 (↓) | N/S |
| 52 | PRDX-6 (P30041) | 1.00:0.58 (↓) | N/S | N/S | N/S | 1.00:0.42 (↓) |
| 53 | Phosphoglycerate kinase 1 (P00558) | 1.00:0.61 (↓) | N/S | 1.00:1.58 (↑) | N/S | 1.00:0.50 (↓) |
| 54 | Phosphoserine aminotransferase (Q9Y617) | 1.00:0.56 (↓) | N/S | 1.00:1.46 (↑) | N/S | 1.00:0.39 (↓) |
| 55 | Profilin-1 (P07737) | 1.00:0.63 (↓) | N/S | 1.00:1.42 (↑) | N/S | 1.00:0.56 (↓) |
| 56 | PDI (A0A024R8S5) | N/S | N/S | 1.00:1.52 (↑) | N/S | 1.00:0.63 (↓) |
| 57 | PDI-A3 (P30101) | N/S | N/S | 1.00:1.66 (↑) | 1.00:0.76 (↓) | N/S |
| 58 | PDI-A4 (P13667) | N/S | N/S | 1.00:2.30 (↑) | N/S | 1.00:0.60 (↓) |
| 59 | PDI-A6 (Endoplasmic reticulum protein 5) (Q15084) | 1.00:1.74 (↑) | N/S | N/S | N/S | 1.00:0.43 (↓) |
| 60 | Protein S100 (A0A590UJ49 or B2R5H0) | 1.00:0.30 (↓) | N/S | 1.00:1.47 (↑) | N/S | 1.00:0.33 (↓) |
| 61 | Protein-arginine deiminase type-1 (Q9ULC6) | 1.00:15.79 (↑) | N/S | N/S | N/S | 1.00:0.21 (↓) |
| 62 | Pyruvate kinase PKM (P14618) | 1.00:0.67 (↓) | N/S | 1.00:1.33 (↑) | N/S | 1.00:0.48 (↓) |
| 63 | RACK-1 (P63244 or D6RF23) | 1.00:0.22 (↓) | N/S | N/S | N/S | 1.00:9.32 (↑) |
| 64 | Signal recognition particle 14 kDa protein (SRP-14) (P37108) | 1.00:0.33 (↓) | N/S | N/S | N/S | N/S |
| 65 | SYNCRIP protein (Q05CK9) | 1.00:0.48 (↓) | N/S | N/S | N/S | N/S |
| 66 | Transketolase (B3KSI4) | 1.00:0.52 (↓) | N/S | N/S | N/S | N/S |
| 67 | D-3-phosphoglycerate dehydrogenase (B3KSC3) | 1.00:0.50 (↓) | N/S | N/S | N/S | N/S |
| 68 | Fumarate hydratase (Epididymis secretory sperm binding protein) (A0A0S2Z4C3) | N/S | 1.00:2.02 (↑) | 1.00:2.43 (↑) | N/S | N/S |
| 69 | Annexin A1 (Q5TZZ9) | N/S | N/S | 1.00:1.46 (↑) | N/S | 1.00:0.43 (↓) |
| 70 | Annexin A2 (A0A024R5Z7) | N/S | N/S | N/S | N/S | 1.00:0.46 (↓) |
| 71 | Annexin A3 (P12429) | N/S | 1.00:0.63 (↓) | N/S | 1.00:0.52 (↓) | 1.00:0.42 (↓) |
| 72 | Annexin A5 (P08758) | N/S | N/S | N/S | N/S | 1.00:0.42 (↓) |
| 73 | Filamin A (Q60FE6) | N/S | N/S | 1.00:1.54 (↑) | N/S | 1.00:0.36 (↓) |
| 74 | Filamin B β (Actin binding protein 278) isoform CRA_a (A0A024R321) | N/S | 1.00:0.44 (↓) | N/S | N/S | N/S |
| 75 | 3-phosphoglycerate dehydrogenase (Q9UMY2 or Q9UMY3) | N/S | N/S | 1.00:2.27 (↑) | N/S | 1.00:0.26 (↓) |
| 76 | 14-3-3 protein α/β (P31946) | N/S | N/S | N/S | N/S | 1.00:0.60 (↓) |
| 77 | 14-3-3 protein σ (P31947) | N/S | N/S | 1.00:1.97 (↑) | N/S | 1.00:0.47 (↓) |
| 78 | 14-3-3 protein γ (P61981) | N/S | N/S | N/S | N/S | 1.00:0.18 (↓) |
| 79 | 14-3-3 protein ε (P62258) | N/S | N/S | N/S | N/S | 1.00:0.38 (↓) |
| 80 | Ubiquitin-activating enzyme E1 (Testicular secretory protein Li 63) (A0A024R1A3) | N/S | N/S | 1.00:1.84 (↑) | N/S | N/S |
| 81 | AHNAK (Desmoyokin) (Q09666) | N/S | N/S | 1.00:0.55 (↓) | N/S | N/S |
| 82 | Keratin type I cytoskeletal 14 (P02533) | N/S | N/S | 1.00:1.86 (↑) | 1.00:1.88 (↑) | N/S |
