
Determination of Biogenic Amines in Different Parts of Lycium barbarum L. by HPLC with Precolumn Dansylation

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Preparation of Samples
2.2. Derivatization Procedure
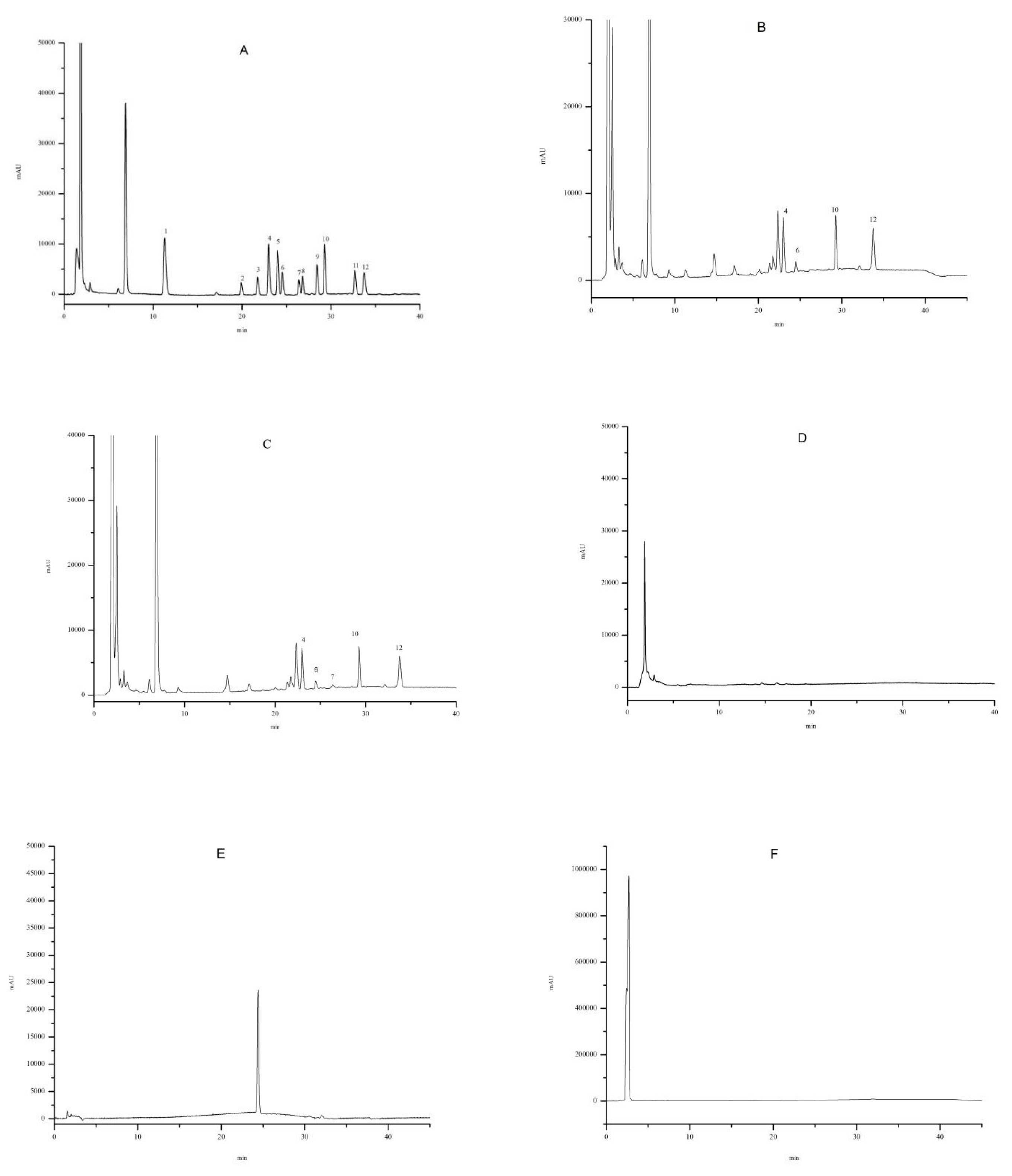
2.3. Identification of the BAs in L. barbarum
2.4. The Matrix Effect
2.5. Method Validation
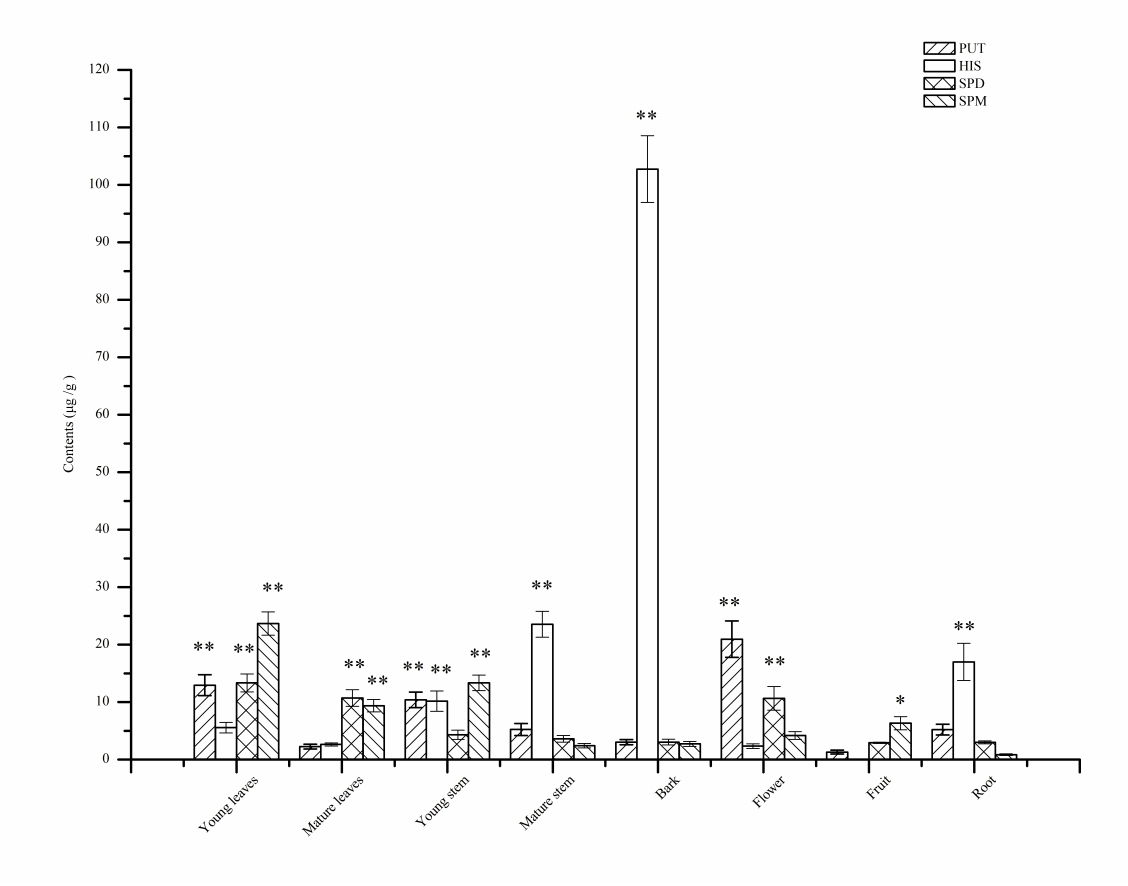
2.6. Distribution of the BAs in L. barbarum
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Preparation of Standard Solutions
3.3. Preparation of Sample Solution
3.4. Derivatization Procedure
3.5. Instrumentation and Conditions
3.6. Method Validation
3.7. Evaluation of the Matrix Effect
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviation
References
- Silla Santos, M.H. Biogenic amines: Their importance in foods. Int. J Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Halász, A.; Baráth, Á.; Simon-Sarkadi, L.; Holzapfel, W. Biogenic amines and their production by microorganisms in food. Trends Food Sci. Technol. 1994, 5, 42–49. [Google Scholar] [CrossRef]
- Önal, A. A review: Current analytical methods for the determination of biogenic amines in foods. Food Chem. 2007, 103, 1475–1486. [Google Scholar] [CrossRef]
- Lovaas, E. Antioxidative effects of polyamines. J. Am. Oil Chem.’ Soc. 1991, 68, 353–358. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- He, E.P.; Tang, L.L.; Guo, Y.J. Effects of spermidine on free radical metabolism and anti-fatigue effects of skeletal muscle in mice. Food Sci. 2014, 35, 229–232. [Google Scholar]
- Wei, C.; Wang, Y.; Li, M.; Li, H.; Lu, X.; Shao, H.; Xu, C. Spermine inhibits endoplasmic reticuum stress-induced apoptosis: A new strategy to prevent cardiomyo-cyte apoptosis. Cell. Physiol. Biochem. 2016, 38, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Thomas, K.; Wittmann, C. Comparison of a capillary electrophoresis method with high-performance liquid chromatography for the determination of biogenic amines in various food samples. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2002, 779, 229–239. [Google Scholar] [CrossRef]
