
Potential Role of Melatonin as an Adjuvant for Atherosclerotic Carotid Arterial Stenosis

{kind=link}
{kind=link}
Abstract
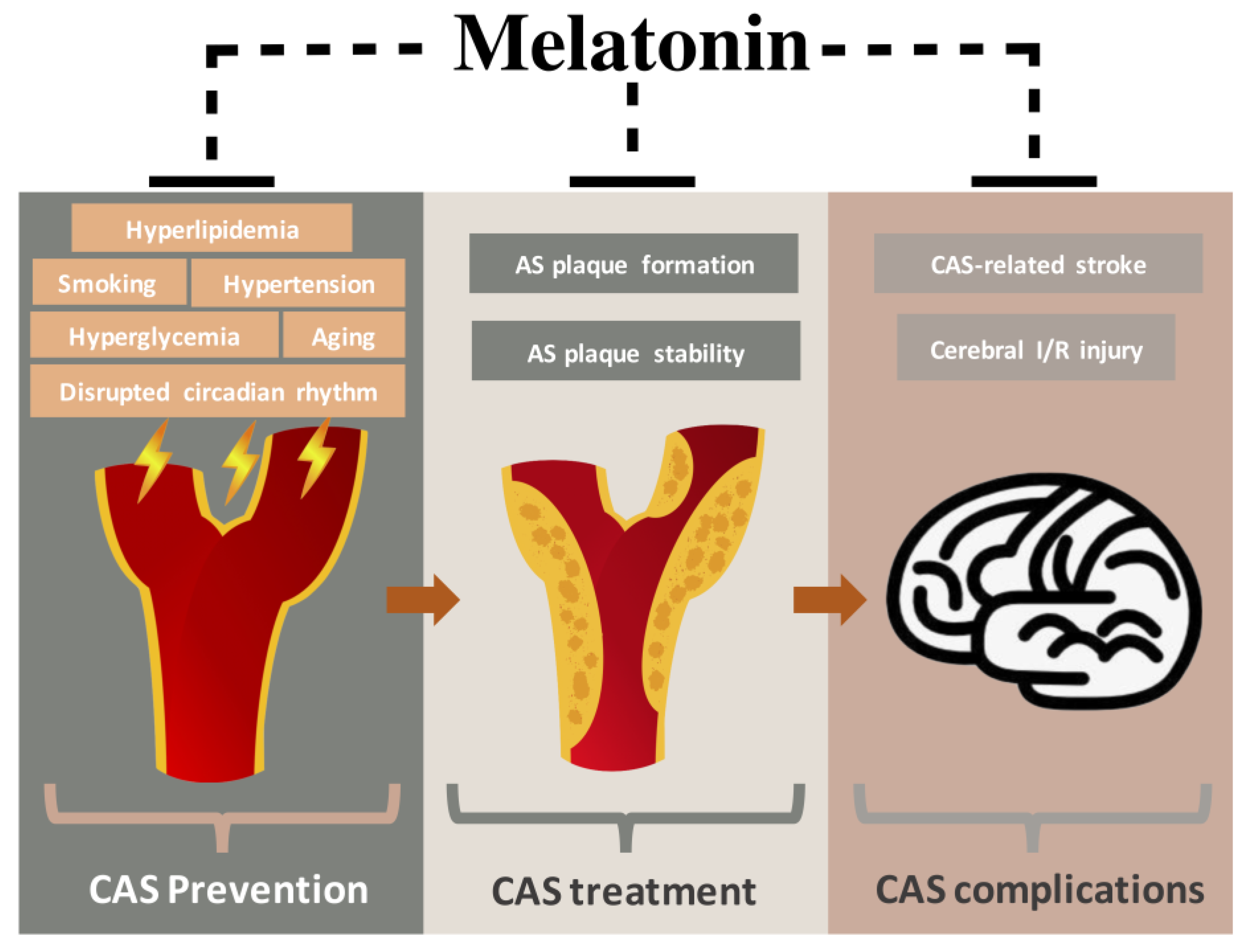
:1. Introduction
2. Pathophysiology of CAS
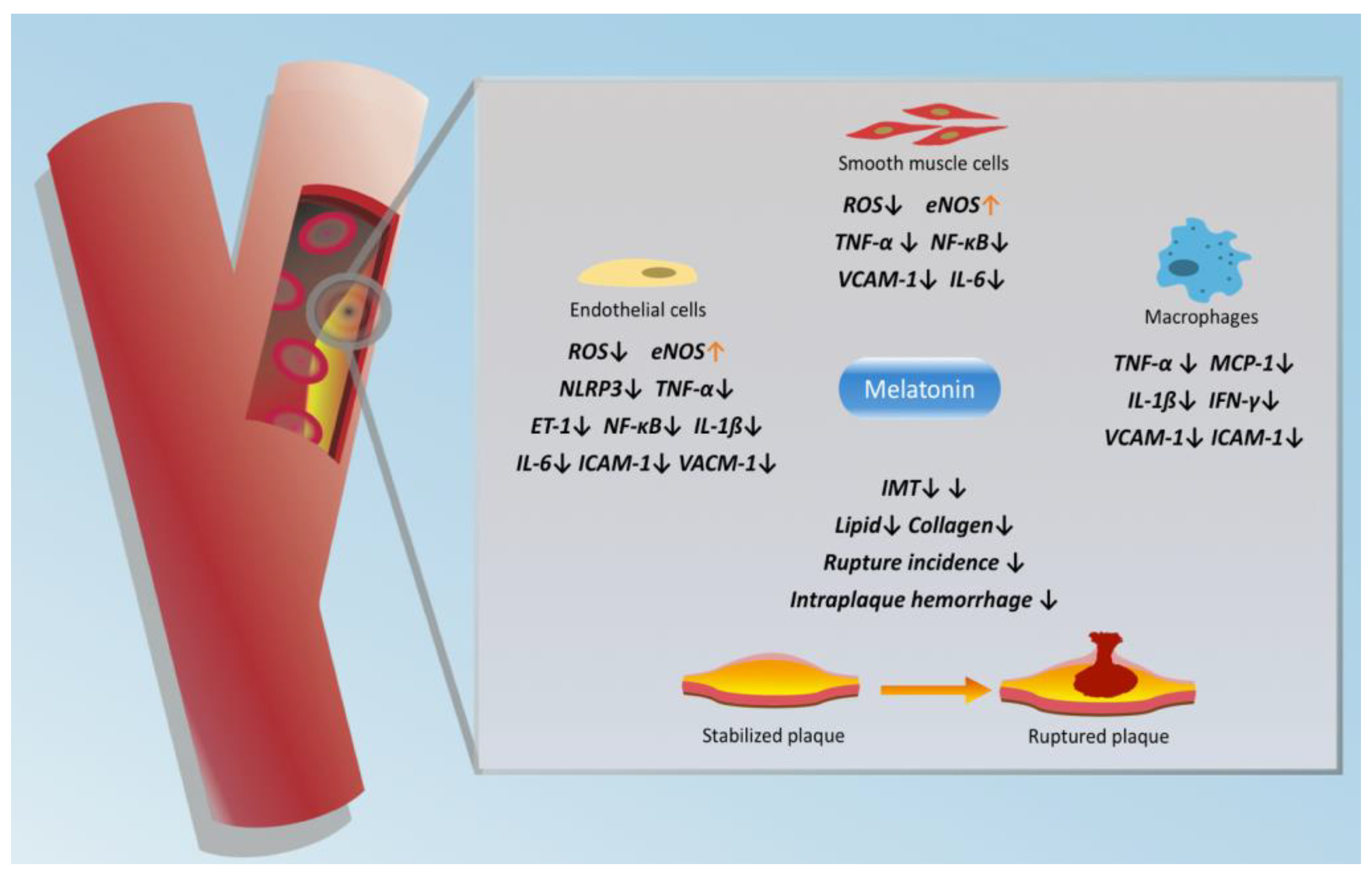
3. Protective Effects of Melatonin in CAS
4. Melatonin & CAS-Related Ischemic Stroke
5. Melatonin & Cerebral Ischemia/Reperfusion Injury
6. Preventive Effects of Melatonin in CAS-Related Risk Factors
6.1. Smoking
6.2. Hypertension
6.3. Hyperlipidemia
6.4. Hyperglycemia
6.5. Aging
6.6. Disrupted Circadian Rhythm
7. Clinical Implications and Limitations
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Weerd, M.; Greving, J.P.; De Jong, A.W.; Buskens, E.; Bots, M.L. Prevalence of Asymptomatic Carotid Artery Stenosis According to Age and Sex. Stroke 2009, 40, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Dharmakidari, S.; Bhattacharya, P.; Chaturvedi, S. Carotid Artery Stenosis: Medical Therapy, Surgery, and Stenting. Curr. Neurol. Neurosci. Rep. 2017, 17, 77. [Google Scholar] [CrossRef]
- Malekmohammad, K.; Sewell, R.D.E.; Rafieian-Kopaei, M. Antioxidants and Atherosclerosis: Mechanistic Aspects. Biomolecules 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting Early Atherosclerosis: A Focus on Oxidative Stress and Inflammation. Oxidative Med. Cell. Longev. 2019, 2019, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Rosales-Corral, S.; Galano, A.; Zhou, X.; Xu, B. Mitochondria: Central Organelles for Melatonin′s Antioxidant and Anti-Aging Actions. Molecules 2018, 23, 509. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Liu, X.; Guo, W. Melatonin attenuates inflammation-related venous endothelial cells apoptosis through modulating the MST1–MIEF1 pathway. J. Cell. Physiol. 2019, 234, 23675–23684. [Google Scholar] [CrossRef] [PubMed]
- Boga, J.A.; Caballero, B.; Potes, Y.; Pérez-Martínez, Z.; Reiter, R.J.; Vega-Naredo, I.; Coto-Montes, A. Therapeutic potential of melatonin related to its role as an autophagy regulator: A review. J. Pineal Res. 2019, 66, e12534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coto-Montes, A.; Boga, J.A.; Rosales-Corral, S.; Fuentes-Broto, L.; Tan, D.-X.; Reiter, R.J. Role of melatonin in the regulation of autophagy and mitophagy: A review. Mol. Cell. Endocrinol. 2012, 361, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Reiter, R.J.; Chen, Y.-D. Role of melatonin in controlling angiogenesis under physiological and pathological conditions. Angiogenesis 2019, 23, 91–104. [Google Scholar] [CrossRef]
- Wang, X.; Bian, Y.; Zhang, R.; Liu, X.; Ni, L.; Ma, B.; Zeng, R.; Zhao, Z.; Song, X.; Liu, C. Melatonin alleviates cigarette smoke-induced endothelial cell pyroptosis through inhibiting ROS/NLRP3 axis. Biochem. Biophys. Res. Commun. 2019, 519, 402–408. [Google Scholar] [CrossRef]
- Ma, B.; Chen, Y.; Wang, X.; Zhang, R.; Niu, S.; Ni, L.; Di, X.; Han, Q.; Liu, C. Cigarette smoke exposure impairs lipid metabolism by decreasing low-density lipoprotein receptor expression in hepatocytes. Lipids Heal. Dis. 2020, 19, 88. [Google Scholar] [CrossRef]
- Tung, Y.-T.; Chiang, P.-C.; Chen, Y.-L.; Chien, Y.-W. Effects of Melatonin on Lipid Metabolism and Circulating Irisin in Sprague-Dawley Rats with Diet-Induced Obesity. Molecules 2020, 25, 3329. [Google Scholar] [CrossRef]
- Sadanandan, N.; Cozene, B.; Cho, J.; Park, Y.J.; Saft, M.; Gonzales-Portillo, B.; Borlongan, C.V. Melatonin—A Potent Therapeutic for Stroke and Stroke-Related Dementia. Antioxidants 2020, 9, 672. [Google Scholar] [CrossRef]
- Mao, Z.-J.; Lin, H.; Xiao, F.-Y.; Huang, Z.-Q.; Chen, Y.-H. Melatonin against Myocardial Ischemia-Reperfusion Injury: A Meta-analysis and Mechanism Insight from Animal Studies. Oxidative Med. Cell. Longev. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Simko, F.; Baka, T.; Krajcirovicova, K.; Bednarova, K.R.; Aziriova, S.; Zorad, S.; Poglitsch, M.; Adamcova, M.; Reiter, R.J.; Paulis, L. Effect of Melatonin on the Renin-Angiotensin-Aldosterone System in l-NAME-Induced Hypertension. Molecules 2018, 23, 265. [Google Scholar] [CrossRef] [Green Version]
- Nduhirabandi, F.; Maarman, G. Melatonin in Heart Failure: A Promising Therapeutic Strategy? Molecules 2018, 23, 1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekin, G.; Isbir, S.; Şener, G.; Özge, Ç.; Çetinel, Ş.; Dericioğlu, O.; Arsan, S.; Çobanoğlu, A. The preventive and curative effects of melatonin against abdominal aortic aneurysm in rats. J. Vasc. Surg. 2018, 67, 1546–1555. [Google Scholar] [CrossRef] [PubMed]
- Léger, D.; Quera-Salva, M.-A.; Vecchierini, M.-F.; Ogrizek, P.A.; Perry, C.A.; Dressman, M. Safety profile of tasimelteon, a melatonin MT1and MT2receptor agonist: Pooled safety analyses from six clinical studies. Expert Opin. Drug Saf. 2015, 14, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Sanchez-Sanchez, J.J.; Kaski, J.C.; Reiter, R.J. Melatonin and circadian biology in human cardiovascular disease. J. Pineal Res. 2010, 49, 14–22. [Google Scholar] [CrossRef]
