
Xanthine Oxidase Inhibitors from Filipendula ulmaria (L.) Maxim. and Their Efficient Detections by HPTLC and HPLC Analyses

 , , ,
, , ,
Abstract
:1. Introduction
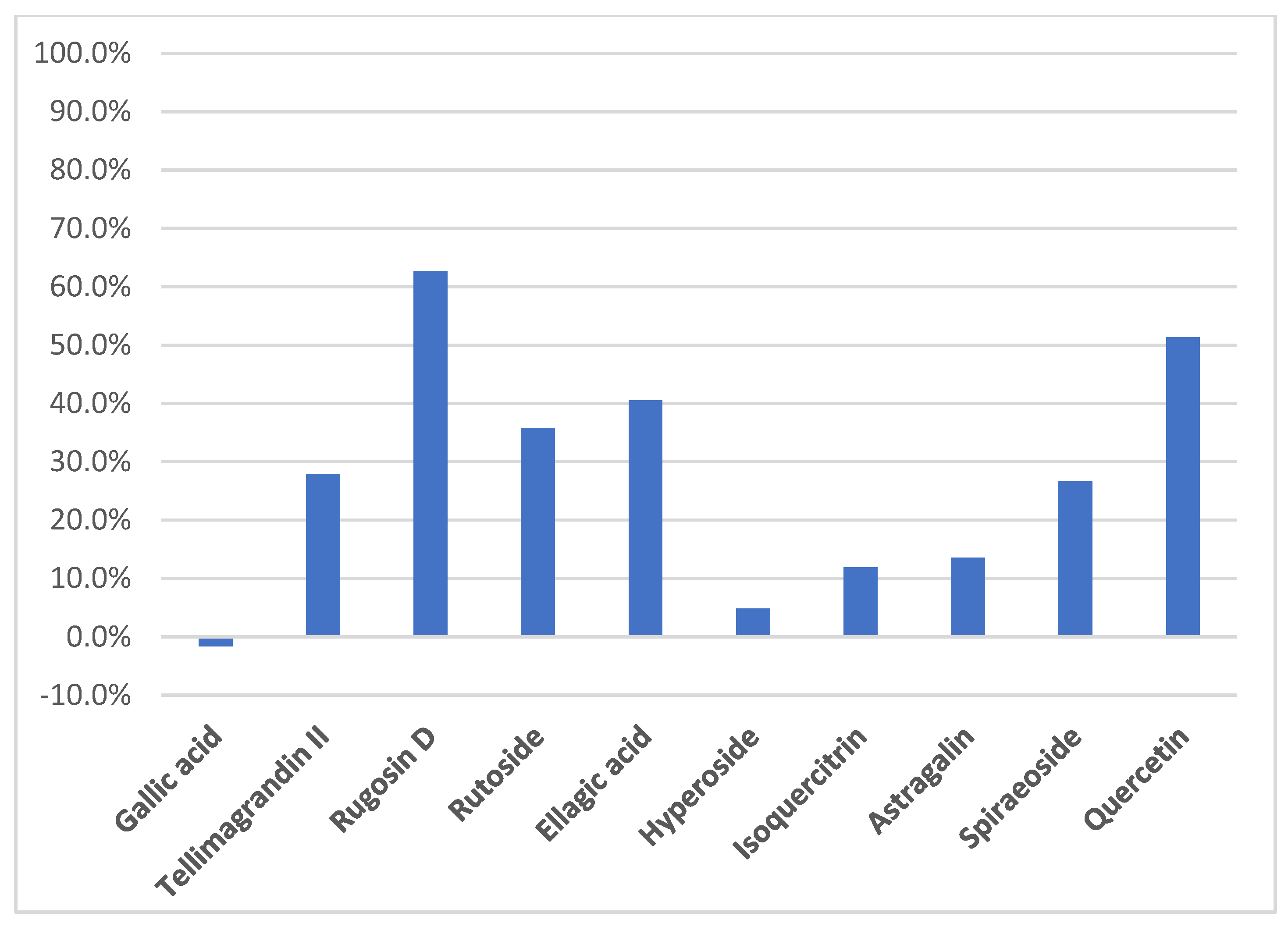
2. Results and Discussion
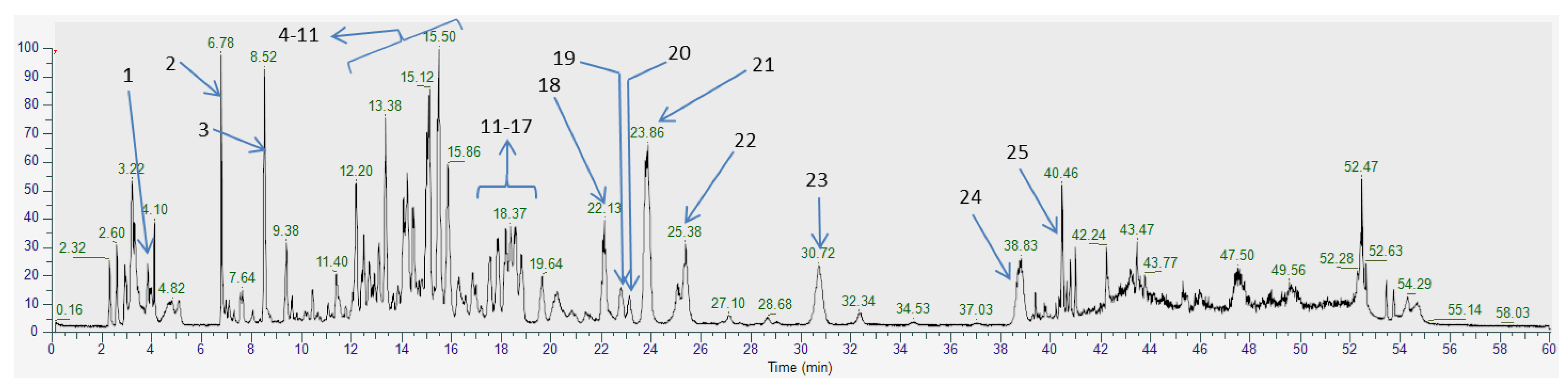
2.1. Phytochemical Profile of F. ulmaria Aerial Parts Methanolic Extract
2.2. Liquid/Liquid Partition
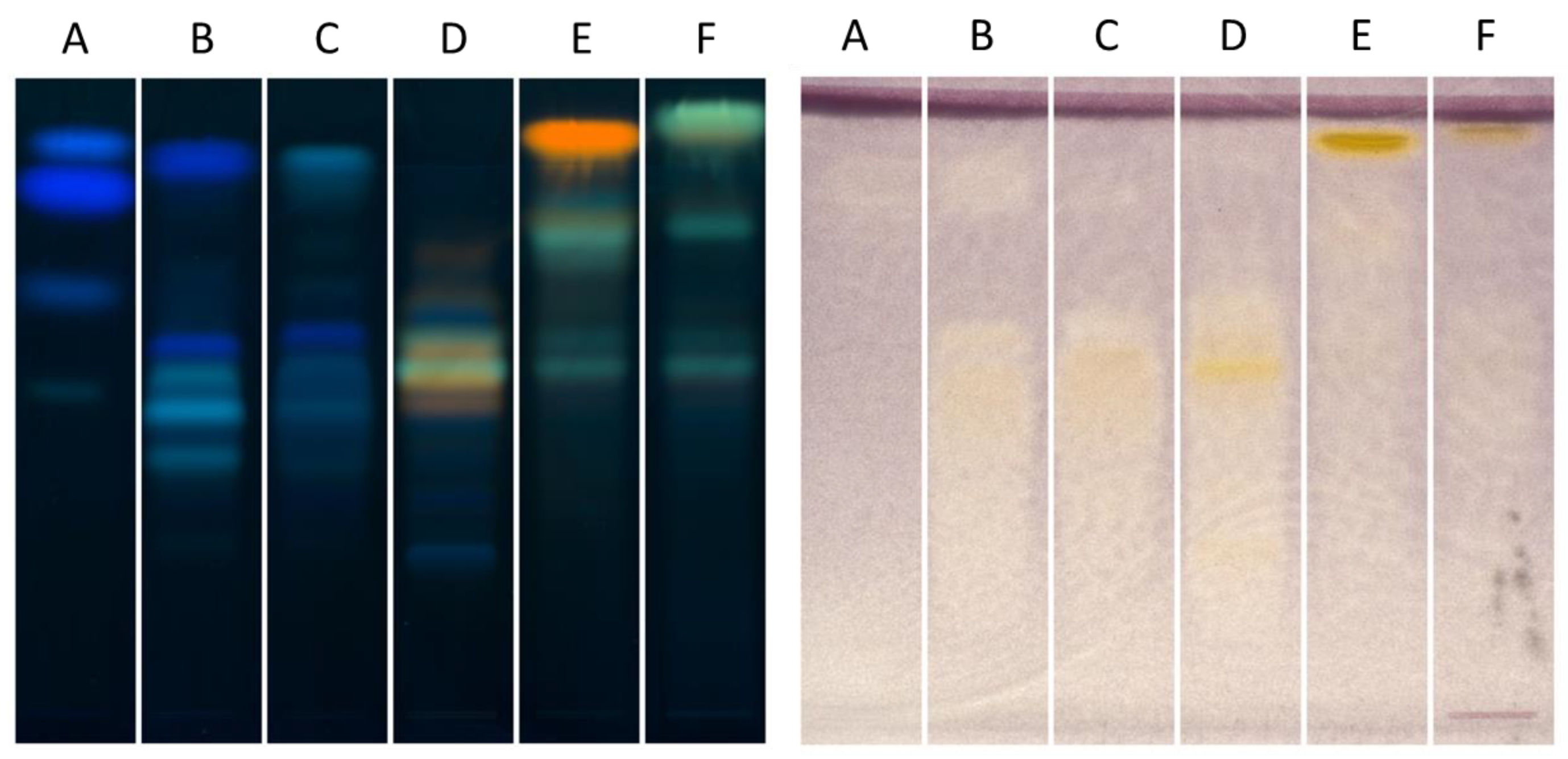
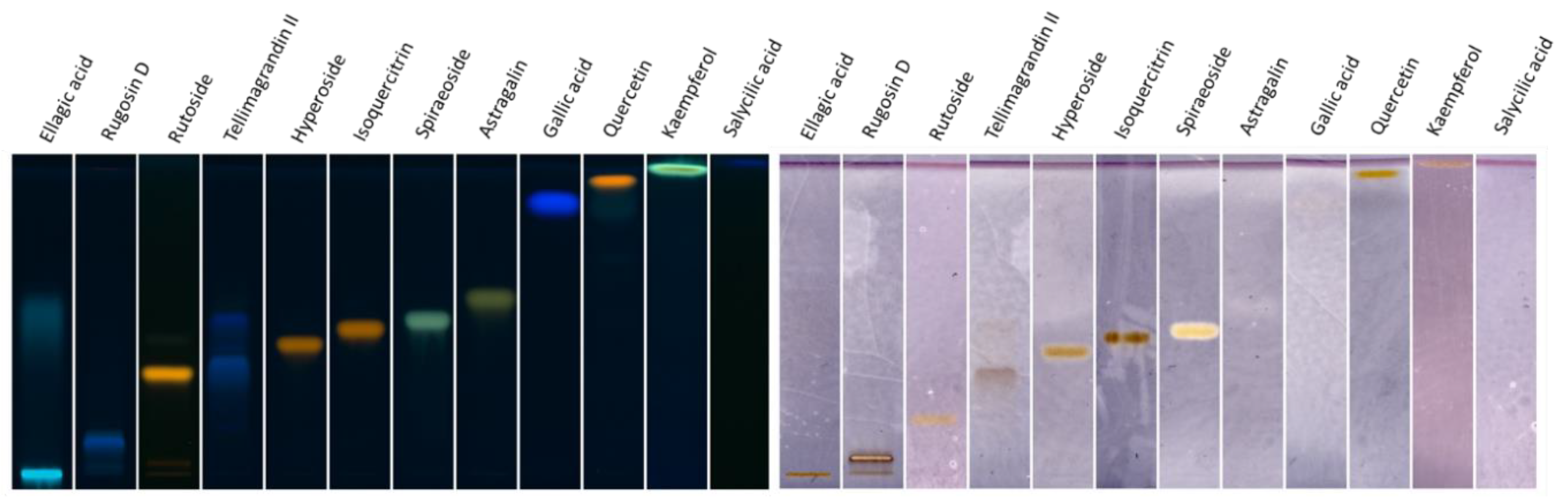
2.3. Investigation of Ethyl Acetate Fraction through HPTLC XO Bioautography
2.4. Determination of Active Compounds by HPTLC Bioautography
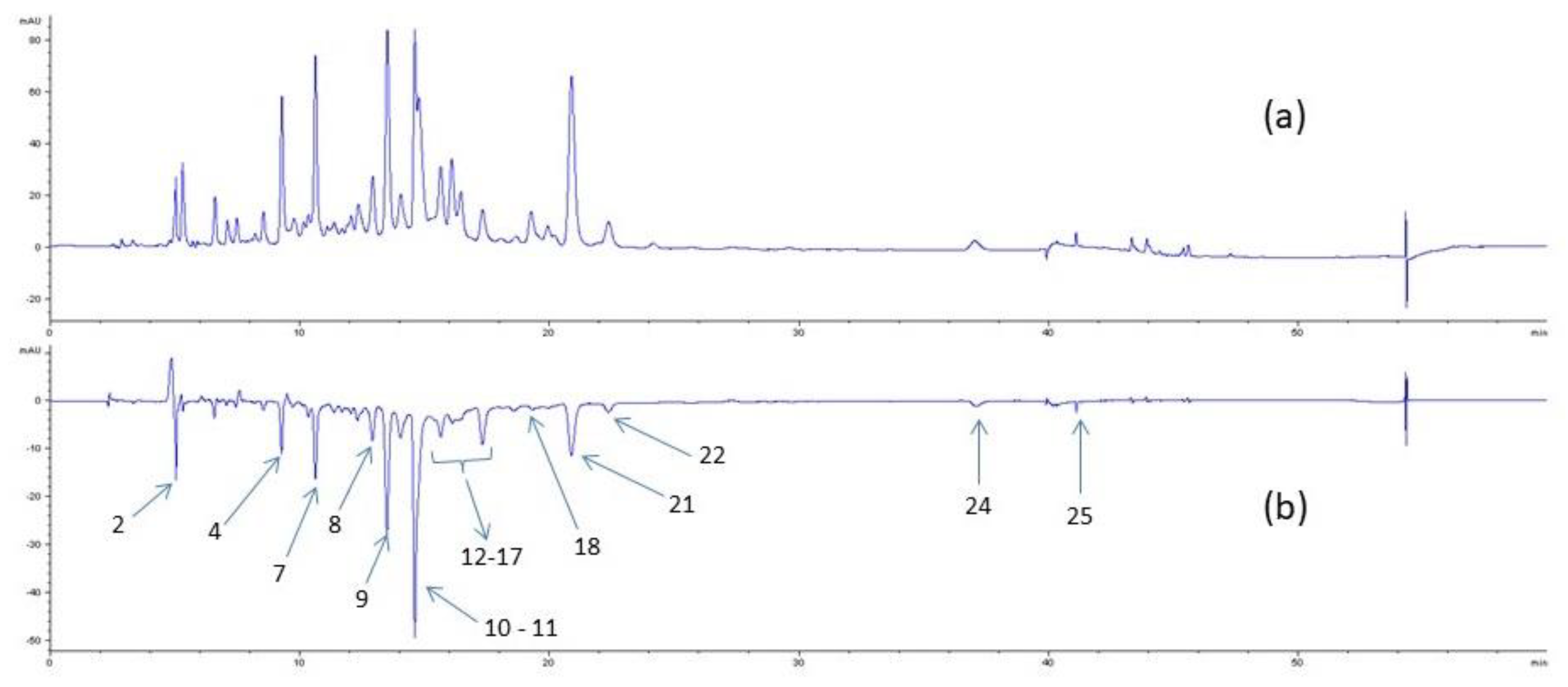
2.5. Detection of Potential Inhibitors by HPLC Analysis
3. Conclusions
4. Materials and Methods
4.1. Plant Material
4.2. Standards and Reagents
4.3. Extraction, Characterisation and Isolation
4.3.1. Extraction of Aerial Parts of F. ulmaria
4.3.2. Characterization of the Crude Extracts
4.3.3. Purification of Compounds from F. ulmaria Aerial Parts
4.4. Xanthine Oxidase Inhibitory Activity
4.5. HPTLC Analysis
4.5.1. Equipment
4.5.2. General Procedure
4.5.3. Natural Products Reagent Derivatization
4.5.4. Xanthine Oxidase Bioautography
4.5.5. Measurement of XO Interaction Involving HPLC Method
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barros, L.; Cabrita, L.; Vilas Boas, M.; Carvalho, A.M.; Fereira, I.C.F.R. Chemical, biochemical and electrochemical assays to evaluate phytochemicals and antioxidant activity of wild plants. Food Chem. 2011, 127, 1600–1608. [Google Scholar] [CrossRef]