- Becker, K.; Southwick, K.; Reardon, J.; Berg, R.; MacCormack, J.N. Histamine poisoning associated with eating tuna burgers. JAMA 2001, 285, 1327–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, H.M.L.J. The biogenic amine contents of Dutch cheese and their toxicological significance. Neth. Milk Dairy J. 1988, 42, 25–42. [Google Scholar]
- Tang, T.; Qian, K.; Shi, T.; Wang, F.; Li, J.; Cao, Y.; Hu, Q. Monitoring the contents of biogenic amines in sufu by HPLC with SPE and pre-column derivatization. Food Control. 2011, 22, 1203–1208. [Google Scholar] [CrossRef]
- Saaid, M.; Saad, B.; Ali, A.S.M.; Saleh, M.I.; Basheer, C.; Lee, H.K. In situ derivatization hollow fibre liquid-phase microextraction for the determination of biogenic amines in food samples. J. Chromatogr. A 2009, 1216, 5165–5170. [Google Scholar] [CrossRef] [PubMed]
- Saaid, M.; Saad, B.; Hashim, N.H.; Ali, A.S.M.; Saleh, M.I. Determination of biogenic amines in selected Malaysian food. Food Chem. 2009, 113, 1356–1362. [Google Scholar] [CrossRef]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Valsamaki, K.; Michaelidou, A.; Polychroniadou, A. Biogenic amine production in Feta cheese. Food Chem. 2000, 71, 259–266. [Google Scholar] [CrossRef]
- People’s Medical Publishing House. Lishizhen Compendium of materia medica: On the university handbook, next volume; People’s Medical Publishing House: Beijing, China, 1982; pp. 2111–2117. [Google Scholar]
- Zhao, J.; Jin, Y.; Yan, Y.; Qin, K.; Qian, D.; Zhang, W.; Peng, H.; Li, B.; Cai, Q.; Chen, Z.; et al. Herbal Textual Research on ″Lycii Fructus″ and ″Lycii Cortex″ in Chinese Classical Prescriptions. Modern Chin. Med. 2020, 22, 1269–1286. [Google Scholar]
- Lu, A.M.; Wang, M.L. On the identification of the original plants in the modernization of Chinese herbal medicine - an example from the taxonomy and exploition of ‘gouqi’. Acta Bot. Boreal.-Occid. Sin. 2003, 23, 1077–1083, (in Chinese English abstract). [Google Scholar]