- Pelisek, J.; Eckstein, H.-H.; Zernecke, A. Pathophysiological Mechanisms of Carotid Plaque Vulnerability: Impact on Ischemic Stroke. Arch. Immunol. Ther. Exp. 2012, 60, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Brinjikji, W.; Huston, J.; Rabinstein, A.A.; Kim, G.-M.; Lerman, A.; Lanzino, G. Contemporary carotid imaging: From degree of stenosis to plaque vulnerability. J. Neurosurg. 2016, 124, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Kassem, M.; Florea, A.; Mottaghy, F.M.; Van Oostenbrugge, R.; Kooi, M.E. Magnetic resonance imaging of carotid plaques: Current status and clinical perspectives. Ann. Transl. Med. 2020, 8, 1266. [Google Scholar] [CrossRef]
- Kramer, C.M.; Treiman, G.S. Vulnerable Plaque in Carotid Arteries Without “Significant” Stenosis. J. Am. Coll. Cardiol. 2020, 76, 2223–2225. [Google Scholar] [CrossRef]
- Polak, J.F.; O’Leary, D.H. Carotid Intima-Media Thickness as Surrogate for and Predictor of CVD. Glob. Hear. 2016, 11, 295–312.e3. [Google Scholar] [CrossRef]
- Yang, G.-H.; Li, Y.-C.; Wang, Z.-Q.; Liu, B.; Ye, W.; Ni, L.; Zeng, R.; Miao, S.-Y.; Wang, L.-F.; Liu, C.-W. Protective effect of melatonin on cigarette smoke-induced restenosis in rat carotid arteries after balloon injury. J. Pineal Res. 2014, 57, 451–458. [Google Scholar] [CrossRef]
- Hu, Z.-P.; Fang, X.-L.; Fang, N.; Wang, X.-B.; Qian, H.-Y.; Cao, Z.; Cheng, Y.; Wang, B.-N.; Wang, Y. Melatonin ameliorates vascular endothelial dysfunction, inflammation, and atherosclerosis by suppressing the TLR4/NF-κB system in high-fat-fed rabbits. J. Pineal Res. 2013, 55, 388–398. [Google Scholar] [CrossRef]
- Li, H.; Leu, Y.-L.; Wu, Y.-C.; Wang, S.-H. Melatonin Inhibits in Vitro Smooth Muscle Cell Inflammation and Proliferation and Atherosclerosis in Apolipoprotein E-Deficient Mice. J. Agric. Food Chem. 2019, 67, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Von Der Thüsen, J.H.; Van Berkel, T.J.C.; Biessen, E.A. Induction of Rapid Atherogenesis by Perivascular Carotid Collar Placement in Apolipoprotein E–Deficient and Low-Density Lipoprotein Receptor–Deficient Mice. Circ. 2001, 103, 1164–1170. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Lin, N.; Sheng, X.; Zhao, Y.; Su, Y.; Xu, L.; Tong, R.; Yan, Y.; Fu, Y.; He, J.; et al. Melatonin stabilizes rupture-prone vulnerable plaques via regulating macrophage polarization in a nuclear circadian receptor RORα-dependent manner. J. Pineal Res. 2019, 67, e12581. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Jiang, X.; Liu, S.; Liu, Y.; Chen, W.; Yang, J.; Zhang, C.; Zhang, W. Melatonin enhances atherosclerotic plaque stability by inducing prolyl-4-hydroxylase α1 expression. J. Hypertens. 2019, 37, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sharma, R.; Misra, S.; Kumar, A.; Nath, M.; Nair, P.; Vibha, D.; Srivastava, A.K.; Prasad, K. CIMT as a risk factor for stroke subtype: A systematic review. Eur. J. Clin. Investig. 2020, 50, e13348. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Kim, K.H.; Park, H.; Cho, J.Y.; Hong, Y.J.; Park, H.W.; Kim, J.H.; Ahn, Y.; Jeong, M.H.; Cho, J.G.; et al. Carotid plaque rather than intima-media thickness as a predictor of recurrent vascular events in patients with acute ischemic stroke. Cardiovasc. Ultrasound 2017, 15, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caso, J.R.; Pradillo, J.M.; Hurtado, O.; Lorenzo, P.; Moro, M.A.; Lizasoain, I. Toll-Like Receptor 4 Is Involved in Brain Damage and Inflammation After Experimental Stroke. Circulation 2007, 115, 1599–1608. [Google Scholar] [CrossRef]
- Chen, K.; Lin, K.; Ko, S.; Chiang, J.Y.; Guo, J.; Yip, H.-K. Melatonin against acute ischaemic stroke dependently via suppressing both inflammatory and oxidative stress downstream signallings. J. Cell. Mol. Med. 2020, 24, 10402–10419. [Google Scholar] [CrossRef]
- Merlo, S.; Luaces, J.P.; Spampinato, S.F.; Toro-Urrego, N.; Caruso, G.I.; D’Amico, F.; Capani, F.; Sortino, M.A. SIRT1 Mediates Melatonin’s Effects on Microglial Activation in Hypoxia: In Vitro and In Vivo Evidence. Biomolecules 2020, 10, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-Y.; Chen, T.-Y.; Lee, M.-Y.; Chen, S.-T.; Hsu, Y.-S.; Kuo, Y.-L.; Chang, G.-L.; Wu, T.-S.; Lee, E.-J. Melatonin decreases neurovascular oxidative/nitrosative damage and protects against early increases in the blood–brain barrier permeability after transient focal cerebral ischemia in mice. J. Pineal Res. 2006, 41, 175–182. [Google Scholar] [CrossRef]
- Tai, S.-H.; Chen, H.-Y.; Lee, E.-J.; Chen, T.-Y.; Lin, H.-W.; Hung, Y.-C.; Huang, S.-Y.; Chen, Y.-H.; Lee, W.-T.; Wu, T.-S. Melatonin inhibits postischemic matrix metalloproteinase-9 (MMP-9) activation via dual modulation of plasminogen/plasmin system and endogenous MMP inhibitor in mice subjected to transient focal cerebral ischemia. J. Pineal Res. 2010, 49, 332–341. [Google Scholar] [CrossRef]
- Lee, E.-J.; Wu, T.-S.; Lee, M.-Y.; Chen, T.-Y.; Tsai, Y.-Y.; Chuang, J.-I.; Chang, G.-L. Delayed treatment with melatonin enhances electrophysiological recovery following transient focal cerebral ischemia in rats. J. Pineal Res. 2004, 36, 33–42. [Google Scholar] [CrossRef]
- Lorente, L.; Martín, M.M.; Abreu-González, P.; Pérez-Cejas, A.; Ramos, L.; Argueso, M.; Solé-Violán, J.; Cáceres, J.J.; Jiménez, A.; García-Marín, V. Serum melatonin levels are associated with mortality in patients with malignant middle cerebral artery infarction. J. Int. Med. Res. 2018, 46, 3268–3277. [Google Scholar] [CrossRef]