| 83 | Keratin type II cytoskeletal 8 (P05787) | N/S | N/S | 1.00:1.48 (↑) | N/S | 1.00:0.52 (↓) |
| 84 | Heterogeneous nuclear ribonucleoprotein (hnRP) D0 (D6RF44) | N/S | N/S | 1.00:1.74 (↑) | N/S | N/S |
| 85 | hnRP K (B4DFF1 or B4DUQ1) | N/S | N/S | 1.00:1.54 (↑) | N/S | 1.00:0.51 (↓) |
| 86 | Adenylyl cyclase-associated protein (CAP1) (B4DI38) | N/S | N/S | 1.00:1.54 (↑) | N/S | 1.00:0.38 (↓) |
| 87 | Reticulon-4 (Q6IPN0 or Q9NQC3) | N/S | N/S | 1.00:1.70 (↑) | N/S | 1.00:0.64 (↓) |
| 88 | Malate dehydrogenase cytoplasmic (P40925) | N/S | N/S | 1.00:1.70 (↑) | N/S | 1.00:0.72 (↓) |
| 89 | Transketolase (A0A0B4J1R6) | N/S | N/S | 1.00:1.62 (↑) | N/S | 1.00:0.34 (↓) |
| 90 | Glucose-6-phosphate isomerase (P06744) | N/S | N/S | 1.00:1.61 (↑) | N/S | 1.00:0.36 (↓) |
| 91 | Proteasome subunit alpha type (Q53GF5 or H0YLC2) | N/S | N/S | 1.00:1.45 (↑) | N/S | 1.00:0.32 (↓) |
| 92 | Proteasome activator complex subunit 2 (Q86SZ7) | N/S | N/S | 1.00:1.63 (↑) | N/S | N/S |
| 93 | Proteasome subunit β type-2 (P49721) | N/S | N/S | N/S | N/S | 1.00:0.42 (↓) |
| 94 | Proteasome subunit β type-3 (A0A087WXQ8) | N/S | N/S | 1.00:1.59 (↑) | N/S | N/S |
| 95 | Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (P04406) | N/S | N/S | 1.00:1.57 (↑) | N/S | 1.00:0.47 (↓) |
| 96 | Proliferating cell nuclear antigen (PCNA) (P12004 or Q6FHF5) | N/S | N/S | 1.00:1.55 (↑) | N/S | 1.00:0.53 (↓) |
| 97 | Exportin-2 (P55060) | N/S | N/S | 1.00:1.62 (↑) | 1.00:1.33 (↑) | N/S |
| 98 | Serpin peptidase inhibitor clade B (Ovalbumin) member 5 isoform CRA_b (SERPINB5) (A0A024R2B6) | N/S | N/S | 1.00:1.52 (↑) | N/S | 1.00:0.51 (↓) |
| 99 | Collagen-binding protein (Serpin H1) (B4DN87) | N/S | N/S | 1.00:1.80 (↑) | N/S | N/S |
| 100 | Ezrin (Q6NUR7) | N/S | N/S | 1.00:1.51 (↑) | N/S | 1.00:0.19 (↓) |
| 101 | Tubulin beta chain (A0A384NYT8) | N/S | N/S | 1.00:1.47 (↑) | N/S | 1.00:0.49 (↓) |
| 102 | Thioredoxin (P10599) | N/S | N/S | 1.00:1.40 (↑) | N/S | 1.00:0.54 (↓) |
| 103 | Thioredoxin domain-containing protein 17 (TXNDC17) (Testicular tissue protein Li 214) (A0A140VJY7) | N/S | N/S | 1.00:1.47 (↑) | N/S | N/S |
| 104 | Alpha-enolase (P06733) | N/S | N/S | 1.00:1.46 (↑) | N/S | 1.00:0.46 (↓) |
| 105 | Triosephosphate isomerase (V9HWK1) | N/S | N/S | 1.00:1.45 (↑) | N/S | 1.00:0.44 (↓) |
| 106 | Alpha-actinin-1 (P12814) | N/S | N/S | 1.00:1.44 (↑) | N/S | N/S |
| 107 | Calreticulin variant (Q53G71) | N/S | N/S | 1.00:1.44 (↑) | N/S | 1.00:0.42 (↓) |
| 108 | L-lactate dehydrogenase A (V9HWB9) | N/S | N/S | 1.00:1.38 (↑) | N/S | 1.00:0.35 (↓) |
| 109 | L-lactate dehydrogenase B (Q5U077) | N/S | N/S | 1.00:1.39 (↑) | N/S | 1.00:0.42 (↓) |
| 110 | ATP synthase subunit α, mitochondrial (V9HW26) | N/S | N/S | N/S | N/S | 1.00:0.44 (↓) |