- Zhao, J.; Ge, L.Y.; Xiong, W.; Leong, F.; Huang, L.Q.; Li, S.P. Advanced development in phytochemicals analysis of medicine and food dual purposes plants used in China (2011–2014). J. Chromatogr. A. 2016, 1428, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Ma, D.; Cheng, Y.; Tian, X.; Lu, Y.; Du, X.; Tang, H.; Chen, J. Three new dimers and two monomers of phenolic amides from the fruits of Lycium barbarum and their antioxidant activities. Agric. Food Chem. 2015, 63, 1067–1075. [Google Scholar] [CrossRef]
- Amagase, H.; Farnsworth, N.R. A reviews of botanical characteristics phytochemistry, clinical relevance in efficacy and safety of Lycium barbarum fruit (Goji). Food Res. Int. 2011, 44, 1702–1717. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A. Goji Berry (Lycium barbarum): Composition and Health Effects-a Review. Nephron Clin. Pract. 2016, 66, 67–76. [Google Scholar]
- Zhao, X. Study on Resource Chemistry of Lycium Barbarum Leaves and Flowers. Master Dissertation, Nanjing University of Chinese Medicine, Nanjing, China, 2020. [Google Scholar]
- Chen, Y.; Tan, F.; Peng, Y. Research Progress in Leaves of Lycium chinese Mill and Lycium barbarum L. Chin. Pharm. J. 2017, 52, 358–361. [Google Scholar]
- Guo, M.; Zhao, J.; Shi, W.; Xu, F.; Cong, W. Advances of Gouqi (Lycium) in Research of Anti-Aging. Guid. J. Tradit. Chin. Med. Pharm. 2019, 25, 124–128. [Google Scholar]
- Yan, X.M.; Dong, J.Z.; Wang, Y. Comparision Studies of Main Active Compounds in young leaves of L. barbarum. Ningxia Food Sci. 2010, 31, 29–32. [Google Scholar]
- Feng, W. Research Status and Trends of Cactus and Lycium barbarum Leaves Applied to Hypoglycemic Health Drink. Food Sci. Technol. Res. 2018, 1, 136. [Google Scholar]
- Gu, J.Q. Preparation of Compound Healthy Beverage from Garlic and Medlar. Food Sci. Technol. 2002, 10, 46. [Google Scholar]
