
Plumbagin, a Potent Naphthoquinone from Nepenthes Plants with Growth Inhibiting and Larvicidal Activities


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
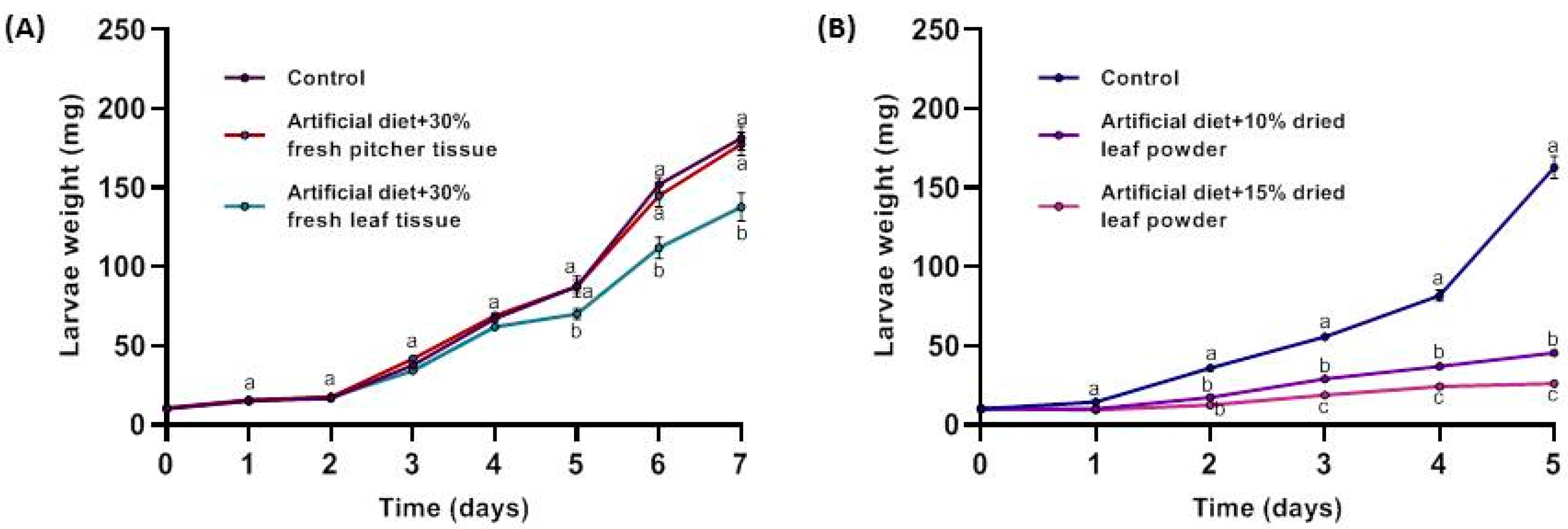
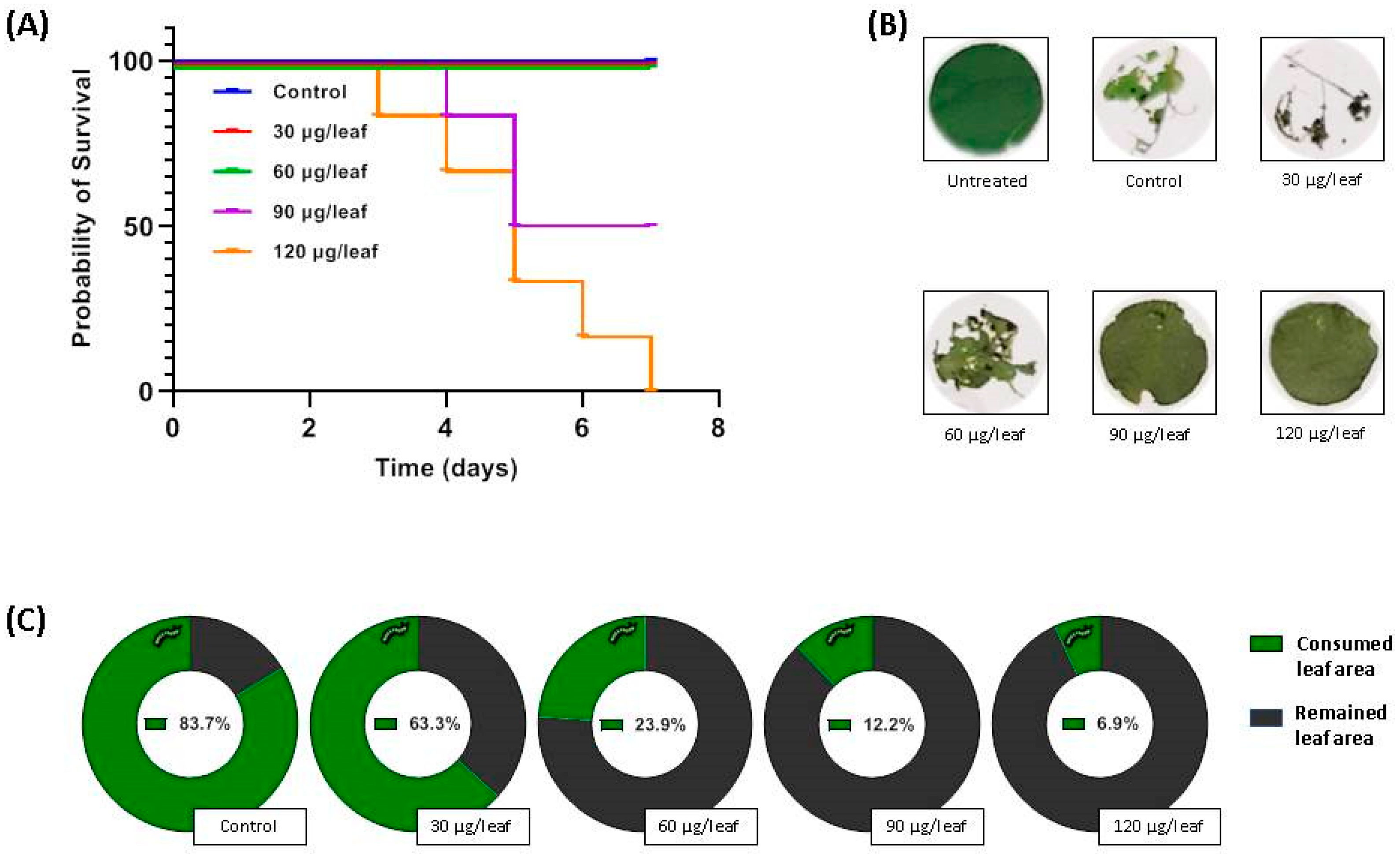
2.1. Effect of Nepenthes x ventrata Tissue on Insect Larvae Growth
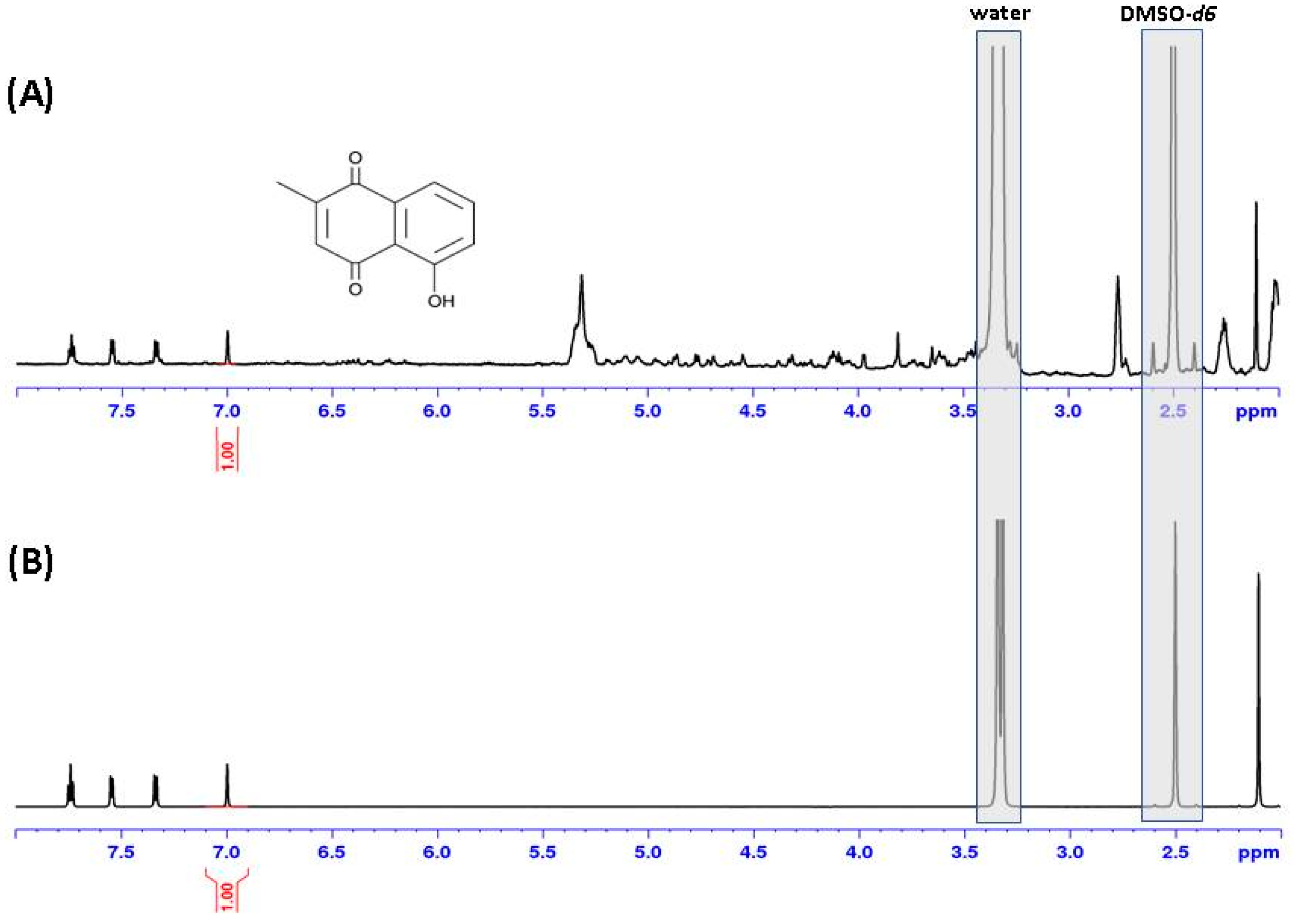
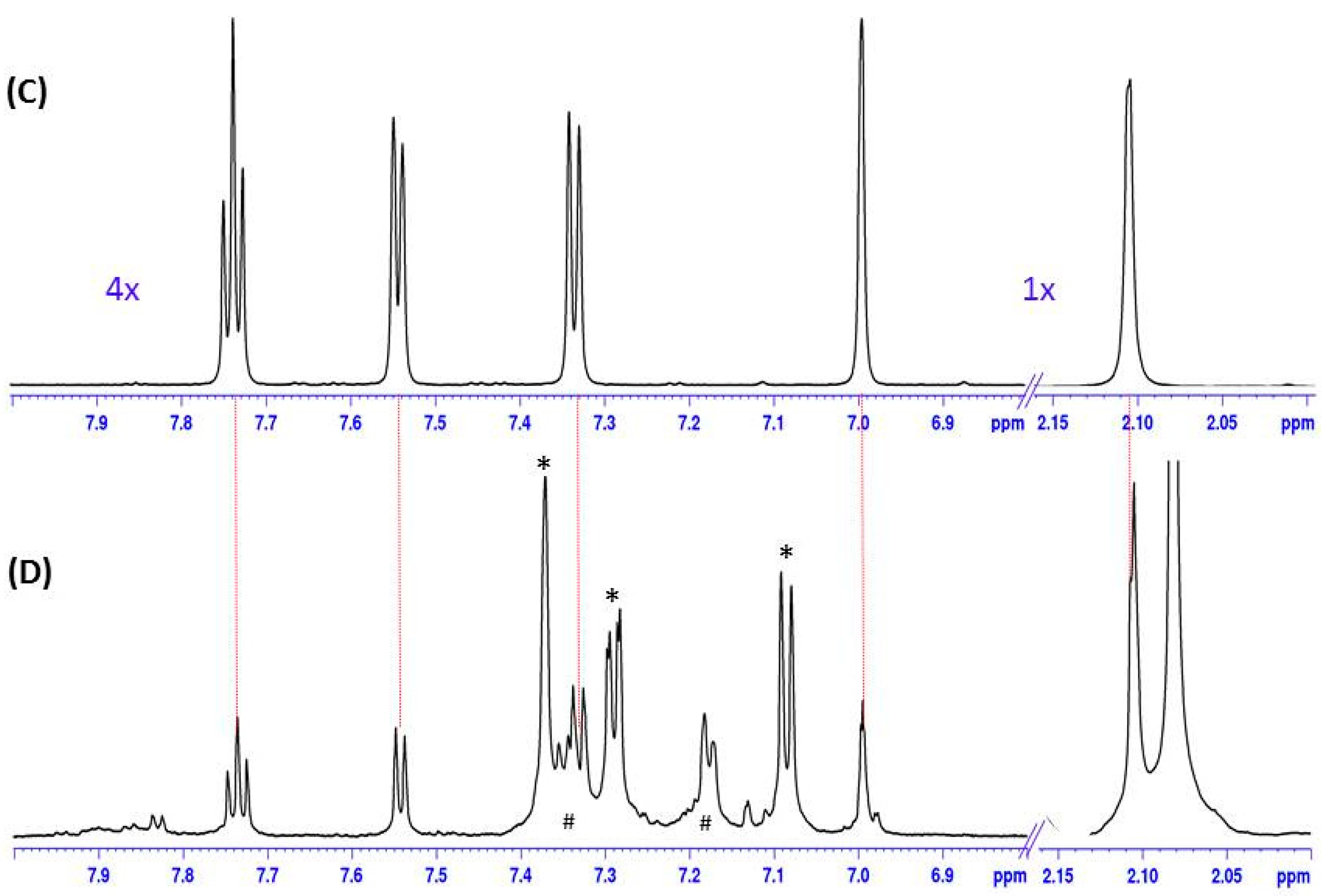
2.2. Plumbagin in Nepenthes x ventrata Tissue
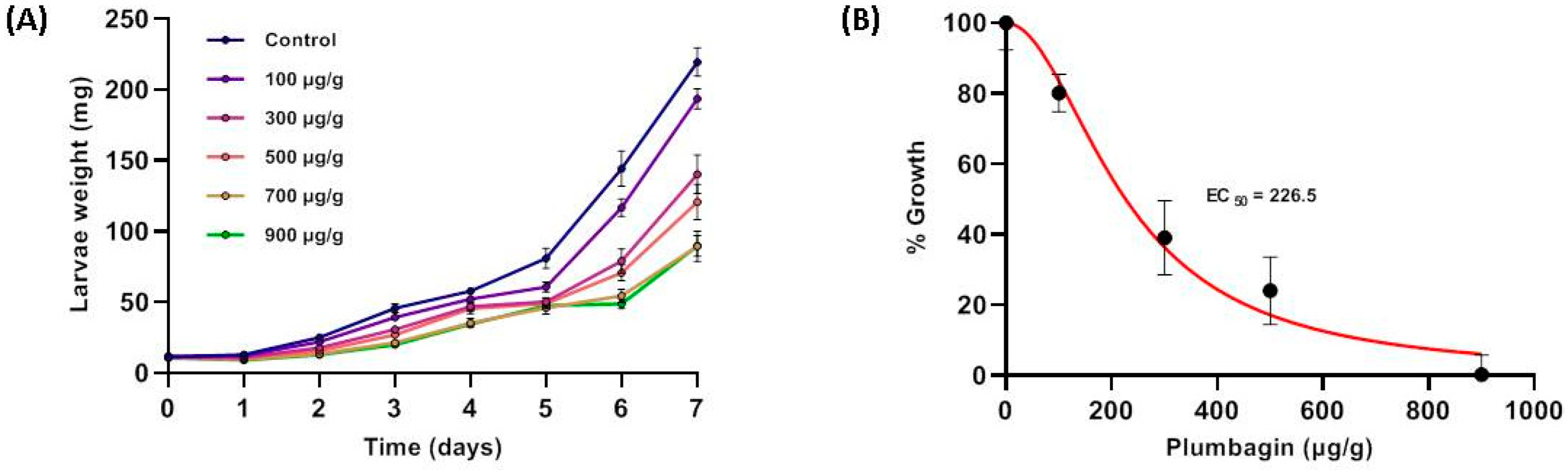
2.3. Growth-Inhibiting and Larvicidal Activities of Plumbagin
3. Materials and Methods
3.1. Insects and Plants
3.2. Feeding Assays
3.3. Isolation of Plumbagin from Nepenthes x ventrata Leaves
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juniper, B.E.; Robins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989. [Google Scholar]
- Thorogood, C.J.; Bauer, U.; Hiscock, S.J. Convergent and divergent evolution in carnivorous pitcher plant traps. New Phytol. 2018, 217, 1035–1041. [Google Scholar] [CrossRef]
- Miguel, S.; Hehn, A.; Bourgaud, F. Nepenthes: State of the art of an inspiring plant for biotechnologists. J. Biotechnol. 2018, 265, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Legendre, G.; Darnowski, D.W. Biotechnology with carnivorous plants. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 270–282. [Google Scholar]
- Chi, V.V. Dictionary of Vietnamese Medicinal Plants; Publishing House Medicine: Hanoi, Vietnam, 2012; Volume 2. [Google Scholar]
- Likhitwitayawuid, K.; Kaewamatawong, R.; Ruangrungsi, N.; Krungkrai, J. Antimalarial naphthoquinones from Nepenthes thorelii. Planta Med. 1998, 64, 237–241. [Google Scholar] [CrossRef] [PubMed]