| 111 | ATP synthase subunit β, mitochondrial (P06576) | N/S | N/S | 1.00:1.37 (↑) | N/S | 1.00:0.59 (↓) |
| 112 | Endoplasmin (P14625) | N/S | N/S | 1.00:1.36 (↑) | N/S | 1.00:0.57 (↓) |
| 113 | Nucleosome assembly protein 1-like 1 (F8W020) | N/S | N/S | 1.00:1.35 (↑) | N/S | N/S |
| 114 | Neutral amino acid transporter B(0) (Q15758) | N/S | N/S | 1.00:1.38 (↑) | N/S | 1.00:0.48 (↓) |
| 115 | LIM and SH3 domain protein 1 (LASP-1) (A8K1D2) | N/S | N/S | 1.00:1.36 (↑) | N/S | N/S |
| 116 | Polyubiquitin-C (F5GYU3) | N/S | N/S | N/S | 1.00:0.56 (↓) | 1.00:0.33 (↓) |
| 117 | HNRPCL1 protein (Q6PKD2) | N/S | N/S | N/S | 1.00:0.46 (↓) | N/S |
| 118 | Fascin (B3KTA3) | N/S | N/S | N/S | 1.00:0.64 (↓) | 1.00:0.36 (↓) |
| 119 | Cathepsin D (A0A1B0GW44) | N/S | N/S | N/S | 1.00:0.70 (↓) | 1.00:0.49 (↓) |
| 120 | Prothymosin alpha (Q15203) | N/S | N/S | N/S | 1.00:0.20 (↓) | 1.00:0.27 (↓) |
| 121 | GTP-binding nuclear protein Ran (P62826) | N/S | N/S | N/S | 1.00:0.72 (↓) | 1.00:0.41 (↓) |
| 122 | NOLC1 (B2RAU8) | N/S | N/S | N/S | 1.00:1.39 (↑) | N/S |
| 123 | Protein kinase C substrate 80K-H isoform 1 (A0A0S2Z4D8) | N/S | N/S | N/S | 1.00:1.35 (↑) | N/S |
| 124 | Actin cytoplasmic 1 (P60709) | N/S | N/S | N/S | N/S | 1.00:0.25 (↓) |
| 125 | Actin cytoplasmic 2 (P63261) | N/S | N/S | N/S | N/S | 1.00:0.47 (↓) |
| 126 | Rab GDP dissociation inhibitor β (P50395) | N/S | N/S | N/S | N/S | 1.00:0.34 (↓) |
| 127 | Eukaryotic translation initiation factor 5A (I3L397) | N/S | N/S | N/S | N/S | 1.00:0.39 (↓) |
| 128 | Phosphoglycerate mutase (Q6FHU2) | N/S | N/S | N/S | N/S | 1.00:0.43 (↓) |
| 129 | Neutral alpha-glucosidase AB (Epididymis secretory sperm binding protein Li 164nA) (V9HWJ0) | N/S | N/S | N/S | N/S | 1.00:0.33 (↓) |
| 130 | Adenosylhomocysteinase (P23526) | N/S | N/S | N/S | N/S | 1.00:0.44 (↓) |
| 131 | 4F2 cell-surface antigen heavy chain (F5GZS6) | N/S | N/S | N/S | N/S | 1.00:0.44 (↓) |
| 132 | Protein SET (A0A0C4DFV9) | N/S | N/S | N/S | N/S | 1.00:0.45 (↓) |
| 133 | Inorganic pyrophosphatase (Epididymis secretory sperm binding protein Li 66p) (V9HWB5) | N/S | N/S | N/S | N/S | 1.00:0.42 (↓) |
| 134 | Aspartate aminotransferase (B3KUZ8) | N/S | N/S | N/S | N/S | 1.00:0.47 (↓) |
| 135 | Plasminogen activator inhibitor 2 (P05120) | N/S | N/S | N/S | N/S | 1.00:0.52 (↓) |
| 136 | Acidic leucine-rich nuclear phosphoprotein 32 family member A (ANP32B) (H0YN26) | N/S | N/S | N/S | N/S | 1.00:0.52 (↓) |
| 137 | Complement component 1 Q subcomponent-binding protein mitochondrial (Q07021) | N/S | N/S | N/S | N/S | 1.00:0.52 (↓) |
| 138 | Fructose-bisphosphate aldolase A (P04075) | N/S | N/S | N/S | N/S | 1.00:0.53 (↓) |
| 139 | Transaldolase (A0A140VK56) | N/S | N/S | N/S | N/S | 1.00:0.53 (↓) |
