
Physiological and Molecular Responses to Acid Rain Stress in Plants and the Impact of Melatonin, Glutathione and Silicon in the Amendment of Plant Acid Rain Stress

 ,
,  ,
,
Abstract
:1. Introduction
2. Impacts of Acid Rain on the Photosynthesis Potential in Plants
3. Reactive Oxygen Species (ROS) Stress and Its Metabolic Mechanism
4. Mechanism of Acid Rain in Related Gene Expression in Plants
5. Impact of Silicon in Plants under Acid Rain Stress Conditions
6. Effect of Glutathione in Plants under Acid Rain Stress
7. Role of Melatonin in Plants under Acid Rain Stress
8. Conclusions and Future Prospective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dwivedi, A.; Tripathi, B. Pollution tolerance and distribution pattern of plants in surrounding area of coal-fired industries. J. Environ. Biol. 2007, 28, 257–263. [Google Scholar]
- Bouwman, A.F.; Vuuren, D.P.V.; Derwent, R.G.; Posch, M. A global analysis of acidification and eutrophication of terrestrial ecosystems. Water Air Soil Pollut. 2002, 141, 349–382. [Google Scholar] [CrossRef]
- Feng, Z. Impacts and control strategies of acid deposition on terrestrial ecosystems in china. Eng. Sci. 2000, 9, 5–11. [Google Scholar]
- Chen, J.; Li, W.; Gao, F. Biogeochemical effects of forest vegetation on acid precipitation-related water chemistry: A case study in southwest china. J. Environ. Monit. 2010, 12, 1799–1806. [Google Scholar] [CrossRef]
- Feng, Z. Ecological effects and control strategies of acid deposition on ecosystems in china. Yunnan Environ. Sci. 2000, 19, 1–6. [Google Scholar]
- Fairfax, J.; Lepp, N. Effect of simulated ‘acid rain’on cation loss from leaves. Nature 1975, 255, 324. [Google Scholar] [CrossRef]
- Seinfeld, J.H.; Pandis, S.N. Atmospheric Chemistry and Physics: From Air Pollution to Climate Change; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Ren, X.; Zhu, J.; Liu, H.; Xu, X.; Liang, C. Response of antioxidative system in rice (Oryza sativa) leaves to simulated acid rain stress. Ecotoxicol. Environ. Saf. 2018, 148, 851–856. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, G.; Yang, C.; Ou, Z.; Peng, C. Responses of chlorophyll fluorescence and xanthophyll cycle in leaves of Schima superba Gardn. & Champ. and Pinus massoniana Lamb. to simulated acid rain at dinghushan biosphere reserve, china. Acta Physiol. Plant. 2007, 29, 33–38. [Google Scholar]
- Liu, T.-W.; Wu, F.-H.; Wang, W.-H.; Chen, J.; Li, Z.-J.; Dong, X.-J.; Patton, J.; Pei, Z.-M.; Zheng, H.-L. Effects of calcium on seed germination, seedling growth and photosynthesis of six forest tree species under simulated acid rain. Tree Physiol. 2011, 31, 402–413. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Sanavy, S.A.M.M.; Gholamhoseini, M.; Joghan, A.K.; Majdi, M.; Kashkooli, A.B. The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol. Mol. Biol. Plants 2013, 19, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, W.-H.; Liu, T.-W.; Wu, F.-H.; Zheng, H.-L. Photosynthetic and antioxidant responses of Liquidambar formosana and Schima superba seedlings to sulfuric-rich and nitric-rich simulated acid rain. Plant Physiol. Biochem. 2013, 64, 41–51. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Bačkor, M.; Stork, F.; Hedbavny, J. Physiological responses of root-less epiphytic plants to acid rain. Ecotoxicology 2011, 20, 348–357. [Google Scholar] [CrossRef]
- Wen, K.; Liang, C.; Wang, L.; Hu, G.; Zhou, Q. Combined effects of lanthanumion and acid rain on growth, photosynthesis and chloroplast ultrastructure in soybean seedlings. Chemosphere 2011, 84, 601–608. [Google Scholar] [CrossRef]
- Liu, T.W.; Niu, L.; Fu, B.; Chen, J.; Wu, F.H.; Chen, J.; Wang, W.H.; Hu, W.J.; He, J.X.; Zheng, H.L. A transcriptomic study reveals differentially expressed genes and pathways respond to simulated acid rain in Arabidopsis thaliana. Genome 2013, 56, 49–60. [Google Scholar] [CrossRef]