- Jarić, S.; Popović, Z.; Mačukanović-Jocić, M.; Djurdjević, L.; Mijatović, M.; Karadžić, B.; Mitrović, M.; Pavlović, P. An ethnobotanical study on the usage of wild medicinal herbs from Kopaonik Mountain (Central Serbia). J. Ethnopharmacol. 2007, 111, 160–175. [Google Scholar] [CrossRef]
- Barros, L.; Alves, M.; Dueñas, C.T.; Silva, S.; Oliviera, R.; Carvalho, A.M.; Henriques, M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Characterization of phenolic compounds in wild medicinal flowers from Portugal by HPLC-DAD-ESI/MS and evaluation of antifungal properties. Ind. Crops Prod. 2013, 44, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, V.G.; Alexandrov, V.A.; Semenov, A.L.; Vysochina, G.I.; Kostikova, V.A.; Baranenko, D.A. The inhibitory effect of Filipendula ulmaria (L.) Maxim. on colorectal carcinogenesis induced in rats by methylnitrosourea. J. Ethnopharmacol. 2018, 227, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Katanić, J.; Boroja, T.; Mihailović, V.; Nikles, S.; Pan, S.-P.; Rosić, G.; Selaković, D.; Joksimović, J.; Mitrović, S.; Bauer, R. In vitro and in vivo assessment of meadowsweet (Filipendula ulmaria) as anti-inflammatory agent. J. Ethnopharmacol. 2016, 193, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.J.; Sousa, D.; Lima, R.T.; Carvalho, A.M.; Ferreira, I.C.F.R.M.; Vasconcelos, H. Flower extracts of Filipendula ulmaria (L.) Maxim inhibit the proliferation of the NCI-H460 tumour cell line. Ind. Crops Prod. 2014, 59, 149–153. [Google Scholar] [CrossRef]
- Bijttebier, S.; Van der Auwera, A.; Voorspoels, S.; Noten, B.; Hermans, N.; Pieters, L.; Apers, S. A first step in the quest for the active constituents in Filipendula ulmaria (Meadowsweet): Comprehensive phytochemical identification by liquid chromatography coupled to quadrupole-orbitrap mass