- Kaku, Y.; Yoshimura, S.-I.; Kokuzawa, J. Factors predictive of cerebral hyperperfusion after carotid angioplasty and stent placement. Am. J. Neuroradiol. 2004, 25, 1403–1408. [Google Scholar]
- Heo, S.-H.; Yoon, K.-W.; Woo, S.-Y.; Park, Y.-J.; Kim, Y.-W.; Kim, K.-H.; Chung, C.-S.; Bang, O.-Y.; Kim, D.-I. Editor’s Choice–Comparison of Early Outcomes and Restenosis Rate Between Carotid Endarterectomy and Carotid Artery Stenting Using Propensity Score Matching Analysis. Eur. J. Vasc. Endovasc. Surg. 2017, 54, 573–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Chen, H.; Jin, J.; Tang, Z.; Yin, P.; Zhong, D.; Li, G. Melatonin ameliorates cerebral ischemia/reperfusion injury through SIRT3 activation. Life Sci. 2019, 239, 117036. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Lu, C.; Li, T.; Wang, W.; Ye, W.; Zeng, R.; Ni, L.; Lai, Z.; Wang, X.; Liu, C. The protective effect of melatonin on brain ischemia and reperfusion in rats and humans: In vivo assessment and a randomized controlled trial. J. Pineal Res. 2018, 65, e12521. [Google Scholar] [CrossRef]
- Wei, N.; Pu, Y.; Yang, Z.; Pan, Y.; Liu, L. Therapeutic effects of melatonin on cerebral ischemia reperfusion injury: Role of Yap-OPA1 signaling pathway and mitochondrial fusion. Biomed. Pharmacother. 2019, 110, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Zang, M.; Zhao, Y.; Gao, L.; Zhong, F.; Qin, Z.; Tong, R.; Ai, L.; Petersen, L.; Yan, Y.; Gao, Y.; et al. The circadian nuclear receptor RORα negatively regulates cerebral ischemia–reperfusion injury and mediates the neuroprotective effects of melatonin. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165890. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Chen, T.Y.; Hung, C.Y.; Tai, S.H.; Huang, S.Y.; Chang, C.C.; Hung, H.Y.; Lee, E.J. Melatonin protects brain against ischemia/reperfusion injury by attenuating endoplasmic reticulum stress. Int. J. Mol. Med. 2018, 42, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zhang, L.-Y.; Chen, Y.; Bai, Y.-P.; Jia, J.; Feng, J.-G.; Liu, K.-X.; Zhou, J. Melatonin alleviates intestinal injury, neuroinflammation and cognitive dysfunction caused by intestinal ischemia/reperfusion. Int. Immunopharmacol. 2020, 85, 106596. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.; Chen, Y.; Sung, P.; Chiang, J.Y.; Chu, Y.; Huang, C.; Huang, C.; Yip, H.-K. Hepatic 31 P-magnetic resonance spectroscopy identified the impact of melatonin-pretreated mitochondria in acute liver ischaemia-reperfusion injury. J. Cell. Mol. Med. 2020, 24, 10088–10099. [Google Scholar] [CrossRef]
- Yang, J.; Liu, H.; Han, S.; Fu, Z.; Wang, J.; Chen, Y.; Wang, L. Melatonin pretreatment alleviates renal ischemia-reperfusion injury by promoting autophagic flux via TLR4/MyD88/MEK/ERK/mTORC1 signaling. FASEB J. 2020, 34, 12324–12337. [Google Scholar] [CrossRef]
- Delnevo, C.D.; Gundersen, D.A.; Manderski, M.T.; Giovenco, D.P.; Giovino, G.A. Importance of Survey Design for Studying the Epidemiology of Emerging Tobacco Product Use Among Youth. Am. J. Epidemiol. 2017, 186, 405–410. [Google Scholar] [CrossRef]
- Ding, N.; Sang, Y.; Chen, J.; Ballew, S.H.; Kalbaugh, C.A.; Salameh, M.J.; Blaha, M.J.; Allison, M.; Heiss, G.; Selvin, E.; et al. Cigarette Smoking, Smoking Cessation, and Long-Term Risk of 3 Major Atherosclerotic Diseases. J. Am. Coll. Cardiol. 2019, 74, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Nakano, Y.; Adachi, S.; Murohara, T. Effects of Tobacco Smoking on Cardiovascular Disease. Circ. J. 2019, 83, 1980–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch, J.; Jin, L.; Richardson, A.; Conklin, D.J. Tobacco Smoke and Endothelial Dysfunction: Role of Aldehydes? Curr. Hypertens. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Smith, J.L.; Hodges, R.E. Serum Levels of Vitamin C in Relation to Dietary and Supplemental Intake of Vitamin C in Smokers and Nonsmokers. Ann. N. Y. Acad. Sci. 1987, 498, 144–152. [Google Scholar] [CrossRef]
- Ozguner, F.; Koyu, A.; Cesur, G. Active smoking causes oxidative stress and decreases blood melatonin levels. Toxicol. Ind. Heal. 2005, 21, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Ursing, C.; Von Bahr, C.; Brismar, K.; Röjdmark, S. Influence of cigarette smoking on melatonin levels in man. Eur. J. Clin. Pharmacol. 2005, 61, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Rodella, L.F.; Favero, G.; Rossini, C.; Foglio, E.; Reiter, R.J.; Rezzani, R. Endothelin-1 as a potential marker of melatonin’s therapeutic effects in smoking-induced vasculopathy. Life Sci. 2010, 87, 558–564. [Google Scholar] [CrossRef]
- Wang, Z.; Ni, L.; Wang, J.; Lu, C.; Ren, M.; Han, W.; Liu, C. The protective effect of melatonin on smoke-induced vascular injury in rats and humans: A randomized controlled trial. J. Pineal Res. 2016, 60, 217–227. [Google Scholar] [CrossRef]
- Danaei, G.; Ding, E.L.; Mozaffarian, D.; Taylor, B.; Rehm, J.; Murray, C.J.L.; Ezzati, M. The Preventable Causes of Death in the United States: Comparative Risk Assessment of Dietary, Lifestyle, and Metabolic Risk Factors. PLoS Med. 2009, 6, e1000058. [Google Scholar] [CrossRef]