- Ju, S.; Wang, L.; Chen, J. Effects of silicon on the growth, photosynthesis and chloroplast ultrastructure of Oryza sativa L. seedlings under acid rain stress. Silicon 2020, 12, 655–664. [Google Scholar] [CrossRef]
- Xalxo, R.; Keshavkant, S. Melatonin, glutathione and thiourea attenuates lead and acid rain-induced deleterious responses by regulating gene expression of antioxidants in Trigonella foenum graecum L. Chemosphere 2019, 221, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Li, M.; Liu, S.; Pan, T.; Ma, C.; Qiu, D. Melatonin-mediate acid rain stress tolerance mechanism through alteration of transcriptional factors and secondary metabolites gene expression in tomato. Ecotoxicol. Environ. Saf. 2020, 200, 110720. [Google Scholar] [CrossRef] [PubMed]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s role in abiotic and biotic plant stresses. Annu. Rev. Phytop. 2017, 55, 85–107. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, T.; Calomme, M.; Borowiak, K. The effect of choline-stabilized orthosilicic acid on microelements and silicon concentration, photosynthesis activity and yield of tomato grown under Mn stress. Plant Physiol. Biochem. 2015, 96, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, A.A.; Rodrigues, F.Á.; do Nascimento, K.J.T. Physiological and biochemical aspects of the resistance of banana plants to fusarium wilt potentiated by silicon. Phytopathology 2012, 102, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Resende, R.; Rodrigues, F.; Gomes, R.; Nascimento, K. Microscopic and biochemical aspects of sorghum resistance to anthracnose mediated by silicon. Ann. Appl. Biol. 2013, 163, 114–123. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Alharby, H.F.; Fujita, M. Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J. Plant. Inter. 2018, 13, 203–212. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant. Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Rodrigo, M.A.M.; Anjum, N.A.; Heger, Z.; Zitka, O.; Vojtech, A.; Pereira, E.; Kizek, R. Role of phytochelatins in redox caused stress in plants and animals. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; IntechOpen: London, UK, 2016. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Hossain, M.; Mahmud, J.A.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Deng, S.-L.; Sun, T.-C.; Li, Y.-Y.; Liu, Y.-X. Melatonin regulates the synthesis of steroid hormones on male reproduction: A review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.X.; Xu, B.; Zhou, X.; Reiter, R.J. Pineal calcification, melatonin production, aging, associated health consequences and rejuvenation of the pineal gland. Molecules 2018, 23, 301. [Google Scholar] [CrossRef] [Green Version]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Beilby, M.J.; Turi, C.E.; Baker, T.C.; Tymm, F.J.; Murch, S.J. Circadian changes in endogenous concentrations of indole-3-acetic acid, melatonin, serotonin, abscisic acid and jasmonic acid in characeae (Chara australis Brown). Plant. Signal. Behav. 2015, 10, e1082697. [Google Scholar] [CrossRef] [Green Version]
- Shiu, S.Y.; Pang, B.; Tam, C.W.; Yao, K.M. Signal transduction of receptor-mediated antiproliferative action of melatonin on human prostate epithelial cells involves dual activation of gαs and gαq proteins. J. Pineal Res. 2010, 49, 301–311. [Google Scholar] [CrossRef]