- Gan, X.; Zhang, Y.X. Development of fermented Tea Fermented Beverage. CCOC—China Chamber of Commerce Industry 1998, 10, 12–13. [Google Scholar]
- Li, H.Y.; Wu, D. Comparison of Trace Element Contents in Tea Leaves of Ningxia with Other Teas. Stud. Trace Elem. Health 2008, 25, 37–38. [Google Scholar]
- Ma, B.L.; Qu, X.B. Determination of spermine and spermidine in wolfberry leaves by HPLC. Ningxia Eng. Technol. 2011, 10, 48–50. [Google Scholar]
- Draisci, R.; Volpe, P.G.; Lucentini, O.L.; Cecilia, A.; Federico, R.; Palleschi, G. Determination of biogenic amines with an electrochemical biosensor and its application to salted anchovies. Food Chem. 1998, 62, 225–232. [Google Scholar] [CrossRef]
- Shalaby, A.R. Multidetection semiquantitative method for determining biogenic amines in foods. Food Chem. 1995, 52, 367–372. [Google Scholar] [CrossRef]
- Antoine, F.R.; Wei, C.I.; Otwell, W.S.; Sims, C.A.; Littell, R.C.; Hogle, A.D.; Marshall, M.R. Gas chromatographic analysis of histamine in mahi-mahi (Coryohaena hippurus). J. Agric. Food Chem. 2002, 50, 4754–4759. [Google Scholar] [CrossRef]
- Draisci, R.; Giannetti, L.; Boria, P.; Lucentini, L.; Palleschi, L.; Cavalli, S. Improved ion chromatography-integrated pulsed amperometric detection method for the evaluation of biogenic amines in food of vegetable or animal origin and in fermented foods. J. Chromatogr. A 1998, 798, 109–116. [Google Scholar] [CrossRef]
- Vinci, G.; Antonelli, M.L. Biogenic amine: Quality index of freshness in red and white meat. Food Control. 2002, 13, 519–524. [Google Scholar] [CrossRef]
- De Mey, E.; Drabik-Markiewicz, G.; De Maere, H.; Peeters, M.C.; Derdelinckx, G.; Paelinck, H.; Kowalska, T. Dabsyl derivatisation as an alternative for dansylation in the detection of biogenic amines in fermented meat products by reversed phase high performance liquid chromatography. Food Chem. 2012, 130, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Buiatti, S.; Boschelle, O.; Mozzon, M.; Battistutta, F. Determination of biogenic amines in alcoholic and non-alcoholic beers by HPLC. Food Chem. 1995, 52, 199–202. [Google Scholar] [CrossRef]
