
Chemical Composition of Tomato Seed Flours, and Their Radical Scavenging, Anti-Inflammatory and Gut Microbiota Modulating Properties

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of the Tomato Seed Flour Extracts
2.2. Total Phenolic Content and Free Radical Scavenging Capacities of Tomato Seed Flour Extracts
2.3. Anti-Inflammatory Capacities of Tomato Seed Flour Extracts
2.4. Gut Microbiota Profile Modulation of Tomato Seed Flour Extracts
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Tomato Seed Flour Extract Preparation
3.3. Ultra-High-Performance Liquid Chromatography-High Resolution Mass Spectrometry (UHPLC-HRMS) Analysis
3.4. Total Phenolic Content
3.5. Relative 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Capacity
3.6. Oxygen Radical Absorbing Capacity (ORAC)
3.7. 2,2′-azinobis (3-ethylbenzothiazoline-6-sulphonic Acid) Diammonium Salt Cation Radical (ABTS•+) Scavenging Capacity
3.8. Anti-Inflammatory Capacity
3.9. Gut Microbiota Analysis
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food Consumption and Nutrient Intakes. Available online: http://www.ers.usda.gov/data-products/food-consumption-and-nutrient-intakes.aspx (accessed on 6 January 2021).
- The World’s TOP40 Biggest Processing Companies. Available online: http://www.tomatonews.com/en/the-worlds-top40-biggest-processing-companies_2_355.html (accessed on 6 January 2021).
- Zuorro, A.; Lavecchia, R.; Medici, F.; Piga, L. Enzyme-Assisted Production of Tomato Seed Oil Enriched with Lycopene from Tomato Pomace. Food Bioprocess Technol. 2013, 6, 3499–3509. [Google Scholar] [CrossRef]
- Elbadrawy, E.; Sello, A. Evaluation of Nutritional Value and Antioxidant Activity of Tomato Peel Extracts. Arab. J. Chem. 2016, 9, S1010–S1018. [Google Scholar] [CrossRef] [Green Version]
- Shao, D.; Bartley, G.E.; Yokoyama, W.; Pan, Z.; Zhang, H.; Zhang, A. Plasma and Hepatic Cholesterol-Lowering Effects of Tomato Pomace, Tomato Seed Oil and Defatted Tomato Seed in Hamsters Fed with High-Fat Diets. Food Chem. 2013, 139, 589–596. [Google Scholar] [CrossRef]
- Chhonker, Y.S.; Pandey, C.P.; Chandasana, H.; Laxman, T.S.; Prasad, Y.D.; Narain, V.S.; Dikshit, M.; Bhatta, R.S. Simultaneous Quantitation of Acetylsalicylic Acid and Clopidogrel along with Their Metabolites in Human Plasma Using Liquid Chromatography Tandem Mass Spectrometry. Biomed. Chromatogr. 2016, 30, 466–473. [Google Scholar] [CrossRef]