spectrometry. Planta Med. 2016, 82, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Fecka, I. Qualitative and quantitative determination of hydrolysable tannins and other polyphenols in herbal products from Meadowsweet and Dog Rose. Phytochem. Anal. 2009, 20, 177–190. [Google Scholar] [CrossRef]
- Katanić, J.; Boroja, T.; Stanković, N.; Mihailović, V.; Mladenović, M.; Kreft, S.; Vrvić, M.M. Bioactivity, stability and phenolic characterization of Filipendula ulmaria (L.) Maxim. Food Funct. 2015, 6, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Ragab, G.; Elshahaly, M.; Bardin, T. Gout: An old disease in new perspective—A review. J. Adv. Res. 2017, 8, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Ying, L.; Calomme, M.; Hu, J.P.; Cimanga, K.; Van Poel, B.; Pieters, L.; Vlietinck, A.J.; Vanden Berghe, D. Structure-activity relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef]
- Robinson, P.C. Gout—An update of aetiology, genetics, co-morbidities and management. Maturitas 2018, 118, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.T.; Awale, S.; Tezuka, Y.; Ueda, J.-Y.; Tran, Q.L.; Kadota, S. Xanthine oxidase inhibitors from the flowers of Chrysanthemum sinense. Planta Med. 2006, 72, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hunyadi, A.; Martins, A.; Danko, B.; Chuang, D.-W.; Trouillas, P.; Chang, F.-R.; Wu, Y.-C.; Falkay, G. Discovery of the first non-planar flavonoid that can strongly inhibit xanthine oxidase: Protoapigenone 1’-O-propargyl ether. Tetrahedron Lett. 2013, 54, 6529–6532. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, G.; Liao, Y.; Pan, J.; Gong, D. Dietary flavonoids as xanthine oxidase inhibitors: Structure-affinity and structure-activity relationship. J. Agric. Food. Chem. 2015, 63, 7784–7794. [Google Scholar] [CrossRef]