- Boutouyrie, P.; Tropeano, A.I.; Asmar, R.; Gautier, I.; Benetos, A.; Lacolley, P.; Laurent, S. Aortic Stiffness Is an Independent Predictor of Primary Coronary Events in Hypertensive Patients. Hypertens. 2002, 39, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Zhang, Q.-Y.; Zhang, Y.-L.; Han, X.; Guo, S.-B.; Li, H.-H. Gallic Acid Attenuates Angiotensin II-Induced Hypertension and Vascular Dysfunction by Inhibiting the Degradation of Endothelial Nitric Oxide Synthase. Front. Pharmacol. 2020, 11, 1121. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, X.; Su, C.; Wu, F.; Sun, J.; Zhang, J.; Yang, X.; Zhang, C.; Zhou, Z.; Zhang, X.; et al. Inhibition of Mitochondrial Oxidative Damage Improves Reendothelialization Capacity of Endothelial Progenitor Cells via SIRT3 (Sirtuin 3)-Enhanced SOD2 (Superoxide Dismutase 2) Deacetylation in Hypertension. Arter. Thromb. Vasc. Biol. 2019, 39, 1682–1698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Thompson, A.M.; Tong, W.; Xu, T.; Chen, J.; Zhao, L.; Kelly, T.N.; Chen, C.-S.; He, J. Biomarkers of inflammation and endothelial dysfunction and risk of hypertension among Inner Mongolians in China. J. Hypertens. 2010, 28, 35–40. [Google Scholar] [CrossRef]
- Bellien, J.; Iacob, M.; Remy-Jouet, I.; Lucas, D.; Monteil, C.; Gutierrez, L.; Vendeville, C.; Dreano, Y.; Mercier, A.; Thuillez, C.; et al. Epoxyeicosatrienoic Acids Contribute With Altered Nitric Oxide and Endothelin-1 Pathways to Conduit Artery Endothelial Dysfunction in Essential Hypertension. Circulation 2012, 125, 1266–1275. [Google Scholar] [CrossRef]
- Roca, F.; Bellien, J.; Iacob, M.; Joannides, R. Endothelium-dependent adaptation of arterial wall viscosity during blood flow increase is impaired in essential hypertension. Atheroscler. 2019, 285, 102–107. [Google Scholar] [CrossRef]
- Skrzypczyk, P.; Ozimek, A.; Ofiara, A.; Szyszka, M.; Sołtyski, J.; Stelmaszczyk-Emmel, A.; Górska, E.; Pańczyk-Tomaszewska, M. Markers of endothelial injury and subclinical inflammation in children and adolescents with primary hypertension. Central Eur. J. Immunol. 2019, 44, 253–261. [Google Scholar] [CrossRef]
- Holmes, S.W.; Sugden, D. Proceedings: The effect of melatonin on pinealectomy-induced hypertension in the rat. Br. J. Pharmacol. 1976, 56, 360P–361P. [Google Scholar]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Reiter, R.J. The potential usefulness of serum melatonin level to predict heart failure in patients with hypertensive cardiomyopathy. Int. J. Cardiol. 2014, 174, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Chen, T.; Li, J.; Xiao, J.; Wang, S.; Guo, X.; Bu, P. Correlations of Serum Cyclophilin A and Melatonin Concentrations with Hypertension-induced Left Ventricular Hypertrophy. Arch. Med. Res. 2017, 48, 526–534. [Google Scholar] [CrossRef]
- Ahsanova, E.; Popov, V.; Bulanova, N.; Morozova, T.; Moscow, R. Hypotensive Action of Melatonin in Patients with Arterial Hypertension. Eur. Cardiol. Rev. 2020, 15, e43. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Reiter, R.J.; Paulis, L. Melatonin as a rational alternative in the conservative treatment of resistant hypertension. Hypertens. Res. 2019, 42, 1828–1831. [Google Scholar] [CrossRef] [Green Version]
- Baltatu, O.C.; Amaral, F.G.; Campos, L.A.; Baltatu, O.C. Melatonin, mitochondria and hypertension. Cell. Mol. Life Sci. 2017, 74, 3955–3964. [Google Scholar] [CrossRef] [PubMed]
- Pechanova, O.; Paulis, L.; Simko, F. Peripheral and Central Effects of Melatonin on Blood Pressure Regulation. Int. J. Mol. Sci. 2014, 15, 17920–17937. [Google Scholar] [CrossRef] [Green Version]
- Shao, G.; Zhang, S.; Nie, J.; Li, J.; Tong, J. Effects of melatonin on mechanisms involved in hypertension using human umbilical vein endothelial cells. J. Toxicol. Environ. Heal. Part A 2017, 80, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Jiang, Z. Melatonin attenuates hypertension and oxidative stress in a rat model of L-NAME-induced gestational hypertension. Vasc. Med. 2020, 25, 295–301. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, X.; Liu, M.; Fan, H.; Zheng, H.; Zhang, S.; Rahman, N.; Wołczyński, S.; Kretowski, A.; Li, X. Melatonin inhibits inflammasome-associated activation of endothelium and macrophages attenuating pulmonary arterial hypertension. Cardiovasc. Res. 2020, 116, 2156–2169. [Google Scholar] [CrossRef] [PubMed]
- Nishi, E.E.; Almeida, V.R.; Amaral, F.G.; Simon, K.A.; Futuro-Neto, H.A.; Pontes, R.B.; Cespedes, J.G.; Campos, R.R.; Bergamaschi, C.T. Melatonin attenuates renal sympathetic overactivity and reactive oxygen species in the brain in neurogenic hypertension. Hypertens. Res. 2019, 42, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Pechanova, O.; Bednarova, K.R.; Krajcirovicova, K.; Celec, P.; Kamodyova, N.; Zorad, S.; Kucharska, J.; Gvozdjakova, A.; Adamcova, M.; et al. Hypertension and Cardiovascular Remodelling in Rats Exposed to Continuous Light: Protection by ACE-Inhibition and Melatonin. Mediat. Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Astorga, C.R.; Gonzaléz-Candia, A.; Candia, A.A.; Figueroa, E.G.; Cañas, D.; Ebensperger, G.; Reyes, R.V.; Llanos, A.J.; Herrera, E. Melatonin Decreases Pulmonary Vascular Remodeling and Oxygen Sensitivity in Pulmonary Hypertensive Newborn Lambs. Front. Physiol. 2018, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Renna, N.F.; Heras, N.D.L.; Miatello, R.M. Pathophysiology of Vascular Remodeling in Hypertension. Int. J. Hypertens. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dinh, Q.N.; Chrissobolis, S.; Diep, H.; Chan, C.T.; Ferens, D.; Drummond, G.R.; Sobey, C.G. Advanced atherosclerosis is associated with inflammation, vascular dysfunction and oxidative stress, but not hypertension. Pharmacol. Res. 2017, 116, 70–76. [Google Scholar] [CrossRef]