- Imbesi, M.; Arslan, A.D.; Yildiz, S.; Sharma, R.; Gavin, D.; Tun, N.; Manev, H.; Uz, T. The melatonin receptor mt1 is required for the differential regulatory actions of melatonin on neuronal ‘clock’gene expression in striatal neurons in vitro. J. Pineal Res. 2009, 46, 87–94. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, M.; Hardeland, R. The melatonin metabolite n1-acetyl-5-methoxykynuramine is a potent singlet oxygen scavenger. J. Pineal Res. 2009, 46, 49–52. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Martín, M.; Macías, M.; Escames, G.; León, J.; Khaldy, H.; Reiter, R.J. Melatonin, mitochondria, and cellular bioenergetics. J. Pineal Res. 2001, 30, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nopparat, C.; Porter, J.E.; Ebadi, M.; Govitrapong, P. The mechanism for the neuroprotective effect of melatonin against methamphetamine-induced autophagy. J. Pineal Res. 2010, 49, 382–389. [Google Scholar] [CrossRef]
- Um, H.J.; Kwon, T.K. Protective effect of melatonin on oxaliplatin induced apoptosis through sustained mcl-1 expression and antioxidant action in renal Carcinoma caki cells. J. Pineal Res. 2010, 49, 283–290. [Google Scholar] [CrossRef]
- Xu, S.C.; He, M.D.; Zhong, M.; Zhang, Y.W.; Wang, Y.; Yang, L.; Yang, J.; Yu, Z.P.; Zhou, Z. Melatonin protects against nickel induced neurotoxicity in vitro by reducing oxidative stress and maintaining mitochondrial function. J. Pineal Res. 2010, 49, 86–94. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. 2010, 85, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-Z.; Deng, X.-P.; Xu, B.-C.; Gao, Z.-J.; Ding, W.-L. Photosynthetic activity and efficiency of Bothriochloa ischaemum and Lespedeza davurica in mixtures across growth periods under water stress. Acta Physiol. Plant. 2014, 36, 1033–1044. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Song, H.; Chen, J.; Long, Y. Antagonism or synergism? Combined effects of enhanced uv-b radiation and acid rain on photosynthesis in seedlings of two c4 plants. Acta Ecol. Sin. 2020, 40, 72–80. [Google Scholar] [CrossRef]
- Zhang, C.; Yi, X.; Gao, X.; Wang, M.; Shao, C.; Lv, Z.; Chen, J.; Liu, Z.; Shen, C. Physiological and biochemical responses of tea seedlings (Camellia sinensis) to simulated acid rain conditions. Ecotoxicol. Environ. Saf. 2020, 192, 110315. [Google Scholar] [CrossRef]
- Reza Yousefi, A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and seedling growth responses of zygophyllum fabago, salsola kali l. And atriplex canescens to peg-induced drought stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Zuo, Z.; Wang, B.; Ying, B.; Zhou, L.; Zhang, R. Monoterpene emissions contribute to thermotolerance in Cinnamomum camphora. Trees 2017, 31, 1759–1771. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2011, 63, 577–597. [Google Scholar] [CrossRef]
- Gao, P.; Zuo, Z.; Wu, X.; Gao, Y.; Gao, R.; Zhang, R. Effects of cycloheximide on photosynthetic abilities, reflectance spectra and fluorescence emission spectra in Phyllostachys edulis. Trees 2016, 30, 719–732. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.Y.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Reiter, R.J. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J. Pineal Res. 2016, 61, 457–469. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous melatonin ameliorates cold-induced damage in tomato plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, H.; Cao, K.; Hu, L.; Du, T.; Baluška, F.; Zou, Z. Beneficial roles of melatonin on redox regulation of photosynthetic electron transport and synthesis of d1 protein in tomato seedlings under salt stress. Front. Plant. Sci. 2016, 7, 1823. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Jin, Z.; Wang, S.; Gong, B.; Wen, D.; Wang, X.; Wei, M.; Shi, Q. Sodic alkaline stress mitigation with exogenous melatonin involves reactive oxygen metabolism and ion homeostasis in tomato. Sci. Hortic. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous melatonin improves seedling health index and drought tolerance in tomato. Plant. Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Hasan, M.K.; Ahammed, G.J.; Yin, L.; Shi, K.; Xia, X.; Zhou, Y.; Yu, J.; Zhou, J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front. Plant. Sci. 2015, 6, 601. [Google Scholar] [CrossRef]