- D’Amoto, P. The Savage Garden; Ten Speed Press: Berkeley, CA, USA, 1998. [Google Scholar]
- Wiard, C.; Morgana, S.; Khalifah, S.; Mahan, M.; Ismael, S.; Buckle, M.; Narayana, A.K.; Sulaiman, M. Antimicrobial screening of plants used for traditional medicine in the state of Perak, Peninsula Malaysia. Fitoterapia 2004, 75, 68–73. [Google Scholar]
- Rey, M.; Yang, M.; Lee, L.; Zhang, Y.; Sheff, J.G.; Sensen, C.W.; Mrazek, H.; Halada, P.; Man, P.; McCarville, J.L.; et al. Addressing proteolytic efficiency in enzymatic degradation therapy for celiac disease. Sci. Rep. 2016, 6, 30980. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-Y.; Peng, S.-Y.; Cheng, Y.-B.; Wang, C.-L.; Farooqi, A.A.; Yu, T.-J.; Hou, M.-F.; Wang, S.-C.; Yem, C.-H.; Chan, L.-P.; et al. Ethyl acetat extract of Nepenthes adrianii x clipeata induces antiproliferation, apoptosis, and DNA damage against oral cancer cells through oxidative stress. Environ. Toxcol. 2019, 34, 891–901. [Google Scholar] [CrossRef]
- Eilenberg, H.; Pnini-Cohen, S.; Rahamim, Y.; Sionov, E.; Segal, E.; Carmeli, S.; Zilberstein, A. Induced production of antifungal naphthoquinones in the pitchers of the carnivorous plant Nepenthes khasiana. J. Exp. Bot. 2010, 61, 911–922. [Google Scholar] [CrossRef] [Green Version]
- Buch, F.; Rott, M.; Rottloff, S.; Paetz, C.; Hilke, I.; Raessler, M.; Mithöfer, A. 2013. Secreted pitfall-trap fluid of carnivorous Nepenthes plants is unsuitable for microbial growth. Ann. Bot. 2013, 111, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Mithöfer, A. Carnivorous pitcher plants: Insights in an old topic. Phytochemistry 2011, 72, 1678–1682. [Google Scholar] [CrossRef]