- Anlı, R.E.; Vural, N.; Yılmaz, S.; Vural, Ỳ.H. The determination of biogenic amines in Turkish red wines. J. Food Compos. Anal. 2004, 17, 53–62. [Google Scholar]
- Zhang, N.; Wang, Y.Q.; Wang, X.N.; Fan, M.Q.; Sun, Y.N.; Liu, Z.L.; Liu, J.L. Identification and determination of biogenic amines in nine kinds of Chinese Tonic-Qi herbs by RP-HPLC with pre-column derivatization. Chin. J. Pharm. Anal. 2017, 37, 1791–1798. [Google Scholar]
- Fan, M.; Ai, Y.; Zhao, W.; Sun, Y.; Liu, J.; Zhang, N. Characterization of 10 biogenic amines in Male Silkworm Moth by HPLC with precolumn derivatization. Curr. Pharm. Anal. 2019, 16, 608–614. [Google Scholar] [CrossRef]
- Wang, X.; Liang, Y.; Wang, Y.; Fan, M.; Sun, Y.; Liu, J.; Zhang, N. Simultaneous determination of 10 kinds of biogenic amines in rat plasma using high-performance liquid chromatography coupled with fluorescence detection. Biomed. Chromatogr. 2018, 32, e4211. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.I.A.; Moreno, J.L.F.; Frenich, A.G.; Vidal, J.L.M. Fast determination of myo-inositol in milk powder by ultra high performance liquid chromatography coupled to tandem mass spectrometry. Food Chem. 2011, 129, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Kang, Y.P.; Park, J.H.; Lee, J.; Kwon, S.W. Simultaneous determination of 23 amino acids and 7 biogenic amines in fermented food samples by liquid chromatography/quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 9174–9182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chai, B.; Liu, J.; Jiang, P.; Zhao, D. Current Situation and Trend of Drying Way of Wolfberry. Agric. Eng. 2020, 10, 46–49. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
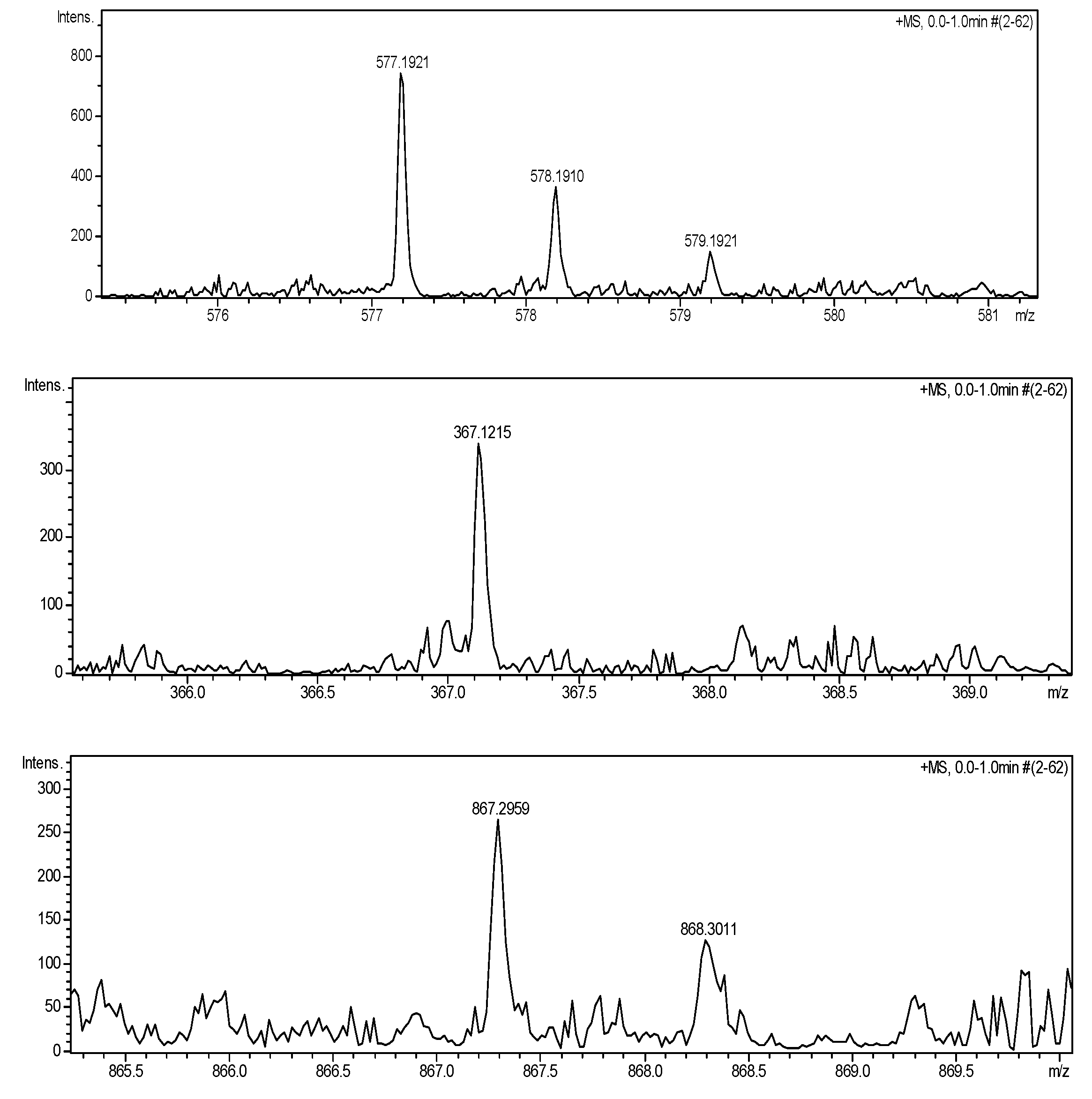
| BAs | Dns-BAs | [M + 23]+ (m/z) | [M + 23]+ (m/z) found |
|---|---|---|---|
| PUT | Dns2-PUT | 577.1914 | 577.1921 |
| HIS | Dns-HIS | 367.1199 | 367.1215 |