- Hu, H.; Wang, L.; Liao, C.; Fan, C.; Zhou, Q.; Huang, X. Combined effects of lead and acid rain on photosynthesis in soybean seedlings. Biol. Trace Elem. Res. 2014, 161, 136–142. [Google Scholar] [CrossRef]
- Debnath, B.; Ahammed, G.J. Effect of acid rain on plant growth and development: Physiological and molecular interventions. In Contaminants in Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 103–114. [Google Scholar]
- Debnath, B.; Hussain, M.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Qiu, D. Exogenous melatonin mitigates acid rain stress to tomato plants through modulation of leaf ultrastructure, photosynthesis and antioxidant potential. Molecules 2018, 23, 388. [Google Scholar] [CrossRef] [Green Version]
- Debnath, B.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Rizwan, H.M.; Pan, T.; Qiu, D. Acid rain deposition modulates photosynthesis, enzymatic and non-enzymatic antioxidant activities in tomato. Int. J. Environ. Res. 2018, 12, 203–214. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Xu, B.; Li, J.; Huang, B. Exogenous melatonin suppresses dark-induced leaf senescence by activating the superoxide dismutase-catalase antioxidant pathway and down-regulating chlorophyll degradation in excised leaves of perennial ryegrass (Lolium perenne L.). Front. Plant. Sci. 2016, 7, 1500. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, T.; Ding, F. Exogenous melatonin delays methyl jasmonate-triggered senescence in tomato leaves. Agronomy 2019, 9, 795. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Korpelainen, H.; Dong, L.; Yi, L. Physiological responses of Elaeocarpus glabripetalus seedlings exposed to simulated acid rain and cadmium. Ecotoxicol. Environ. Saf. 2019, 175, 118–127. [Google Scholar] [CrossRef]
- Kummerová, M.; Krulová, J.; Zezulka, Š.; Tříska, J. Evaluation of fluoranthene phytotoxicity in pea plants by hill reaction and chlorophyll fluorescence. Chemosphere 2006, 65, 489–496. [Google Scholar] [CrossRef]
- Zhang, F.; Cheng, M.; Sun, Z.; Wang, L.; Zhou, Q.; Huang, X. Combined acid rain and lanthanum pollution and its potential ecological risk for nitrogen assimilation in soybean seedling roots. Environ. Pollut. 2017, 231, 524–532. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutzendubel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Di Toppi, L.S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen peroxide in plants: A versatile molecule of the reactive oxygen species network. J. Integr. Plant. Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. Ros generation in plants: Boon or bane? Plant. Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Bio. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Montillet, J.-L.; Chamnongpol, S.; Rustérucci, C.; Dat, J.; Van De Cotte, B.; Agnel, J.-P.; Battesti, C.; Inzé, D.; Van Breusegem, F.; Triantaphylidès, C. Fatty acid hydroperoxides and H2O2 in the execution of hypersensitive cell death in tobacco leaves. Plant. Physiol. 2005, 138, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Qiao, F.; Zhang, X.-M.; Liu, X.; Chen, J.; Hu, W.-J.; Liu, T.-W.; Liu, J.-Y.; Zhu, C.-Q.; Ghoto, K.; Zhu, X.-Y. Elevated nitrogen metabolism and nitric oxide production are involved in arabidopsis resistance to acid rain. Plant. Physiol. Biochem. 2018, 127, 238–247. [Google Scholar] [CrossRef]
- Xia, B.; Sun, Z.; Wang, L.; Zhou, Q.; Huang, X. Analysis of the combined effects of lanthanum and acid rain, and their mechanisms, on nitrate reductase transcription in plants. Ecotoxicol. Environ. Saf. 2017, 138, 170–178. [Google Scholar] [CrossRef]