- Last, A.R.; Ference, J.D.; Menzel, E.R. Hyperlipidemia: Drugs for Cardiovascular Risk Reduction in Adults. Am. Fam. Physician 2017, 95, 78–87. [Google Scholar] [PubMed]
- Li, C.-Y.; Ma, L.; Yu, B. Circular RNA hsa_circ_0003575 regulates oxLDL induced vascular endothelial cells proliferation and angiogenesis. Biomed. Pharmacother. 2017, 95, 1514–1519. [Google Scholar] [CrossRef]
- Cho, K.; Choi, S.H. ASK1 Mediates Apoptosis and Autophagy during oxLDL-CD36 Signaling in Senescent Endothelial Cells. Oxidative Med. Cell. Longev. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Zeng, Z.; Jiaojiao, C.; Peng, W.; Yami, L.; Tingting, Z.; Jun, T.; Shiyuan, W.; Jinyan, X.; Dangheng, W.; Zhisheng, J.; et al. OxLDL induces vascular endothelial cell pyroptosis through miR-125a-5p/TET2 pathway. J. Cell. Physiol. 2018, 234, 7475–7491. [Google Scholar] [CrossRef]
- Lee, W.-J.; Ou, H.-C.; Hsu, W.-C.; Chou, M.-M.; Tseng, J.-J.; Hsu, S.-L.; Tsai, K.-L.; Sheu, W.H.-H. Ellagic acid inhibits oxidized LDL-mediated LOX-1 expression, ROS generation, and inflammation in human endothelial cells. J. Vasc. Surg. 2010, 52, 1290–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, L.; Peng, Y.; Xiang, R.; Li, X.; Li, Z. Ox-LDL Causes Endothelial Cell Injury Through ASK1/NLRP3-Mediated Inflammasome Activation via Endoplasmic Reticulum Stress. Drug Des. Dev. Ther. 2020, 14, 731–744. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Zheng, Q.; Chen, J.; Tan, X.; Li, Q.; Ding, L.; Zhang, R.; Xiaolong, L. FGF21 mitigates atherosclerosis via inhibition of NLRP3 inflammasome-mediated vascular endothelial cells pyroptosis. Exp. Cell Res. 2020, 393, 112108. [Google Scholar] [CrossRef]
- The AIM-HIGH Investigators Niacin in Patients with Low HDL Cholesterol Levels Receiving Intensive Statin Therapy. N. Engl. J. Med. 2011, 365, 2255–2267. [CrossRef] [PubMed] [Green Version]
- Kypreos, K.E.; Gkizas, S.; Rallidis, L.S.; Karagiannides, I. HDL particle functionality as a primary pharmacological target for HDL-based therapies. Biochem. Pharmacol. 2013, 85, 1575–1578. [Google Scholar] [CrossRef]
- Ouimet, M.; Barrett, T.J.; Fisher, A.E. HDL and Reverse Cholesterol Transport. Circ. Res. 2019, 124, 1505–1518. [Google Scholar] [CrossRef]
- Xepapadaki, E.; Zvintzou, E.; Kalogeropoulou, C.; Filou, S.; Kypreos, K.E. Τhe Antioxidant Function of HDL in Atherosclerosis. Angiol. 2019, 71, 112–121. [Google Scholar] [CrossRef]
- Hu, J.; Xi, D.; Zhao, J.; Luo, T.; Liu, J.; Lu, H.; Li, M.; Xiong, H.; Guo, Z. High-density Lipoprotein and Inflammation and Its Significance to Atherosclerosis. Am. J. Med. Sci. 2016, 352, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-González, P.; Garcia-Gonzalez, M.; Hita, J.J.F.; Vargas, M.; Reiter, R.J. Elevated levels of oxidized low-density lipoprotein and impaired nocturnal synthesis of melatonin in patients with myocardial infarction. Atherosclerosis 2005, 180, 101–105. [Google Scholar] [CrossRef]
- Hussain, S.A.-R. Effect of melatonin on cholesterol absorption in rats. J. Pineal Res. 2007, 42, 267–271. [Google Scholar] [CrossRef]
- Kitagawa, A.; Ohta, Y.; Ohashi, K. Melatonin improves metabolic syndrome induced by high fructose intake in rats. J. Pineal Res. 2012, 52, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, H.; Mori, N.; Mori, W. Effects of melatonin on genetic hypercholesterolemia in rats. Atherosclerosis 1988, 69, 269–272. [Google Scholar] [CrossRef]
- Mohammadi-Sartang, M.; Ghorbani, M.; Mazloom, Z. Effects of melatonin supplementation on blood lipid concentrations: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2018, 37, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Mandala, A.; Naaz, S.; Giri, S.; Jain, M.; Bandyopadhyay, D.; Reiter, R.J.; Roy, S.S. Melatonin protects against lipid-induced mitochondrial dysfunction in hepatocytes and inhibits stellate cell activation during hepatic fibrosis in mice. J. Pineal Res. 2017, 62, e12404. [Google Scholar] [CrossRef]
- Mi, Y.; Tan, D.; He, Y.; Zhou, X.; Zhou, Q.; Ji, S. Melatonin Modulates lipid Metabolism in HepG2 Cells Cultured in High Concentrations of Oleic Acid: AMPK Pathway Activation may Play an Important Role. Cell Biophys. 2018, 76, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsson, B.G.; Wikström, J.; Jakubowicz, R.; Marmon, S.K.; Carlsson, N.-G.; Jansson, N.; Undeland, I.; Holmäng, A.; Gan, L.-M.; Lönn, M.; et al. Dietary herring improves plasma lipid profiles and reduces atherosclerosis in obese low-density lipoprotein receptor-deficient mice. Int. J. Mol. Med. 2011, 29, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Lu, C.; Zhao, W.; Shao, X. Melatonin attenuates TNF-α-mediated hepatocytes damage via inhibiting mitochondrial stress and activating the Akt-Sirt3 signaling pathway. J. Cell. Physiol. 2019, 234, 20969–20979. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Bai, X.; Lin, Y.; Li, Z.; Fu, J.; Li, M.; Zhao, T.; Yang, H.; Xu, R.; et al. Melatonin prevents endothelial cell pyroptosis via regulation of long noncoding RNA MEG3/miR-223/NLRP3 axis. J. Pineal Res. 2017, 64, e12449. [Google Scholar] [CrossRef]