- Vial, T.; Tan, W.-L.; Xiang, B.W.W.; Missé, D.; Deharo, E.; Marti, G.; Pompon, J. Dengue Virus Reduces AGPAT1 Expression to Alter Phospholipids and Enhance Infection in Aedes Aegypti. PLoS Pathog. 2019, 15, e1008199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, V.; Vicent, C.; Cerezo, M.; Mauch-Mani, B.; Dean, J.; Flors, V. Detection, Characterization and Quantification of Salicylic Acid Conjugates in Plant Extracts by ESI Tandem Mass Spectrometric Techniques. Plant Physiol. Biochem. 2012, 53, 19–26. [Google Scholar] [CrossRef]
- Al Kadhi, O.K.; Melchini, A.; Mithen, R.; Saha, S. Development of a LC-MS/MS Method for the Simultaneous Detection of Tricarboxylic Acid Cycle Intermediates in a Range of Biological Matrices. J. Anal. Methods Chem. 2017, 2017, 5391832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Jiang, J.; Duan, J.; Xie, Y.; Wang, J.; Zhai, S. Development and Validation of a Sensitive Liquid Chromatography-Tandem Mass Spectrometry Method for the Determination of Naringin and Its Metabolite, Naringenin, in Human Plasma. J. Chromatogr. Sci. 2014, 52, 654–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, R.M.; El-Halawany, A.M.; Saleh, D.O.; Naggar, E.M.B.E.; El-Shabrawy, A.E.-R.O.; El-Hawary, S.S. HPLC-DAD-MS/MS Profiling of Phenolics from Securigera Securidaca Flowers and Its Anti-Hyperglycemic and Anti-Hyperlipidemic Activities. Rev. Bras. Farmacogn. 2015, 25, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Comparative Metabolic Phenotyping of Tomato (Solanum lycopersicum) for the Identification of Metabolic Signatures in Cultivars Differing in Resistance to Ralstonia Solanacearum. Int. J. Mol. Sci. 2018, 19, 2558. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic Profiling of the Host Response of Tomato (Solanum Lycopersicum) Following Infection by Ralstonia Solanacearum. Int. J. Mol. Sci. 2019, 20, 3945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Meulebroek, L.; Bussche, J.V.; Steppe, K.; Vanhaecke, L. Ultra-High Performance Liquid Chromatography Coupled to High Resolution Orbitrap Mass Spectrometry for Metabolomic Profiling of the Endogenous Phytohormonal Status of the Tomato Plant. J. Chromatogr. A 2012, 1260, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Hvattum, E.; Ekeberg, D. Study of the Collision-Induced Radical Cleavage of Flavonoid Glycosides Using Negative Electrospray Ionization Tandem Quadrupole Mass Spectrometry. J. Mass Spectrom. 