- Ju, S.; Wang, L.; Yin, N.; Li, D.; Wang, Y.; Zhang, C. Silicon alleviates simulated acid rain stress of Oryza sativa L. seedlings by adjusting physiology activity and mineral nutrients. Protoplasma 2017, 254, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, H.; Zhong, Y.; Huang, J.; Fu, X.; Wang, L.; Teng, W. Growth and physiological response of an endangered tree, Horsfieldia hainanensis Merr. to simulated sulfuric and nitric acid rain in southern china. Plant. Physiol. Biochem. 2019, 144, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Fontenele, N.M.B.; Otoch, M.d.L.O.; Gomes-Rochette, N.F.; de Menezes Sobreira, A.C.; Barreto, A.A.G.C.; de Oliveira, F.D.B.; Costa, J.H.; Borges, S.d.S.S.; do Nascimento, R.F.; de Melo, D.F. Effect of lead on physiological and antioxidant responses in two Vigna unguiculata cultivars differing in pb-accumulation. Chemosphere 2017, 176, 397–404. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Chen, F.; Wang, F.; Wu, F.; Mao, W.; Zhang, G.; Zhou, M. Modulation of exogenous glutathione in antioxidant defense system against cd stress in the two barley genotypes differing in cd tolerance. Plant. Physiol. Biochem. 2010, 48, 663–672. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant. Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Jing, X.; Tang, H.; Li, X.; Gong, B.; Shi, Q. Using transcriptome to discover a novel melatonin-induced sodic alkaline stress resistant pathway in Solanum lycopersicum L. Plant. Cell Physiol. 2019, 60, 2051–2064. [Google Scholar] [CrossRef]
- Zheng, S.; Pan, T.; Ma, C.; Qiu, D. Differential gene expression of longan under simulated acid rain stress. B. Environ. Contam. Tox. 2017, 98, 726–731. [Google Scholar] [CrossRef]
- Liu, T.; Ma, J.; Li, M.; Pan, T.; Ma, C.; Qiu, D. Cdna-aflp analysis reveals inducible gene expression in tomato leaves in response to simulated acid rain. Appl. Ecol. Environ. Res. 2019, 17, 6515–6533. [Google Scholar] [CrossRef]
- Liu, T.-W.; Fu, B.; Niu, L.; Chen, J.; Wang, W.-H.; He, J.-X.; Pei, Z.-M.; Zheng, H.-L. Comparative proteomic analysis of proteins in response to simulated acid rain in arabidopsis. J. Proteome Res. 2011, 10, 2579–2589. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Wang, G.; Ji, J.; Shi, L.; Guan, C.; Jin, C. Comparative transcriptome analysis of transcription factors in different maize varieties under salt stress conditions. Plant. Growth Regul. 2017, 81, 183–195. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.; Gong, W.; Sun, J.; Pan, Z.; Xu, F.; Lu, Y.; Du, X. Comprehensive analysis of differentially expressed genes and transcriptional regulation induced by salt stress in two contrasting cotton genotypes. BMC Genom. 2014, 15, 760. [Google Scholar] [CrossRef] [Green Version]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of myb and bhlh plant transcription factors. Plant. J. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant. Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Ap2/erf family transcription factors in plant abiotic stress responses. BBA Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Song, A.; Li, Z.; Fan, F.; Liang, Y. Silicon ameliorates manganese toxicity by regulating both physiological processes and expression of genes associated with photosynthesis in rice (Oryza sativa L.). Plant. Soil 2015, 397, 289–301. [Google Scholar] [CrossRef]