- Ma, S.; Chen, J.; Feng, J.; Zhang, R.; Fan, M.; Han, D.; Li, X.; Li, C.; Ren, J.; Wang, Y.; et al. Melatonin Ameliorates the Progression of Atherosclerosis via Mitophagy Activation and NLRP3 Inflammasome Inhibition. Oxidative Med. Cell. Longev. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Djordjevic, B.; Cvetkovic, T.; Stoimenov, T.J.; Despotovic, M.; Zivanovic, S.; Basic, J.; Veljkovic, A.; Velickov, A.; Kocic, G.; Pavlovic, D.; et al. Oral supplementation with melatonin reduces oxidative damage and concentrations of inducible nitric oxide synthase, VEGF and matrix metalloproteinase 9 in the retina of rats with streptozotocin/nicotinamide induced pre-diabetes. Eur. J. Pharmacol. 2018, 833, 290–297. [Google Scholar] [CrossRef]
- Dos Santos, R.M.; Marani, F.; Chiba, F.Y.; Mattera, M.S.D.L.C.; Tsosura, T.V.S.; Tessarin, G.W.L.; Pereira, R.F.; Belardi, B.E.; Pinheiro, B.C.E.S.; Matsushita, D. Melatonin promotes reduction in TNF levels and improves the lipid profile and insulin sensitivity in pinealectomized rats with periodontal disease. Life Sci. 2018, 213, 32–39. [Google Scholar] [CrossRef]
- Hagensen, M.K.; Mortensen, M.B.; Kjolby, M.; Palmfeldt, J.; Bentzon, J.F.; Gregersen, S. Increased retention of LDL from type 1 diabetic patients in atherosclerosis-prone areas of the murine arterial wall. Atherosclerosis 2019, 286, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, E.; Jha, J.C.; Sharma, A.; Wilkinson-Berka, J.L.; Jandeleit-Dahm, K.A.; De Haan, J.B. Are reactive oxygen species still the basis for diabetic complications? Clin. Sci. 2015, 129, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Morieri, M.L.; Longato, E.; Mazzucato, M.; Di Camillo, B.; Cocchiglia, A.; Gubian, L.; Sparacino, G.; Avogaro, A.; Fadini, G.P.; De Kreutzenberg, S.V. Improved long-term cardiovascular outcomes after intensive versus standard screening of diabetic complications: An observational study. Cardiovasc. Diabetol. 2019, 18, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, T.; Deedwania, P.C. Cardiovascular outcome trials of the newer anti-diabetic medications. Prog. Cardiovasc. Dis. 2019, 62, 342–348. [Google Scholar] [CrossRef]
- Tian, J.; Liu, Y.; Liu, Y.; Chen, K.; Lyu, S. Cellular and Molecular Mechanisms of Diabetic Atherosclerosis: Herbal Medicines as a Potential Therapeutic Approach. Oxidative Med. Cell. Longev. 2017, 2017, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katakami, N. Mechanism of Development of Atherosclerosis and Cardiovascular Disease in Diabetes Mellitus. J. Atheroscler. Thromb. 2018, 25, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerhill, I.V.; Grechko, A.V.; Yet, S.-F.; Sobenin, I.A.; Ivanova, E. Yet The Atherogenic Role of Circulating Modified Lipids in Atherosclerosis. Int. J. Mol. Sci. 2019, 20, 3561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, A.; Maestroni, G.J. Role of the pineal gland and melatonin in the development of autoimmune diabetes in non-obese diabetic mice. J. Pineal Res. 1996, 20, 164–172. [Google Scholar] [CrossRef]
- Conti, A.; Maestroni, G.J.M. Melatonin Rhythms in Mice: Role in Autoimmune and Lymphoproliferative Diseases. Pineal Gland Cancer 2001, 840, 395–407. [Google Scholar] [CrossRef]
- Teodoro, B.G.; Baraldi, F.G.; Sampaio, I.H.; Bomfim, L.H.M.; Queiroz, A.L.; Passos, M.A.; Carneiro, E.M.; Alberici, L.C.; Gomis, R.; Amaral, F.G.D.; et al. Melatonin prevents mitochondrial dysfunction and insulin resistance in rat skeletal muscle. J. Pineal Res. 2014, 57, 155–167. [Google Scholar] [CrossRef]
- Bonnefond, A.; Clément, N.A.; Fawcett, K.; Yengo, L.; Vaillant, E.; Guillaume, J.-L.; Dechaume, A.; Payne, F.; Roussel, R.; The Meta-Analysis of Glucose and Insulin-Related Traits Consortium (MAGIC); et al. Rare MTNR1B variants impairing melatonin receptor 1B function contribute to type 2 diabetes. Nat. Genet. 2012, 44, 297–301. [Google Scholar] [CrossRef]
- Sartori, C.; Dessen, P.; Mathieu, C.; Monney, A.; Bloch, J.; Nicod, P.; Scherrer, U.; Duplain, H. Melatonin Improves Glucose Homeostasis and Endothelial Vascular Function in High-Fat Diet-Fed Insulin-Resistant Mice. Endocrinology 2009, 150, 5311–5317. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Ni, L.; Zhao, Z.; Liu, X.; Lai, Z.; Di, X.; Xie, Z.; Song, X.; Wang, X.; Zhang, R.; et al. Melatonin attenuates smoking-induced hyperglycemia via preserving insulin secretion and hepatic glycogen synthesis in rats. J. Pineal Res. 2018, 64, e12475. [Google Scholar] [CrossRef] [Green Version]
- Agil, A.; Navarro-Alarcón, M.; Ruiz, R.; Abuhamadah, S.; El-Mir, M.-Y.; Vázquez, G.F. Beneficial effects of melatonin on obesity and lipid profile in young Zucker diabetic fatty rats. J. Pineal Res. 2010, 50, 207–212. [Google Scholar] [CrossRef]
- Amin, A.H.; El-Missiry, M.A.; Othman, A.I. Melatonin ameliorates metabolic risk factors, modulates apoptotic proteins, and protects the rat heart against diabetes-induced apoptosis. Eur. J. Pharmacol. 2015, 747, 166–173. [Google Scholar] [CrossRef]