- Hatcher, C.R.; Ryves, D.B.; Millett, J. The function of secondary metabolites in plant carnivory. Ann. Bot. 2020, 125, 399–411. [Google Scholar] [CrossRef]
- Raj, G.; Kurup, R.; Hussain, A.A.; Baby, S. Distribution of naphthoquinones, plumbagin, droserone, and 5–O–methyl droserone in chitin-induced and uninduced Nepenthes khasiana: Molecular events in prey capture. J. Exp. Bot. 2011, 62, 5429–5436. [Google Scholar] [CrossRef] [Green Version]
- Rischer, H.; Hamm, A.; Bringmann, G. Nepenthes insignis uses a C2-portion of the carbon skeleton of L-alanine acquired via its carnivorous organs, to build up the allelochemical plumbagin. Phytochemistry 2002, 59, 603–609. [Google Scholar] [CrossRef]
- Schlauer, J.; Nerz, J.; Rischer, H. Carnivorous plant chemistry. Acta Bot. Gall. 2005, 152, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cai, Y.; He, C.; Chen, M.; Li, H. Anticancer properties and pharmaceutical applications of plumbagin: A review. Am. J. Chin. Med. 2017, 45, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.K.; Panda, M.; Biswal, B.K. Emerging role of plumbagin: Cytotoxic potential and pharmaceutical relevance towards cancer therapy. Food Chem. Toxicol. 2019, 215, 566–583. [Google Scholar] [CrossRef]
- Yin, Z.H.; Zhang, J.J.; Chen, L.; Gio, Q.F.; Yang, B.C.; Zhang, W.; Kang, W.Y. Anticancer effects and mechanisms of action of plumbagin: Review of research advances. Biomed Res. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Panichayupakaranant, P.; Ahmad, M.I. Plumbagin and its role in chronic diseases. Adv. Exp. Med. Biol. 2016, 929, 229–246. [Google Scholar]