- Nagao, A.; Seki, M.; Kobayashi, H. Inhibition of xanthine oxidase by flavonoids. Biosci Biotechnol. Biochem. 1999, 63, 1787–1790. [Google Scholar] [CrossRef]
- Corni, G.; Brighenti, V.; Pellati, F.; Morlock, G.E. Effect-direct analysis of bioactif compounds in Cannabis Sativa, L. by high-performance thin-layer chromatography. J. Chromatogr. A 2020, 1629, 461511. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Morton, D.W. Hyphenated TCL as a Tool in the Effect-Directed Discovery of Bioactive Natural Products. Appl. Sci. 2020, 10, 1123. [Google Scholar] [CrossRef] [Green Version]
- Nitta, Y.; Kikuzaki, H.; Azuma, T.; Ye, Y.; Sakaue, M.; Higuchi, Y.; Komori, H.; Ueno, H. Inhibitory activity of Filipendula ulmaria constituents on recombinant human histidine decarboxylase. Food. Chem. 2013, 138, 1551–1556. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Fruglova, M.Y. A new quercetin glycoside and other phenolics compounds from the genus Filipendula. Chem. Nat. Comp. 2013, 49, 610–616. [Google Scholar] [CrossRef]
- Samardžić, S.; Arsenijević, J.; Božić, D.; Milenković, M.; Tešević, V.; Maksimović, Z. Antioxidant, anti-inflammatory and gastroprotective activity of Filipendula ulmaria (L.) Maxim. and Filipendula vulgaris Moench. J. Ethnopharmacol. 2018, 213, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Sowndhararajan, K.; Joseph, J.M.; Rajendrakumaran, D. In vitro xanthine oxidase inhibitory activity of methanol extracts of Erythrina indica Lam. leaves and stem bark. Asian Pac. J. Trop. Biomed. 2012, 2, S1415–S1417. [Google Scholar] [CrossRef]