- Liang, Y.; Hua, H.; Zhu, Y.G.; Zhang, J.; Cheng, C.; Römheld, V. Importance of plant species and external silicon concentration to active silicon uptake and transport. New Phytol. 2006, 172, 63–72. [Google Scholar] [CrossRef]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.; Sanglard, L.M.; Reis, J.V.; Detmann, E.; Rodrigues, F.Á.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Cooke, J.; Leishman, M.R. Consistent alleviation of abiotic stress with silicon addition: A meta-analysis. Funct. Ecol. 2016, 30, 1340–1357. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Naranjo, E.; Andrades-Moreno, L.; Davy, A.J. Silicon alleviates deleterious effects of high salinity on the halophytic grass Spartina densiflora. Plant. Physiol. Biochem. 2013, 63, 115–121. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Siti Nor Akmar, A.; Rafii, M.Y.; Azizi, P.; Tengoua, F.; Nurul Mayzaitul Azwa, J.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The effect of silicon on photosynthesis and expression of its relevant genes in rice (Oryza sativa L.) under high-zinc stress. PLoS ONE 2014, 9, e113782. [Google Scholar] [CrossRef]
- Velikova, V.; Tsonev, T.; Yordanov, I. Light and co2 responses of photosynthesis and chlorophyll fluorescence characteristics in bean plants after simulated acid rain. Physiol. Plant. 1999, 107, 77–83. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, S.; Gong, H.-j.; Zhao, H.-l.; Li, H.-l.; Hu, Y.-h.; Wang, Y.-c. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integr. Agr. 2018, 17, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.-Y.; Lin, W.-P.; Rui, L.; Jiang, N.-H.; Cai, K.-Z. Physiological response and phenolic metabolism in tomato (Solanum lycopersicum) mediated by silicon under Ralstonia solanacearum infection. J. Integr. Agr. 2018, 17, 2160–2171. [Google Scholar] [CrossRef] [Green Version]
- Silva, I.T.; Rodrigues, F.Á.; Oliveira, J.R.; Pereira, S.C.; Andrade, C.C.L.; Silveira, P.R.; Conceição, M.M. Wheat resistance to bacterial leaf streak mediated by silicon. J. Phytopathol. 2010, 158, 253–262. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Mohmood, I.; Pacheco, M.; Duarte, A.C.; Pereira, E.; Umar, S.; Ahmad, A.; Khan, N.A.; Iqbal, M. Modulation of glutathione and its related enzymes in plants’ responses to toxic metals and metalloids—A review. Environ. Exp. Bot. 2012, 75, 307–324. [Google Scholar] [CrossRef]
- Hasan, M.K.; Liu, C.; Wang, F.; Ahammed, G.J.; Zhou, J.; Xu, M.-X.; Yu, J.-Q.; Xia, X.-J. Glutathione-mediated regulation of nitric oxide, s-nitrosothiol and redox homeostasis confers cadmium tolerance by inducing transcription factors and stress response genes in tomato. Chemosphere 2016, 161, 536–545. [Google Scholar] [CrossRef]
- Teh, C.Y.; Mahmood, M.; Shaharuddin, N.A.; Ho, C.L. In vitro rice shoot apices as simple model to study the effect of nacl and the potential of exogenous proline and glutathione in mitigating salinity stress. Plant Growth Regul. 2015, 75, 771–781. [Google Scholar] [CrossRef]
- Xalxo, R.; Sahu, K. Acid rain-induced oxidative stress regulated metabolic interventions and their amelioration mechanisms in plants. Biologia 2017, 72, 1387–1393. [Google Scholar] [CrossRef]
- Zhang, Y.; Yao, Q.; Shi, Y.; Li, X.; Hou, L.; Xing, G.; Ahammed, G.J. Elevated co2 improves antioxidant capacity, ion homeostasis, and polyamine metabolism in tomato seedlings under ca (no3) 2-induced salt stress. Sci. Hortic. 2020, 273, 109644. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.; Lu, X.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [Green Version]
- Erland, L.A.; Murch, S.J.; Reiter, R.J.; Saxena, P.K. A new balancing act: The many roles of melatonin and serotonin in plant growth and development. Plant. Signal. Behav. 2015, 10, e1096469. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant. Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Front. Plant. Sci. 2017, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Wang, S.; Deng, X.; Yin, L.; Xiong, B.; Wang, X. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 48. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Wang, W.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Gong, X.; Shi, S.; Dou, F.; Song, Y.; Ma, F. Exogenous melatonin alleviates alkaline stress in malus hupehensis rehd. By regulating the biosynthesis of polyamines. Molecules 2017, 22, 1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.E.; Mao, J.J.; Sun, L.Q.; Huang, B.; Ding, C.B.; Gu, Y.; Liao, J.Q.; Hu, C.; Zhang, Z.W.; Yuan, S. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol. Plant. 2018, 164, 349–363. [Google Scholar] [CrossRef]