- Aung, H.; Chia, L.; Goh, N.K.; Chia, T.F.; Ahmad, A.A.; Pare, P.W.; Mabry, T.J. Phenolic constituents from the leaves of the carnivorous Nepenthes gracilis. Fitoterapia 2002, 73, 445–447. [Google Scholar] [CrossRef]
- Wan, A.S.; Aexel, R.T.; Ramsey, R.B.; Nicholas, H.J. Sterols and triterpenes of the pitcher plant. Phytochemistry 1972, 11, 456–461. [Google Scholar] [CrossRef]
- Dávila-Lara, A.; Rodríguez-López, C.E.; O’Connor, S.E.; Mithöfer, A. Metabolomics analysis reveals tissue-specific metabolite compositions in leaf blade and traps of carnivorous Nepenthes plants. Int. J. Mol. Sci. 2020, 21, 4376. [Google Scholar] [CrossRef]
- Carmickle, R.N.; Horner, J.D. Impact of the specialist herbivore Exyra semicrocea on the carnivorous plant Sarracenia alata: A field experiment testing the effects of tissue loss and diminished prey capture on plant growth. Plant Ecol. 2019, 220, 553–561. [Google Scholar] [CrossRef]
- Lamb, T.; Kalies, E.L. An overview of lepidopteran herbivory on North American pitcher plants (Sarracenia), with a novel observation of feeding on Sarracenia flava. J. Lepid. Soc. 2020, 74, 193–197. [Google Scholar]
- Merbach, M.A.; Zizka, G.; Fiala, B.; Merbach, D.; Booth, W.E.; Maschwitz, U. Why a carnivorous plant cooperates with an ant–selective defense against pitcher-nutritional mutualism in a pitcher plant destroying weevils in the myrmecophytic pitcher plant Nepenthes bicalcarata Hook. F. Ecotropica 2007, 13, 45–56. [Google Scholar]