- Unno, T.; Sugimoto, A.; Kakuda, T. Xanthine oxidase inhibitors from the leaves of Lagerstroemia speciosa (L.) Pers. J. Ethnopharmacol. 2004, 93, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Arti, N.; Enkhtaivan, G.; Gyun-Seok, P.; Doo-Hwan, K.; Shivraj Hariram, N. Novel insights of multi-functional properties of flavonol glucosides from red onion (Allium cepa L.) solid waste—In vitro and in silico approach. Food. Chem. 2021, 335, 127650. [Google Scholar]
- Yuan, M.; Liu, Y.; Xiao, A.; Leng, J.; Liao, L.; Ma, L.; Liu, L. The interaction of dietary flavonoids with xanthine oxidase in vitro: Molecular property-binding affinity relationship aspects. RSC Adv. 2019, 9, 10781–10788. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, R.; Zhang, G.; Gong, D. Mechanistic insights into the inhibition of quercetin on xanthine oxidase. Int. J. Biol. Macromol. 2018, 112, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Masuoka, N.; Nihei, K.I.; Masuoka, T.; Kuroda, K.; Sasaki, K.; Kubo, I. The inhibition of uric acid formation catalysed by xanthine oxidase properties of the alkyl caffeates and cardol. J. Food. Res. 2012, 1, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Ko, E.Y.; Kim, D.H.; Keum, Y.S. Screening of ferulic acid related compounds as inhibitors of xanthine oxidase and cyclooxygenase-2 with anti-inflammatory activity. Rev. Bras. Farmacogn. 2016, 26, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, G.; Pan, J.; Gong, D. Novel insights into the inhibitory mechanism of kaempferol on xanthine oxidase. J. Agric. Food. Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef]