- Gao, W.; Zhang, Y.; Feng, Z.; Bai, Q.; He, J.; Wang, Y. Effects of melatonin on antioxidant capacity in naked oat seedlings under drought stress. Molecules 2018, 23, 1580. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Gao, F.; Ni, Z.; Lin, L.; Deng, Q.; Tang, Y.; Wang, X.; Luo, X.; Xia, H. Melatonin improves heat tolerance in kiwifruit seedlings through promoting antioxidant enzymatic activity and glutathione s-transferase transcription. Molecules 2018, 23, 584. [Google Scholar] [CrossRef] [Green Version]
- Debnath, B.; Hussain, M.; Li, M.; Lu, X.; Sun, Y.; Qiu, D. Exogenous melatonin improves fruit quality features, health promoting antioxidant compounds and yield traits in tomato fruits under acid rain stress. Molecules 2018, 23, 1868. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Yu, Q.; Zhang, Q.; Wang, Z.; Pan, Y.; Larssen, T.; Tang, J.; Mulder, J. Acid deposition in asia: Emissions, deposition, and ecosystem effects. Atmos. Environ. 2016, 146, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under nacl stress. J. Agr. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef]
- Borghesi, E.; González-Miret, M.L.; Escudero-Gilete, M.L.; Malorgio, F.; Heredia, F.J.; Meléndez-Martínez, A.J. Effects of salinity stress on carotenoids, anthocyanins, and color of diverse tomato genotypes. J. Agr. Food Chem. 2011, 59, 11676–11682. [Google Scholar] [CrossRef]
- Saleem, A.; Ashraf, M.; Akram, N.; Al-Qurainy, F. Salinity-induced changes in key anti-oxidant enzyme activities and in the levels of some anti-oxidants, osmo-protectants, inorganic ions, and chlorophyll pigments in okra fruit (Abelmoschus esculentus L.). J. Hortic. Sci. Biotech. 2012, 87, 271–277. [Google Scholar] [CrossRef]
- Zushi, K.; Ono, M.; Matsuzoe, N. Light intensity modulates antioxidant systems in salt-stressed tomato (Solanum lycopersicum L. Cv. Micro-tom) fruits. Sci. Hortic. 2014, 165, 384–391. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2014, 66, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Liu, T.; Chen, J.A.; Wang, W.; Simon, M.; Wu, F.; Hu, W.; Chen, J.B.; Zheng, H. A combined proteomic and transcriptomic analysis on sulfur metabolism pathways of Arabidopsis thaliana under simulated acid rain. PLoS ONE 2014, 9, e90120. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Li, S.; Zhang, L.; Qi, Y.; Ren, S. Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Tan, D.X.; Reiter, R.J.; Ye, T.; Yang, F.; Chan, Z. Melatonin induces class a1 heat-shock factors (hsfa 1s) and their possible involvement of thermotolerance in arabidopsis. J. Pineal Res. 2015, 58, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zheng, G.; Li, W.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.G.; Tan, D.X. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.-X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass (Cynodon dactylon L. Pers.) by exogenous melatonin. J. Exp. Bot. 2014, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Back, K. 2-hydroxymelatonin promotes the resistance of rice plant to multiple simultaneous abiotic stresses (combined cold and drought). J. Pineal Res. 2016, 61, 303–316. [Google Scholar] [CrossRef]





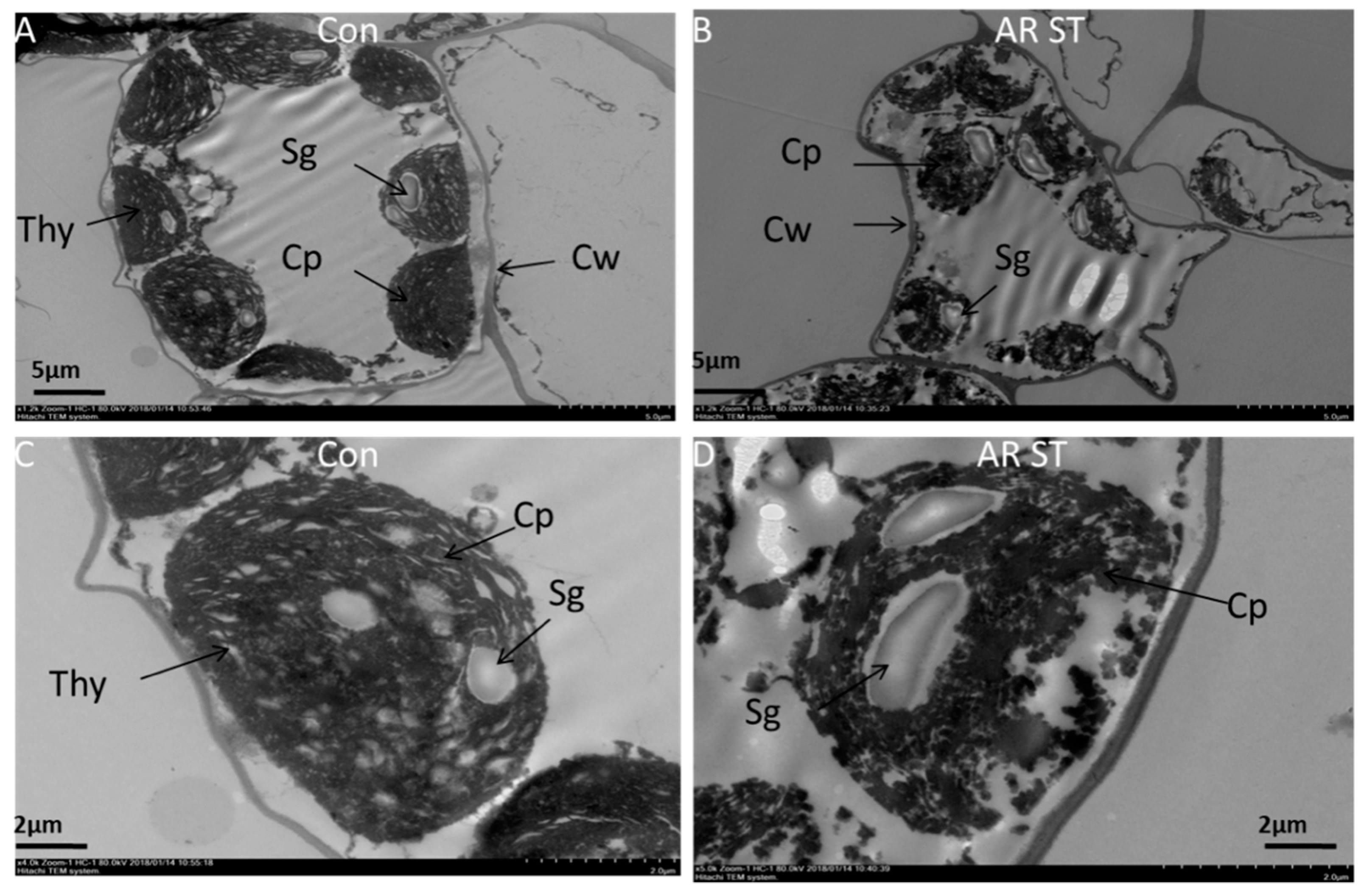{kind=link}
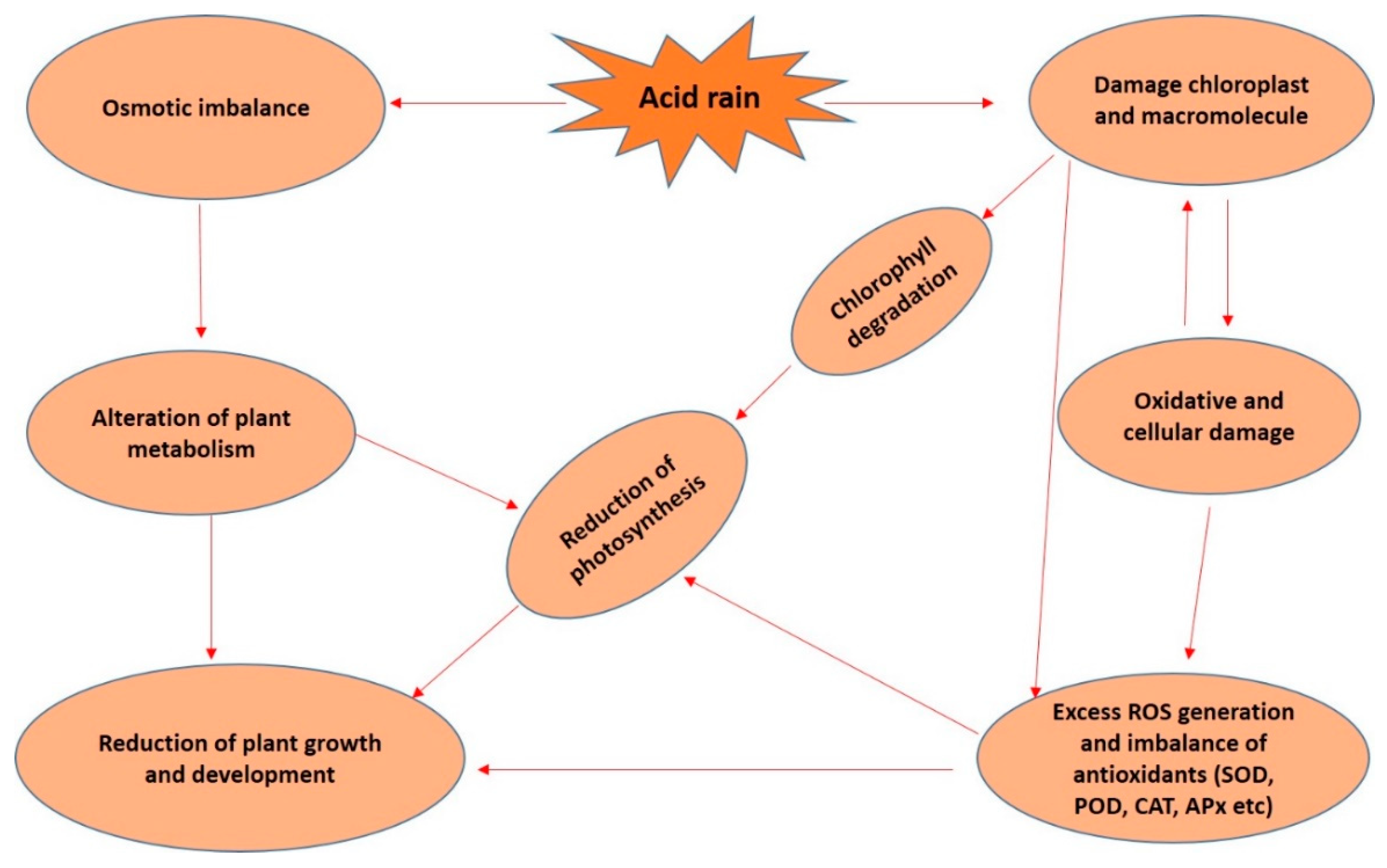{kind=link}
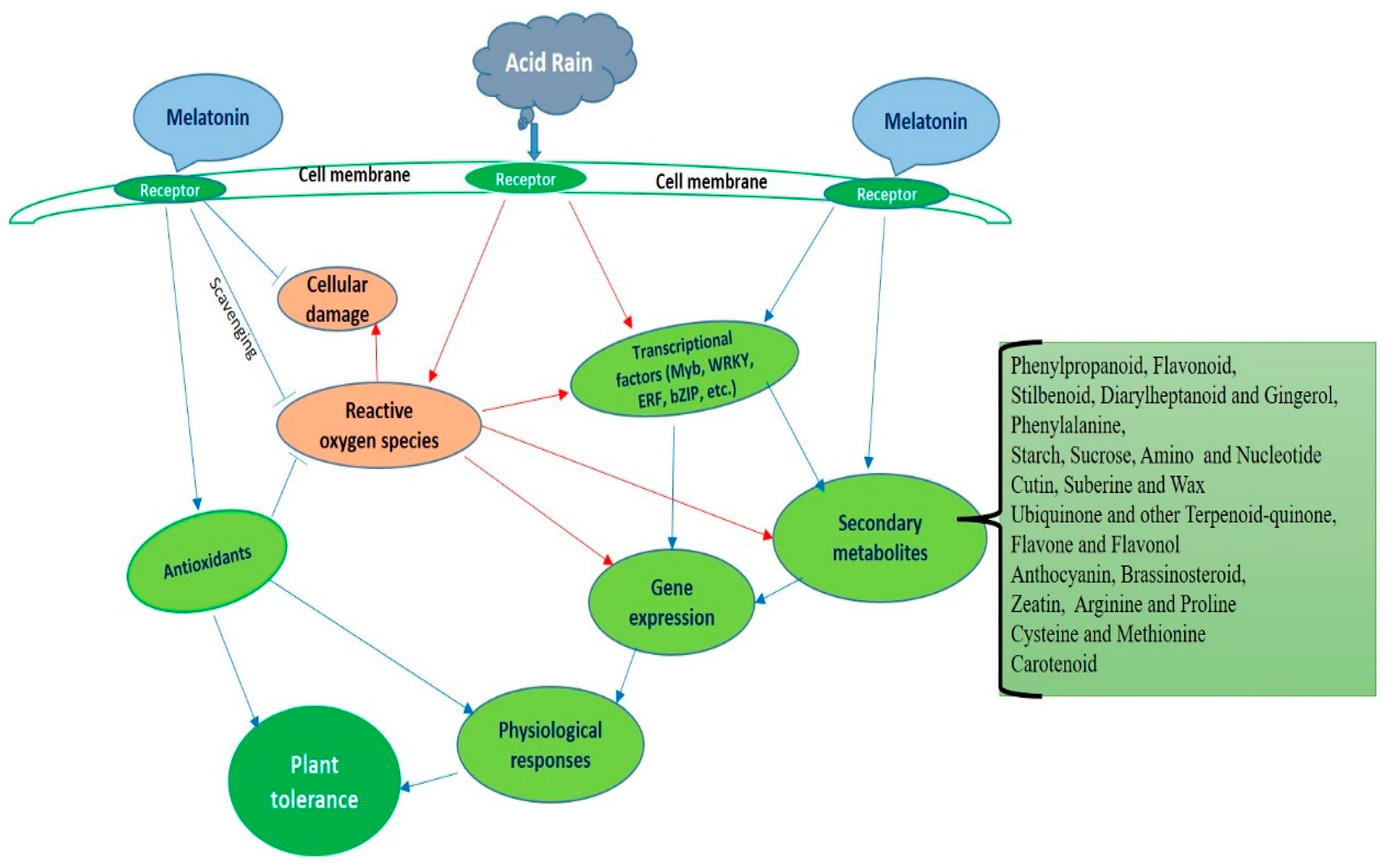{kind=link}
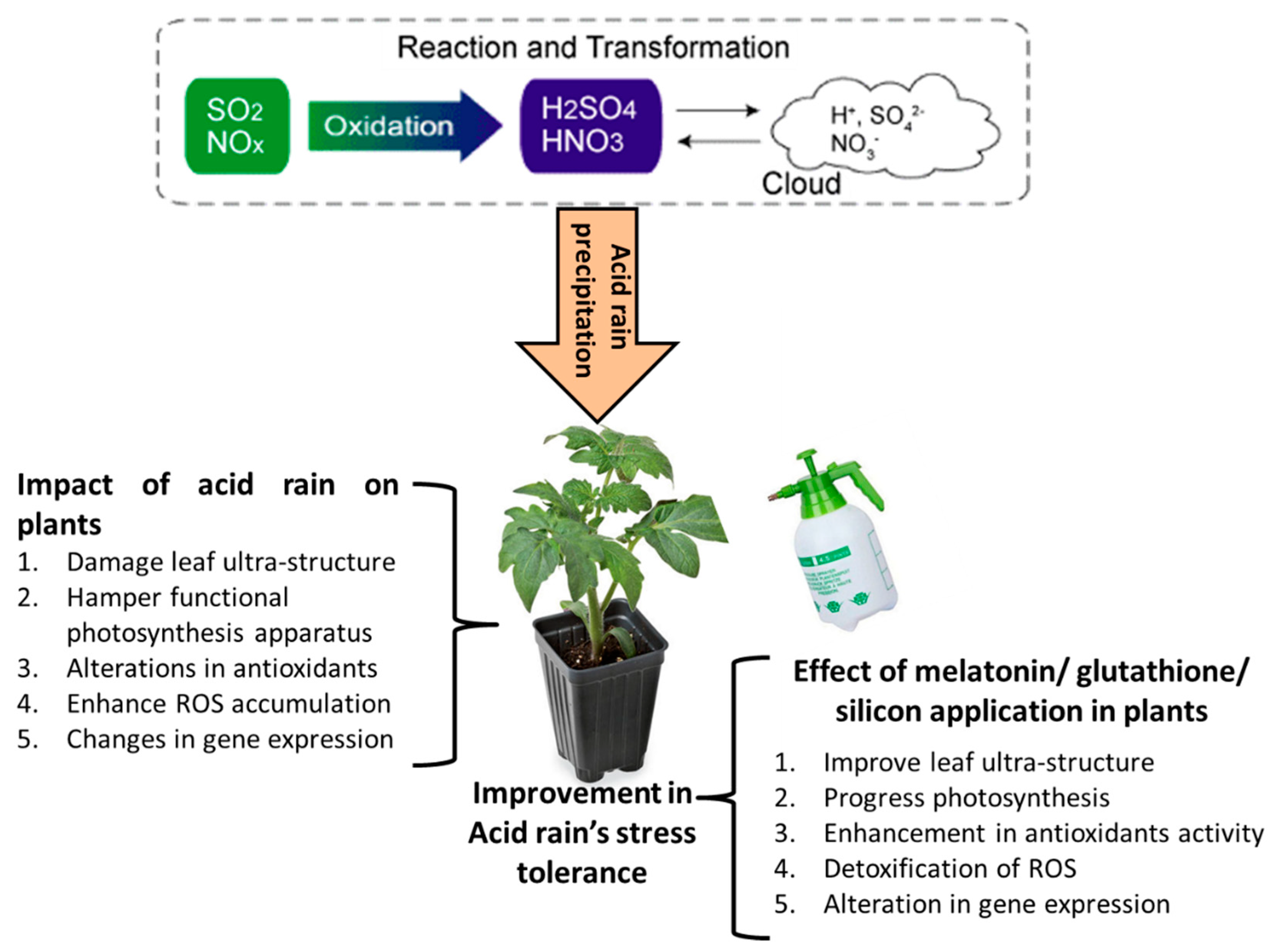{kind=link}
| Gene Description | Function | Expression | |
|---|---|---|---|
| Control vs. AR-Stressed Plants | AR-Stressed Plants vs. AR-Stressed Plants Treated with Melatonin | ||