- Gilbert, K.J.; Nitta, J.H.; Talavera, G.; Pierce, N.E. Keeping an eye on coloration: Ecological correlates of the evolution of pitcher traits in the genus Nepenthes (Caryophyllales). Biol. J. Linn. Soc. 2018, 123, 321–337. [Google Scholar] [CrossRef] [Green Version]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of carnivory in angiosperms. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 22–42. [Google Scholar]
- Devi, S.P.; Kumaria, S.; Rao, S.R.; Tandon, P. Carnivorous plants as a source of potent bioactive compounds: Naphthoquinones. Tropical Plant Biol. 2016, 9, 267–279. [Google Scholar] [CrossRef]
- Bubla, P.; Adam, V.; Havel, L.; Kizek, R. Noteworthy secondary metabolites naphthoquinones—their occurrence, pharmacological properties and analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Widhalm, J.R.; Rhodes, D. Biosynthesis and molecular actions of specialized 1,4-naphthoquinone natural products produced by horticultural plants. Hortic. Res. 2016, 3, 16046. [Google Scholar] [CrossRef]
- Tripathi, K.S.; Rengasamy, K.R.R.; Biswal, B.K. Plumbagin engenders apoptosis in lung cancer celles via caspase-9 activation and targeting mitochondrial-mediated ROD induction. Arch. Pharm. Res. 2020, 43, 242–256. [Google Scholar] [CrossRef]
- Soderquist, C.J. Juglone and allelopathy. J. Chem. Educ. 1973, 50, 782–783. [Google Scholar] [CrossRef]
- Willis, R.J. Juglans spp., juglone and allelopathy. Allelopath. J. 2000, 7, 1–55. [Google Scholar]
- Buch, F.; Pauchet, Y.; Rott, M.; Mithöfer, A. Characterization and heterologous expression of a novel PR-1 protein from traps of the carnivorous plant Nepenthes mirabilis. Phytochemistry 2014, 100, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Sreelatha, T.; Hymavathi, A.; Babu, K.S.; Murthy, J.M.; Pathipati, U.R.; Rao, J.M. Synthesis and insect antifeedant activity of plumbagin derivatives with the amino acid moiety. Agric. Food Chem. 2009, 57, 6090–6094. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Takada, N.; Ueda, M. Mechanism of antifeedant activity of plumbagin, a compound concerning the chemical defense in carnivorous plant. Tetrahedron Lett. 2004, 45, 7115–7119. [Google Scholar] [CrossRef]
- Tokunaga, T.; Dohmura, A.; Takada, N.; Ueda, M. Cytotoxic antifeedant from Dionaea muscipula Ellis: A defensive mechanism of carnivorous plants against predators. Bull. Chem. Soc. Jpn. 2004, 77, 537–541. [Google Scholar] [CrossRef]
- Akhtar, Y.; Isman, M.B.; Niehaus, L.A.; Lee, C.-H.; Lee, H.-S. Antifeedant and toxic effects of naturally occurring and synthetic quinones to the cabbage looper, Trichoplusia ni. Crop Prot. 2012, 31, 8–14. [Google Scholar] [CrossRef]
- Hu, W.; Du, W.; Bai, S.; Lv, S.; Chen, G. Phenoloxidase, an effective bioactivity target for botanical insecticide screening from green walnut husks. Nat. Prod. Res. 2018, 32, 2848–2851. [Google Scholar] [CrossRef]
- Pavela, R. Efficacy of naphthoquinones as insecticides against the house fly, Musca domestica L. Ind. Crops Prod. 2013, 43, 745–750. [Google Scholar] [CrossRef]
- Meents, A.K.; Chen, S.-P.; Reichelt, M.; Lu, H.-H.; Bartram, S.; Yeh, K.-W.; Mithöfer, A. Volatile DMNT systemically induces jasmonate-independent direct anti-herbivore defense in leaves of sweet potato (Ipomoea batatas) plants. Sci. Rep. 2019, 9, 17431. [Google Scholar] [CrossRef] [Green Version]
- Kubo, I.; Uchida, M.; Klocke, J.A. An insect ecdysis inhibitor from the African medical plant Plumbago capensis (Plumbaginaceae); a naturally occurring chitin synthetase inhibitor. Agric. Biol. Chem. 1983, 47, 911–913. [Google Scholar]