- Xiong, F.; Tang, S.; Su, H.; Tang, H.; Jiang, P.; Zhou, Q.; Wang, Y.; Zhu, H. Melatonin ameliorates myocardial apoptosis by suppressing endoplasmic reticulum stress in rats with long-term diabetic cardiomyopathy. Mol. Med. Rep. 2017, 17, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Che, H.; Wang, Y.; Li, H.; Li, Y.; Sahil, A.; Lv, J.; Liu, Y.; Yang, Z.; Dong, R.; Xue, H.; et al. Melatonin alleviates cardiac fibrosis via inhibiting lncRNA MALAT1/miR-141-mediated NLRP3 inflammasome and TGF-β1/Smads signaling in diabetic cardiomyopathy. FASEB J. 2020, 34, 5282–5298. [Google Scholar] [CrossRef]
- De Melo, I.M.F.; Ferreira, C.G.M.; Souza, E.H.L.D.S.; Almeida, L.L.; De Sá, F.B.; Neto, C.J.C.L.; De Castro, M.V.P.; Teixeira, V.W.; Teixeira-Álvaro, A.C. Melatonin regulates the expression of inflammatory cytokines, VEGF and apoptosis in diabetic retinopathy in rats. Chem. Interact. 2020, 327, 109183. [Google Scholar] [CrossRef]
- Hamczyk, M.R.; Nevado, R.M.; Barettino, A.; Fuster, V.; Andrés, V. Biological Versus Chronological Aging. J. Am. Coll. Cardiol. 2020, 75, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Iurciuc, S.; Cimpean, A.; Mitu, F.; Heredea, R.; Iurciuc, M. Vascular aging and subclinical atherosclerosis: Why such a “never ending” and challenging story in cardiology? Clin. Interv. Aging 2017, 12, 1339–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, F.-Y.; Sun, C.-K.; Sung, P.-H.; Chen, K.-H.; Chua, S.; Sheu, J.-J.; Chung, S.-Y.; Chai, H.-T.; Chen, Y.-L.; Huang, T.-H.; et al. Daily melatonin protects the endothelial lineage and functional integrity against the aging process, oxidative stress, and toxic environment and restores blood flow in critical limb ischemia area in mice. J. Pineal Res. 2018, 65, e12489. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Sinclair, D.A. The Intersection between Aging and Cardiovascular Disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef]
- Oliveira, H.C.; Vercesi, A.E. Mitochondrial bioenergetics and redox dysfunctions in hypercholesterolemia and atherosclerosis. Mol. Asp. Med. 2020, 71, 100840. [Google Scholar] [CrossRef]
- Barja, G. The Mitochondrial Free Radical Theory of Aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar] [CrossRef]
- Herrmann, W.; Herrmann, M. The Importance of Telomere Shortening for Atherosclerosis and Mortality. J. Cardiovasc. Dev. Dis. 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of Dysfunction in the Aging Vasculature and Role in Age-Related Disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-K.; Hsu, P.-C.; Tsai, W.-C.; Chen, Y.-C.; Lee, H.-H.; Lee, W.-H.; Chu, C.-Y.; Lee, C.-S.; Yen, H.-W.; Lin, T.-H.; et al. Gender differences in major adverse cardiovascular outcomes among aged over 60 year-old patients with atherosclerotic cardiovascular disease. Medicine 2020, 99, e19912. [Google Scholar] [CrossRef]
- Tamarindo, G.H.; Gobbo, M.G.; Taboga, S.R.; Almeida, E.A.; Góes, R.M. Melatonin ameliorates degenerative alterations caused by age in the rat prostate and mitigates high-fat diet damages. Cell Biol. Int. 2021, 45, 92–106. [Google Scholar] [CrossRef]
- Igarashi-Migitaka, J.; Seki, A.; Ikegame, M.; Honda, M.; Sekiguchi, T.; Mishima, H.; Shimizu, N.; Matsubara, H.; Srivastav, A.K.; Hirayama, J.; et al. Oral administration of melatonin contained in drinking water increased bone strength in naturally aged mice. Acta Histochem. 2020, 122, 151596. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Kawamoto, M.; Sato, S.; Tamura, I.; Maekawa, R.; Taketani, T.; Aasada, H.; Takaki, E.; Nakai, A.; Reiter, R.J.; et al. Long-term melatonin treatment delays ovarian aging. J. Pineal Res. 2016, 62, e12381. [Google Scholar] [CrossRef]
- Jauhari, A.; Baranov, S.V.; Suofu, Y.; Kim, J.; Singh, T.; Yablonska, S.; Li, F.; Wang, X.; Oberly, P.; Minnigh, M.B.; et al. Melatonin inhibits cytosolic mitochondrial DNA–induced neuroinflammatory signaling in accelerated aging and neurodegeneration. J. Clin. Investig. 2020, 130, 3124–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrzadi, S.; Hemati, K.; Reiter, R.J.; Hosseinzadeh, A. Mitochondrial dysfunction in age-related macular degeneration: Melatonin as a potential treatment. Expert Opin. Ther. Targets 2020, 24, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wu, C.; Muhammad, J.S.; Yan, D.; Tsuneyama, K.; Hatta, H.; Cui, Z.-G.; Inadera, H. Melatonin sensitises shikonin-induced cancer cell death mediated by oxidative stress via inhibition of the SIRT3/SOD2-AKT pathway. Redox Biol. 2020, 36, 101632. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, X.; Wang, G.; Chen, L.; Yang, H.; He, F.; Lin, J. Melatonin Rescues the Ti Particle-Impaired Osteogenic Potential of Bone Marrow Mesenchymal Stem Cells via the SIRT1/SOD2 Signaling Pathway. Calcif. Tissue Int. 2020, 107, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Volt, H.; García, J.A.; Doerrier, C.; Díaz-Casado, M.E.; Guerra-Librero, A.; López, L.C.; Escames, G.; Tresguerres, J.A.; Acuña-Castroviejo, D. Same molecule but different expression: Aging and sepsis trigger NLRP3 inflammasome activation, a target of melatonin. J. Pineal Res. 2016, 60, 193–205. [Google Scholar] [CrossRef]
- Hardeland, R. Aging, Melatonin, and the Pro- and Anti-Inflammatory Networks. Int. J. Mol. Sci. 2019, 20, 1223. [Google Scholar] [CrossRef] [Green Version]