- Hatano, T.; Yasuhara, T.; Yoshihara, R.; Agata, I.; Noro, T.; Okuda, T. Effects of interaction of tannins with co-existing substances. VII. Inhibitory effects of tannins and related polyphenols on xanthine oxydase. Chem. Pharm. Bull. 1990, 38, 1224–1229. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, G.; Hu, Y.; Ma, Y. Effect of luteolin on xanthine oxidase inhibition kinetics and interaction mechanism merging with docking simulation. Food. Chem. 2013, 141, 3766–3773. [Google Scholar] [CrossRef] [PubMed]
- Ramallo, I.A.; Furlan, R.L.E. A rapid TLC autographic method for the detection of xanthine oxidase inhibitors and superoxide scavengers. Phytochem. Anal. 2006, 17, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Hu, Y.J.; Xu, P.; Liang, W.Q.; Zhou, J.; Liu, P.G.; Cheng, L.; Pu, J.B. Screening of potential xanthine oxidase inhibitors in Gnaphalium hypoleucum DC. by immobilized metal affinity chromatography and ultrafiltration-ultra performance liquid chromatography-mass spectrometry. Molecules 2016, 21, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Rt (min) | Compound | Formula | M − Hexp (m/z) | MS2 Fragment | Reference |
|---|---|---|---|---|---|---|
| 1 | 3.84 | Quinic acid | C7H12O6 | 191.0549 | 191/85/192/127/93 | Standard |
| 2 | 6.81 | Citric acid | C6H8O7 | 191.0189 | 111/87/85/191/129 | Standard |
| 3 | 8.52 | Gallic acid | C7H6O5 | 169.0131 | 125/169/126/170/97 | Standard |
| 4 | 12.97 | Chlorogenic acid | C16H18O9 | 353.0882 | 191/353/85/161/179 | Standard |
| 5 | 12.73 | Rugosin B | C41H30O27 | 953.0912 | 301/275/249/765/909 | Bijttebier et al., 2016 |
| 6 | 13.10 | Catechin | C15H14O6 | 289.0717 | 289/245/109/125/203 | Standard |