- Mitchell, M.J.; Smith, S.L. Effects of the chitin synthetase inhibitor plumbagin and its 2-dimethyl derivative juglone on insect ecdysone 20-monooxigenase activity. Experientia 1988, 44, 990–991. [Google Scholar] [CrossRef]
- Gujar, G.T. Interaction of plumbagin with hormones in the cotton stainer, Dysdercus koenigii Fabricius. Proc. Natl. Acad. Sci. India B 1993, 9, 477–482. [Google Scholar]
- Pradeepa, V.; Senthil-Nathan, S.; Sathish-Narayanan, S.; Selin-Rani, S.; Vasantha-Srinivasan, P.; Thanigaivel, A.; Ponsankar, A.; Edwin, E.S.; Sakthi-Bagavathy, M.; Kalaivani, K.; et al. Potential mode of action of a novel plumbagin as a mosquito repellent against the malarial vector Anopheles stephensi, (Culicidae: Diptera). Pestic. Biochem. Physiol. 2016, 134, 84–93. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman-Soad, A.; Dávila-Lara, A.; Paetz, C.; Mithöfer, A. Plumbagin, a Potent Naphthoquinone from Nepenthes Plants with Growth Inhibiting and Larvicidal Activities. Molecules 2021, 26, 825. https://doi.org/10.3390/molecules26040825
Rahman-Soad A, Dávila-Lara A, Paetz C, Mithöfer A. Plumbagin, a Potent Naphthoquinone from Nepenthes Plants with Growth Inhibiting and Larvicidal Activities. Molecules. 2021; 26(4):825. https://doi.org/10.3390/molecules26040825
Chicago/Turabian StyleRahman-Soad, Asifur, Alberto Dávila-Lara, Christian Paetz, and Axel Mithöfer. 2021. "Plumbagin, a Potent Naphthoquinone from Nepenthes Plants with Growth Inhibiting and Larvicidal Activities" Molecules 26, no. 4: 825. https://doi.org/10.3390/molecules26040825