- Rodella, L.F.; Favero, G.; Rossini, C.; Foglio, E.; Bonomini, F.; Reiter, R.J.; Rezzani, R. Aging and vascular dysfunction: Beneficial melatonin effects. AGE 2011, 35, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Martino, T.A.; Tata, N.; Belsham, D.D.; Chalmers, J.; Straume, M.; Lee, P.; Pribiag, H.; Khaper, N.; Liu, P.P.; Dawood, F.; et al. Disturbed Diurnal Rhythm Alters Gene Expression and Exacerbates Cardiovascular Disease with Rescue by Resynchronization. Hypertension 2007, 49, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Vujovic, N.; Williams, J.S.; Scheer, F.A. Impact of Circadian Disruption on Cardiovascular Function and Disease. Trends Endocrinol. Metab. 2019, 30, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Cai, A.; Zhong, Q.; Liu, C.; Zhou, D.; Li, X.; Zhang, Y.; Feng, Y.; Zhou, Y. Associations of systolic and diastolic blood pressure night-to-day ratios with atherosclerotic cardiovascular diseases. Hypertens. Res. 2016, 39, 874–878. [Google Scholar] [CrossRef]
- Tang, H.; Zhu, M.; Zhao, G.; Fu, W.; Shi, Z.; Ding, Y.; Tang, X.; Guo, D. Loss of CLOCK under high glucose upregulates ROCK1-mediated endothelial to mesenchymal transition and aggravates plaque vulnerability. Atherosclerosis 2018, 275, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Xue, S.; Zhao, G.; Fang, C.; Cai, L.; Shi, Z.; Fu, W.; Qian, R.; Zhang, P.; Tang, X.; et al. CLOCK Disruption Ag-gravates Carotid Artery Stenosis through Endoplasmic Reticulum Stress-Induced Endothelial-Mesenchymal Transition. Am. J. Transl. Res. 2020, 12, 7885–7898. [Google Scholar] [PubMed]
- Yang, G.; Chen, L.; Grant, G.R.; Paschos, G.; Song, W.-L.; Musiek, E.S.; Lee, V.; McLoughlin, S.C.; Grosser, T.; Cotsarelis, G.; et al. Timing of expression of the core clock gene Bmal1 influences its effects on aging and survival. Sci. Transl. Med. 2016, 8, 324ra16. [Google Scholar] [CrossRef] [Green Version]
- Takaguri, A.; Sasano, J.; Akihiro, O.; Satoh, K. The role of circadian clock gene BMAL1 in vascular proliferation. Eur. J. Pharmacol. 2020, 872, 172924. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, K.; Yamagami, Y.; Tatsumi, S.; Kurumatani, N.; Saeki, K. Indoor light pollution and progression of carotid atherosclerosis: A longitudinal study of the HEIJO-KYO cohort. Environ. Int. 2019, 133, 105184. [Google Scholar] [CrossRef] [PubMed]
- Imbesi, M.; Arslan, A.D.; Yildiz, S.; Sharma, R.; Gavin, D.; Tun, N.; Manev, H.; Uz, T. The melatonin receptor MT1 is required for the differential regulatory actions of melatonin on neuronal ‘clock’ gene expression in striatal neurons in vitro. J. Pineal Res. 2009, 46, 87–94. [Google Scholar] [CrossRef]
- Renzi, A.; Glaser, S.; DeMorrow, S.; Mancinelli, R.; Meng, F.; Franchitto, A.; Venter, J.; White, M.; Francis, H.; Han, Y.; et al. Melatonin inhibits cholangiocyte hyperplasia in cholestatic rats by interaction with MT1 but not MT2 melatonin receptors. Am. J. Physiol. Liver Physiol. 2011, 301, G634–G643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, M.; Rauch, A.; Korf, H.-W.; Von Gall, C. The Endogenous Melatonin (MT) Signal Facilitates Reentrainment of the Circadian System to Light-Induced Phase Advances by Acting Upon MT2 Receptors. Chrono Int. 2012, 29, 415–429. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Romo-Nava, F.; Buijs, F.N.; Valdés-Tovar, M.; Benítez-King, G.; Basualdo, M.; Perusquía, M.; Heinze, G.; Escobar, C.; Buijs, R.M. Olanzapine-induced early cardiovascular effects are mediated by the biological clock and prevented by melatonin. J. Pineal Res. 2017, 62, e12402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, D.I.; González-Fernández, B.; Crespo, I.; San-Miguel, B.; Álvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin modulates dysregulated circadian clocks in mice with diethylnitrosamine-induced hepatocellular carcinoma. J. Pineal Res. 2018, 65, e12506. [Google Scholar] [CrossRef]
- Rahim, I.; Djerdjouri, B.; Sayed, R.K.; Fernández-Ortiz, M.; Fernández-Gil, B.; Hidalgo-Gutiérrez, A.; López, L.C.; Escames, G.; Reiter, R.J.; Acuña-Castroviejo, D. Melatonin administration to wild-type mice and nontreated NLRP3 mutant mice share similar inhibition of the inflammatory response during sepsis. J. Pineal Res. 2017, 63, e12410. [Google Scholar] [CrossRef]
- Peschke, E.; Bähr, I.; Mühlbauer, E. Experimental and clinical aspects of melatonin and clock genes in diabetes. J. Pineal Res. 2015, 59, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R. Melatonin and inflammation-Story of a double-edged blade. J. Pineal Res. 2018, 65, e12525. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Vico, A.; Lardone, P.J.; Álvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: Buffering the Immune System. Int. J. Mol. Sci. 2013, 14, 8638–8683. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Rong, Z.; Liu, C. Potential Role of Melatonin as an Adjuvant for Atherosclerotic Carotid Arterial Stenosis. Molecules 2021, 26, 811. https://doi.org/10.3390/molecules26040811
Zhang R, Ni L, Di X, Ma B, Niu S, Rong Z, Liu C. Potential Role of Melatonin as an Adjuvant for Atherosclerotic Carotid Arterial Stenosis. Molecules. 2021; 26(4):811. https://doi.org/10.3390/molecules26040811
Chicago/Turabian StyleZhang, Rui, Leng Ni, Xiao Di, Baitao Ma, Shuai Niu, Zhihua Rong, and Changwei Liu. 2021. "Potential Role of Melatonin as an Adjuvant for Atherosclerotic Carotid Arterial Stenosis" Molecules 26, no. 4: 811. https://doi.org/10.3390/molecules26040811