| 7 | 13.38 | Tellimagrandin I | C34H26O22 | 785.0887 | 301/275/785/249/169 | Bijttebier et al., 2016 |
| 8 | 14.48 | Rugosin E | C75H54O48 | 860.0865c | 301/275/169/249/785 | Bijttebier et al., 2016 |
| 9 | 15.12 | Rugosin A | C48H34O31 | 1105.1011 | 301/275/166/1061/937 | Bijttebier et al., 2016 |
| 10 | 15.50 | Tellimagrandin II | C41H30O26 | 937.0958 | 301/275/937/169/249 | Bijttebier et al., 2016 |
| 11 | 15.86 | Rugosin D | C82H58O52 | 936.0930c | 301/169/275/451/767 | Standard |
| 12 | 17.01 | Rutoside | C27H30O16 | 609.1459 | 300/609/301/271/255 | Standard |
| 13 | 17.88 | Ellagic acid | C14H6O8 | 300.9987 | 301/302/229/257/283 | Standard |
| 14 | 18.16 | Isoquercitrin | C21H20O12 | 463.0885 | 300/463/301/271/255 | Standard |
| 15 | 18,37 | Quercetin-3-O-(2″-O-galloyl)-β-galactopyranoside | C28H24O16 | 615.0996 | 301/151/178/313/302 | Bijttebier et al., 2016 |
| 16 | 18.58 | Hyperoside | C21H20O12 | 463.0882 | 300/463/301/271/255 | Standard |
| 17 | 18.70 | Miquelianin | C21H18O13 | 477.0677 | 301/477/151/179/255 | Bijttebier et al., 2016 |
| 18 | 22.13 | Astragalin | C21H20O11 | 447.093 | 284/447/285/151/107 | Standard |
| 19 | 22.78 | Astragalin-2″-O-gallate | C28H24O15 | 599.1052 | 285/313/257/169/229 | Chen et al., 2018; Samardžić et al., 2018 |
| 20 | 23.11 | Isorhamnetin-O-hexoside | C22H22O12 | 477.1041 | 477/314/271/243/285 | Bijttebier et al., 2016 |
| 21 | 23.86 | Spiraeoside | C21H20O12 | 463.0880 | 301/151/300/463/178 | Standard |
| 22 | 25.38 | Kaempferol-4′-O-glucoside | C21H20O11 | 447.0934 | 447/284/285/151/448 | Bijttebier et al., 2016 |
| 23 | 30.72 | Salicylic acid | C7H6O3 | 137.0229 | 93/137/94/138/65 | Standard |