2003, 38, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Francescato, L.N.; Debenedetti, S.L.; Schwanz, T.G.; Bassani, V.L.; Henriques, A.T. Identification of Phenolic Compounds in Equisetum Giganteum by LC–ESI-MS/MS and a New Approach to Total Flavonoid Quantification. Talanta 2013, 105, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Horikawa, M.; Sato, S.; Miyake, H.; Setou, M. Discovery of Lipid Biomarkers Correlated with Disease Progression in Clear Cell Renal Cell Carcinoma Using Desorption Electrospray Ionization Imaging Mass Spectrometry. Oncotarget 2019, 10, 1688–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calixto, N.O.; Cordeiro, M.S.; Giorno, T.B.S.; Oliveira, G.G.; Lopes, N.P.; Fernandes, P.D.; Pinto, A.C.; Rezende, C.M. Chemical Constituents of Psychotria Nemorosa Gardner and Antinociceptive Activity. J. Braz. Chem. Soc. 2017, 28, 707–723. [Google Scholar] [CrossRef]
- Oliviero, F.; Scanu, A.; Zamudio-Cuevas, Y.; Punzi, L.; Spinella, P. Anti-Inflammatory Effects of Polyphenols in Arthritis. J. Sci. Food Agric. 2018, 98, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Fenercioglu, A.K.; Saler, T.; Genc, E.; Sabuncu, H.; Altuntas, Y. The Effects of Polyphenol-Containing Antioxidants on Oxidative Stress and Lipid Peroxidation in Type 2 Diabetes Mellitus without Complications. J. Endocrinol. Investig. 2010, 33, 118–124. [Google Scholar] [CrossRef]
- Bose, M.; Lambert, J.D.; Ju, J.; Reuhl, K.R.; Shapses, S.A.; Yang, C.S. The Major Green Tea Polyphenol, (-)-Epigallocatechin-3-Gallate, Inhibits Obesity, Metabolic Syndrome, and Fatty Liver Disease in High-Fat-Fed Mice. J. Nutr. 2008, 138, 1677–1683. [Google Scholar] [CrossRef]
- Quiñones, M.; Miguel, M.; Aleixandre, A. Beneficial Effects of Polyphenols on Cardiovascular Disease. Pharmacol. Res. 2013, 68, 125–131. [Google Scholar] [CrossRef]
- Paterson, J.R.; Lawrence, J.R. Salicylic Acid: A Link between Aspirin, Diet and the Prevention of Colorectal Cancer. QJM Int. J. Med. 2001, 94, 445–448. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant Properties, Radical Scavenging Activity and Biomolecule Protection Capacity of Flavonoid Naringenin and Its Glycoside Naringin: A Comparative Study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef]
- Bugianesi, R.; Catasta, G.; Spigno, P.; D’Uva, A.; Maiani, G. Naringenin from Cooked Tomato Paste Is Bioavailable in Men. J. Nutr. 2002, 132, 3349–3352. [Google Scholar] [CrossRef]
- Jan, A.T.; Kamli, M.R.; Murtaza, I.; Singh, J.B.; Ali, A.; Haq, Q.M.R. Dietary Flavonoid Quercetin and Associated Health Benefits—An Overview. Food Rev. Int. 2010, 26, 302–317. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A Review of the Dietary Flavonoid, Kaempferol on Human Health and Cancer Chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, Antioxidant and Anti-Inflammatory Activities of Kaempferol and Its Corresponding Glycosides and the Enzymatic Preparation of Kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and Metabolism of Polyphenols in the Gut and Impact on Health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Suzuki, T.; Clydesdale, F.M.; Pandolf, T. Solubility of Iron in Model Systems Containing Organic Acids and Lignin. J. Food Prot. 1992, 55, 893–898. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int. J. Tryptophan Res. 2018, 11, 1178646918802282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Anzai, M.; Inui, M.; Nishimoto, T.; Inoue, G. Utilization of N-acetyl-L-tryptophan given intravenously to unrestrained adult rats. J. Nutr. Sci. Vitaminol. 1980, 26, 381–388. [Google Scholar] [CrossRef]
- Berg, C.P.; Rose, W.C.; Marvel, C.S. Tryptophane and growth: III. 3-indolepropionic acid and 3-indolepyruvic acid as supplementing agents in diets deficient in tryptophane. J. Biol. Chem. 1929, 85, 219–231. [Google Scholar] [CrossRef]
- Jones, D.A. Rosacea, Reactive Oxygen Species, and Azelaic Acid. J. Clin. Aesthet. Dermatol. 2009, 2, 26–30. [Google Scholar] [PubMed]
- Toor, R.K.; Savage, G.P. Antioxidant Activity in Different Fractions of Tomatoes. Food Res. Int. 2005, 38, 487–494. [Google Scholar] [CrossRef]
- Kaur, C.; Kapoor, H.C. Anti-Oxidant Activity and Total Phenolic Content of Some Asian Vegetables. Int. J. Food Sci. Technol. 2002, 37, 153–161. [Google Scholar] [CrossRef]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and Pitfalls in Measuring Antioxidant Efficacy: A Critical Evaluation of ABTS, DPPH, and ORAC Assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Zhou, K.; Yu, L. Total Phenolic Contents and Antioxidant Properties of Commonly Consumed Vegetables Grown in Colorado. LWT Food Sci. Technol. 2006, 39, 1155–1162. [Google Scholar] [CrossRef]
- Perea-Domínguez, X.P.; Hernández-Gastelum, L.Z.; Olivas-Olguin, H.R.; Espinosa-Alonso, L.G.; Valdez-Morales, M.; Medina-Godoy, S. Phenolic Composition of Tomato Varieties and an Industrial Tomato By-Product: Free, Conjugated and Bound Phenolics and Antioxidant Activity. J. Food Sci. Technol. 2018, 55, 3453–3461. [Google Scholar] [CrossRef]
- Navarrete, S.; Alarcón, M.; Palomo, I. Aqueous Extract of Tomato (Solanum lycopersicum L.) and Ferulic Acid Reduce the Expression of TNF-α and IL-1β in LPS-Activated Macrophages. Molecules 2015, 20, 15319–15329. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadioglu, O.; Nass, J.; Saeed, M.E.M.; Schuler, B.; Efferth, T. Kaempferol Is an Anti-Inflammatory Compound with Activity towards NF-ΚB Pathway Proteins. Anticancer Res. 2015, 35, 2645–2650. [Google Scholar] [PubMed]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Gálvez, J.; Zarzuelo, A. In Vivo Quercitrin Anti-Inflammatory Effect Involves Release of Quercetin, Which Inhibits Inflammation through down-Regulation of the NF-ΚB Pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [Green Version]