| Caffeoyl-CoA O-methyltransferase-like | Biosynthesis of secondary metabolites, Phenylpropanoid biosynthesis, Flavonoid biosynthesis, Stilbenoid, diarylheptanoid and gingerol biosynthesis, Phenylalanine metabolism, Metabolic pathways | Downregulated | Upregulated |
| Probable galacturonosyl transferase-like 10-like | Biosynthesis of secondary metabolites, Starch and sucrose metabolism, Amino sugar and nucleotide sugar metabolism | Downregulated | Upregulated |
| Cytochrome P450 94A1-like | Biosynthesis of secondary metabolites, Cutin, suberine and wax biosynthesis, Stilbenoid, diarylheptanoid and gingerol biosynthesis, Metabolic pathways, Limonene and pinene degradation | Downregulated | Upregulated |
| Salutaridinol 7-O-acetyltransferase-like | Biosynthesis of secondary metabolites, Phenylpropanoid biosynthesis, Flavonoid biosynthesis, Stilbenoid, diarylheptanoid and gingerol biosynthesis | Downregulated | Upregulated |
| 4-coumarate-CoA ligase 2-like | Biosynthesis of secondary metabolites, Phenylpropanoid biosynthesis, Phenylalanine metabolism, Metabolic pathways, Ubiquinone and other terpenoid-quinone biosynthesis | Downregulated | Upregulated |
| Anthocyanidin 3-O-glucosyltransferase-like | Biosynthesis of secondary metabolites, Metabolic pathways, Flavone and flavonol biosynthesis, Anthocyanin biosynthesis | Downregulated | Downregulated |
| Secologanin synthase-like isoform 1 | Biosynthesis of secondary metabolites, Metabolic pathways, Brassinosteroid biosynthesis, Zeatin biosynthesis | Downregulated | Upregulated |
| Caffeoyl-CoA O-methyl transferase-like isoform 2 | Biosynthesis of secondary metabolites, Phenylpropanoid biosynthesis, Flavonoid biosynthesis, Stilbenoid, diarylheptanoid and gingerol biosynthesis, Phenylalanine metabolism, Metabolic pathways | Downregulated | Downregulated |
| S-adenosylmethionine decarboxylase proenzyme-like | Arginine and proline metabolism, Cysteine and methionine metabolism, Metabolic pathways | Downregulated | Upregulated |
| Cyanidin-3-O-glucoside 2-O-glucuronosyltransferase-like | Flavone and flavonol biosynthesis, Zeatin biosynthesis | Downregulated | Upregulated |
| Abscisic acid 8′-hydroxylase 1-like | Carotenoid biosynthesis | Downregulated | Upregulated |
| MYB-related protein Myb4-like | Stress-responsive MYB family transcriptional factor | Downregulated | Upregulated |
| Probable WRKY transcription factor 33-like | Stress-responsive WRKY family transcriptional factor Plant–pathogen interaction | Downregulated | Upregulated |
| Ethylene-responsive transcription factor 1-like | Stress-responsive ERF family transcriptional factors Plant hormone signal transduction | Downregulated | Upregulated |
| Uncharacterized protein LOC101262884 isoform 1 | Stress-responsive bZIP family transcriptional factor | Downregulated | Upregulated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debnath, B.; Sikdar, A.; Islam, S.; Hasan, K.; Li, M.; Qiu, D. Physiological and Molecular Responses to Acid Rain Stress in Plants and the Impact of Melatonin, Glutathione and Silicon in the Amendment of Plant Acid Rain Stress. Molecules 2021, 26, 862. https://doi.org/10.3390/molecules26040862
Debnath B, Sikdar A, Islam S, Hasan K, Li M, Qiu D. Physiological and Molecular Responses to Acid Rain Stress in Plants and the Impact of Melatonin, Glutathione and Silicon in the Amendment of Plant Acid Rain Stress. Molecules. 2021; 26(4):862. https://doi.org/10.3390/molecules26040862
Chicago/Turabian StyleDebnath, Biswojit, Ashim Sikdar, Shahidul Islam, Kamrul Hasan, Min Li, and Dongliang Qiu. 2021. "Physiological and Molecular Responses to Acid Rain Stress in Plants and the Impact of Melatonin, Glutathione and Silicon in the Amendment of Plant Acid Rain Stress" Molecules 26, no. 4: 862. https://doi.org/10.3390/molecules26040862