| 24 | 38.83 | Quercetin | C15H10O7 | 301.0351 | 301/151/179/121/107 | Standard |
| 25 | 40.46 | Kaempferol | C15H10O6 | 285.0418 | 285/286/257/185/229 | Standard |
| Compound | IC50 (µg/mL) | IC50 (µM) |
|---|---|---|
| Allopurinol (control) | 2.9 ± 0.1 | 17.2 ± 0.8 |
| Gallic acid | >300 | >50 |
| Salicylic acid | >300 | >50 |
| Rugosin D | 67.0 ± 1.1 | 35.7 ± 2.1 |
| Kaempferol | 3.7 ± 0.2 | 12.9 ± 0.6 |
| Quercetin | 1.07 ± 0.06 | 3.5 ± 0.2 |
| Hyperoside | >100 | >50 |
| Isoquercitrin | >100 | >50 |
| Spiraeoside | 0.31 ± 0.01 | 0.66 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gainche, M.; Ogeron, C.; Ripoche, I.; Senejoux, F.; Cholet, J.; Decombat, C.; Delort, L.; Berthon, J.-Y.; Saunier, E.; Caldefie Chezet, F.; et al. Xanthine Oxidase Inhibitors from Filipendula ulmaria (L.) Maxim. and Their Efficient Detections by HPTLC and HPLC Analyses. Molecules 2021, 26, 1939. https://doi.org/10.3390/molecules26071939
Gainche M, Ogeron C, Ripoche I, Senejoux F, Cholet J, Decombat C, Delort L, Berthon J-Y, Saunier E, Caldefie Chezet F, et al. Xanthine Oxidase Inhibitors from Filipendula ulmaria (L.) Maxim. and Their Efficient Detections by HPTLC and HPLC Analyses. Molecules. 2021; 26(7):1939. https://doi.org/10.3390/molecules26071939
Chicago/Turabian StyleGainche, Maël, Clémence Ogeron, Isabelle Ripoche, François Senejoux, Juliette Cholet, Caroline Decombat, Laetitia Delort, Jean-Yves Berthon, Etienne Saunier, Florence Caldefie Chezet, and et al. 2021. "Xanthine Oxidase Inhibitors from Filipendula ulmaria (L.) Maxim. and Their Efficient Detections by HPTLC and HPLC Analyses" Molecules 26, no. 7: 1939. https://doi.org/10.3390/molecules26071939