- Choe, U.; Li, Y.; Gao, B.; Yu, L.; Wang, T.T.Y.; Sun, J.; Chen, P.; Liu, J.; Yu, L. Chemical Compositions of Cold-Pressed Broccoli, Carrot, and Cucumber Seed Flours and Their in Vitro Gut Microbiota Modulatory, Anti-Inflammatory, and Free Radical Scavenging Properties. J. Agric. Food Chem. 2018, 66, 9309–9317. [Google Scholar] [CrossRef]
- Kim, Y.S.; Milner, J.A. Dietary Modulation of Colon Cancer Risk. J. Nutr. 2007, 137, 2576S–2579S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.J.; Rocha, E.R.; Paster, B.J. The Medically Important Bacteroides spp. in Health and Disease. In The Prokaryotes: Volume 7: Proteobacteria: Delta, Epsilon Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 381–427. ISBN 978-0-387-30747-3. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review Article: Bifidobacteria as Probiotic Agents-Physiological Effects and Clinical Benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia Muciniphila and Improved Metabolic Health during a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut 2016, 65, 426. [Google Scholar] [CrossRef] [Green Version]
- Amar, J.; Burcelin, R.; Ruidavets, J.B.; Cani, P.D.; Fauvel, J.; Alessi, M.C.; Chamontin, B.; Ferriéres, J. Energy Intake Is Associated with Endotoxemia in Apparently Healthy Men. Am. J. Clin. Nutr. 2008, 87, 1219–1223. [Google Scholar] [CrossRef] [Green Version]
- Precup, G.; Vodnar, D.-C. Gut Prevotella as a Possible Biomarker of Diet and Its Eubiotic versus Dysbiotic Roles: A Comprehensive Literature Review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef] [PubMed]
- la Reau, A.J.; Suen, G. The Ruminococci: Key Symbionts of the Gut Ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhou, K. Antioxidant Properties of Bran Extracts from ‘Platte’ Wheat Grown at Different Locations. Food Chem. 2005, 90, 311–316. [Google Scholar] [CrossRef]
- Choe, U.; Li, Y.; Gao, B.; Yu, L.; Wang, T.T.Y.; Sun, J.; Chen, P.; Yu, L. The Chemical Composition of a Cold-Pressed Milk Thistle Seed Flour Extract, and Its Potential Health Beneficial Properties. Food Funct. 2019, 10, 2461–2470. [Google Scholar] [CrossRef]