| 140 | Macrophage migration inhibitory factor (P14174) | N/S | N/S | N/S | N/S | 1.00:0.55 (↓) |
| 141 | Endoplasmic reticulum resident protein 29 (P30040) | N/S | N/S | N/S | N/S | 1.00:0.49 (↓) |
| 142 | Calmodulin-2 (P0DP24) | N/S | N/S | N/S | N/S | 1.00:0.59 (↓) |
| 143 | RAN binding protein 1 isoform CRA_g (A0A140VK94) | N/S | N/S | N/S | N/S | 1.00:0.59 (↓) |
| 144 | Vinculin isoform CRA_c (A0A024QZN4) | N/S | N/S | N/S | N/S | 1.00:0.32 (↓) |
| 145 | Small ubiquitin-related modifier (A0A024R8S3) | N/S | N/S | N/S | N/S | 1.00:0.37 (↓) |
| 146 | Calnexin (P27824) | N/S | N/S | N/S | N/S | 1.00:0.61 (↓) |
| 147 | Nucleolin (P19338) | N/S | N/S | N/S | N/S | 1.00:0.49 (↓) |
| 148 | RPLP1 protein (Q6ICQ4) | N/S | N/S | N/S | N/S | 1.00:0.64 (↓) |
| 149 | FK506 binding protein 12 (Q1JUQ3) | N/S | N/S | N/S | N/S | 1.00:0.12 (↓) |
| 150 | Lysosome-associated membrane glycoprotein 1 (B3KRY3) | N/S | N/S | N/S | N/S | 1.00:0.26 (↓) |
| 151 | Transgelin-2 (P37802) | N/S | N/S | N/S | N/S | 1.00:0.52 (↓) |
| Target Gene | Sequence | References |
|---|---|---|
| TNF-α | Forward: 5′-CCAGGCAGTCAGATCATCTTCTC-3′ | [130] |
| Reverse: 5′-AGCTTGAGGGTTTGCTACAACAT-3′ | ||
| NF-κB | Forward: 5′-GACGAGAACGGAGACACA-3′ | Designed with NCBI Primer Blast |
| Reverse: 5′-TGGTTGGTAGGTTGACAAC-3′ | ||
| COX-2 | Forward 5′-TGCGCCTTTTCAAGGATGGA-3′ | Designed with NCBI Primer Blast |
| Reverse 5′-CCCCACAGCAAACCGTAGAT-3′ | ||
| MMP-1 | Forward 5′-GGGAGATCATCGGGACAACTC-3′ | [131] |
| Reverse 5′-TGAGCATCCCCTCCAATACC-3′ | ||
| Cyclin D1 | Forward 5′-TGCGCTGCTACCGTTGACT-3′ | [132] |
| Reverse 5′-AGCGATGTGAATATTTCCAAACC-3′ | ||
| Bax | Forward 5′-GTCGCCCTTTTCTACTTTGCCAG-3′ | [126] |
| Reverse 5′-TCCAGCCCAACAGCCGCTCC-3′ | ||
| 18S ribosomal RNA | Forward: 5′-GGCCCTGTAATTGGAATGAGTC-3′ | [133] |
| Reverse: 5′-CCAAGATCCAACTACGAGCTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahendra, C.K.; Abidin, S.A.Z.; Htar, T.T.; Chuah, L.-H.; Khan, S.U.; Ming, L.C.; Tang, S.Y.; Pusparajah, P.; Goh, B.H. Counteracting the Ramifications of UVB Irradiation and Photoaging with Swietenia macrophylla King Seed. Molecules 2021, 26, 2000. https://doi.org/10.3390/molecules26072000
Mahendra CK, Abidin SAZ, Htar TT, Chuah L-H, Khan SU, Ming LC, Tang SY, Pusparajah P, Goh BH. Counteracting the Ramifications of UVB Irradiation and Photoaging with Swietenia macrophylla King Seed. Molecules. 2021; 26(7):2000. https://doi.org/10.3390/molecules26072000
Chicago/Turabian StyleMahendra, Camille Keisha, Syafiq Asnawi Zainal Abidin, Thet Thet Htar, Lay-Hong Chuah, Shafi Ullah Khan, Long Chiau Ming, Siah Ying Tang, Priyia Pusparajah, and Bey Hing Goh. 2021. "Counteracting the Ramifications of UVB Irradiation and Photoaging with Swietenia macrophylla King Seed" Molecules 26, no. 7: 2000. https://doi.org/10.3390/molecules26072000