- Cheng, Z.; Moore, J.; Yu, L. High-Throughput Relative DPPH Radical Scavenging Capacity Assay. J. Agric. Food Chem. 2006, 54, 7429–7436. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.A. Factors Influencing the Antioxidant Activity Determined by the ABTS•+ Radical Cation Assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]





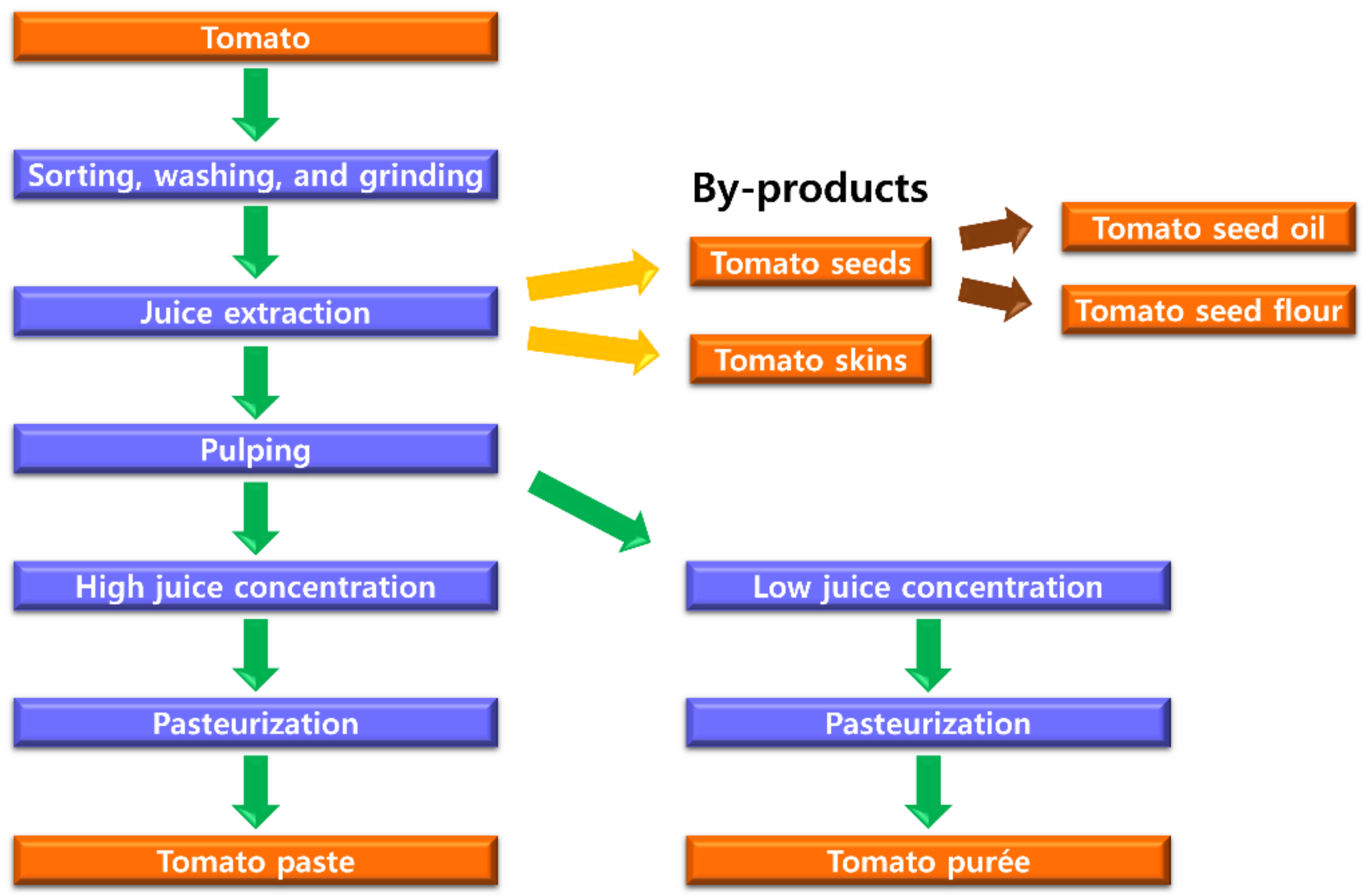{kind=link}
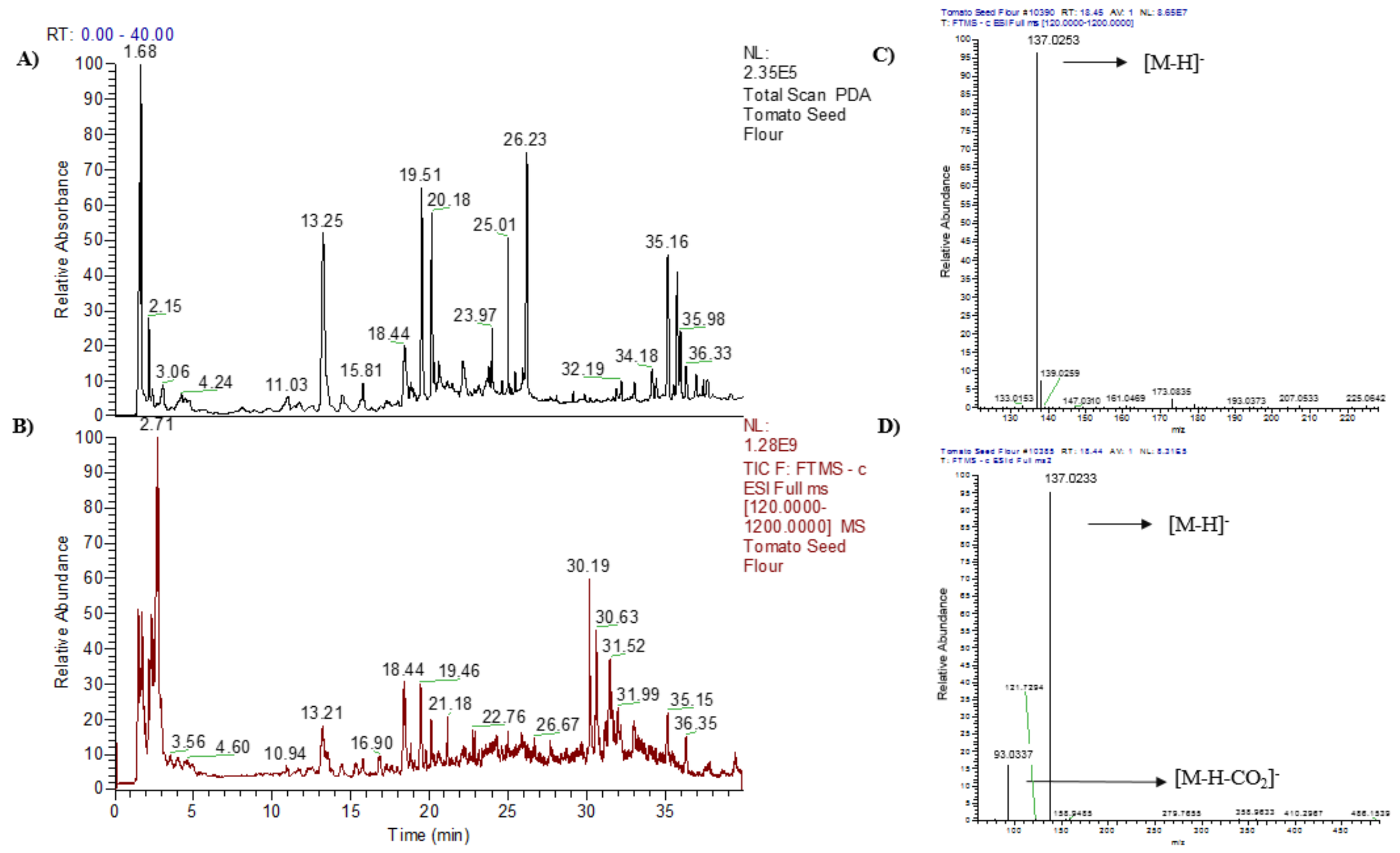{kind=link}
{kind=link}
{kind=link}
| ID | Rt (min) | Exptl. [M–H]− | Fragment Ions | Formula | Mass Error (mmμ) | Tentative Identification | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 1.68 | 133.0135 | 71.0131 | C4H6O5 | −0.75 | Malic acid | [6] |
| 2 | 4.95 | 161.0448 | 323.097 ([2M–H]−) 143.0343 ([M–H2O]−) | C6H10O5 | −0.75 | 2-Hydroxyadipic acid | [7] |
| 3 | 18.44 | 137.0237 | 93.0337 ([M–H–CO2]−) | C7H6O3 | −0.72 | Salicylic acid | [8,9] |
| 4 | 18.81 | 579.1708 | 271.0595 ([narigenin–H]−) | C27H32O14 | −1.13 | Naringin | [10,11] |
| 5 | 19.46 | 245.0919 | 491.1914 ([2M–H]−) | C13H14N2O3 | −1.27 | N-Acetyl-tryptophan | [12,13,14] |
| 6 | 19.48 | 625.1381 | 300.0259 ([quercetin–H]−•) 301.0337 ([quercetin–H]−) | C27H30O17 | −2.92 | Quercetin-di-O-hexoside | [15,16] |
| 7 | 20.14 | 609.1426 | 284.0310 ([kaempferol–H]−•) 285.0389 ([kaempferol–H]−) | C27H30O16 | −3.51 | Kaempferol-di-O-hexoside | [16] |
| 8 | 21.17 | 187.0991 | 375.1929 ([2M–H]−) | C9H16O4 | 1.52 | Azelaic acid | [17,18] |
| Sample | TPC | ORAC | DPPH | ABTS |
|---|---|---|---|---|
| mg GAE/g | µmoles TE/g | µmoles TE/g | µmoles TE/g | |
| TSF1 | 2.00 ± 0.11 | 88.57 ± 2.42 | 3.57 ± 0.09 | 3.39 ± 0.08 |
| TSF2 | 1.97 ± 0.30 | 86.32 ± 7.01 | 3.81 ± 0.20 | 3.58 ± 0.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, U.; Sun, J.; Bailoni, E.; Chen, P.; Li, Y.; Gao, B.; Wang, T.T.Y.; Rao, J.; Yu, L. Chemical Composition of Tomato Seed Flours, and Their Radical Scavenging, Anti-Inflammatory and Gut Microbiota Modulating Properties. Molecules 2021, 26, 1478. https://doi.org/10.3390/molecules26051478
Choe U, Sun J, Bailoni E, Chen P, Li Y, Gao B, Wang TTY, Rao J, Yu L. Chemical Composition of Tomato Seed Flours, and Their Radical Scavenging, Anti-Inflammatory and Gut Microbiota Modulating Properties. Molecules. 2021; 26(5):1478. https://doi.org/10.3390/molecules26051478
Chicago/Turabian StyleChoe, Uyory, Jianghao Sun, Elena Bailoni, Pei Chen, Yanfang Li, Boyan Gao, Thomas T. Y. Wang, Jiajia Rao, and Liangli (Lucy) Yu. 2021. "Chemical Composition of Tomato Seed Flours, and Their Radical Scavenging, Anti-Inflammatory and Gut Microbiota Modulating Properties" Molecules 26, no. 5: 1478. https://doi.org/10.3390/molecules26051478