| SPD | Dns3-SPD | 867.3003 | 867.2959 |
| Biogenic Amines | Linear Range (μg/mL) | R2 | LOD (μg/mL) | LOQ (μg/mL) | Intra-Day Precision (%RSD) ( n = 5) | Inter-Day Precision (%RSD) ( n = 5) | Recovery (Young Leaves Samples) (% mean ± SD, n = 3) | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 μg/mL | 5 μg/mL | 10 μg/mL | |||||||
| MET | 0.10–10 | 0.9999 | 0.015 | 0.05 | 0.98 | 2.54 | 89.1 ± 4.9 | 91.7 ± 1.3 | 86.5 ± 1.1 |
| TRP | 0.10–10 | 0.9990 | 0.030 | 0.10 | 0.81 | 1.53 | 79.3 ± 4.6 | 92.1 ± 1.2 | 91.7 ± 1.0 |
| PEA | 0.25–10 | 0.9997 | 0.075 | 0.25 | 1.01 | 1.73 | 92.6 ± 5.3 | 94.3 ± 1.6 | 90.5 ± 1.2 |
| PUT | 0.10–50 | 0.9996 | 0.015 | 0.05 | 2.69 | 4.38 | 88.2 ± 5.4 | 90.6 ± 1.6 | 88.6 ± 1.2 |
| CAD | 0.10–10 | 0.9998 | 0.030 | 0.10 | 0.93 | 1.67 | 86.3 ± 4.3 | 93.6 ± 1.6 | 95.5 ± 1.1 |
| HIS | 0.10–10 | 0.9993 | 0.030 | 0.10 | 1.27 | 2.39 | 89.5 ± 4.7 | 91.2 ± 1.2 | 87.9 ± 1.1 |
| 5-HT | 0.25–10 | 0.9989 | 0.075 | 0.25 | 1.33 | 2.06 | 110.3 ± 5.1 | 88.1 ± 2.0 | 87.7 ± 1.7 |
| TYR | 0.25–10 | 0.9997 | 0.075 | 0.25 | 0.66 | 0.91 | 86.5 ± 4.1 | 88.2 ± 1.1 | 89.4 ± 1.3 |
| SPD | 0.10–10 | 0.9997 | 0.030 | 0.10 | 1.42 | 2.82 | 89.7 ± 5.7 | 92.6 ± 1.9 | 90.4 ± 1.4 |
| DA | 0.25–10 | 0.9989 | 0.075 | 0.25 | 1.09 | 2.43 | 93.1 ± 4.8 | 94.1 ± 2.5 | 88.5 ± 1.7 |
| SPM | 0.10–10 | 0.9996 | 0.030 | 0.10 | 2.07 | 3.31 | 96.5 ± 4.4 | 94.2 ± 1.4 | 90.1 ± 1.1 |
| Samples | Concentrations of BAs (mg/kg) (Mean ± SD) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MET | TRP | PEA | PUT | CAD | HIS | 5-HT | TYR | SPD | DA | SPM | |
| Young leaves | ND | ND | ND | 12.94 ± 1.8 ab | ND | 5.6 ± 0.9 c | ND | ND | 13.3 ± 1.6 a | ND | 23.7 ± 2.0 a |
| Mature leaves | ND | ND | ND | 2.3 ± 0.4 d | ND | 2.6 ± 0.3 c | ND | ND | 10.7 ± 1.5 abc | ND | 9.4 ± 1.1 bc |
| Young stem | ND | ND | ND | 10.4 ± 1.4 abc | ND | 10.2 ± 1.8 c | ND | ND | 4.3 ± 0.8 bc | ND | 13.3 ± 1.4 b |
| Mature stem | ND | ND | ND | 5.2 ± 1.1 acd | ND | 23.5 ± 2.2 b | ND | ND | 3.6 ± 0.6 bc | ND | 2.4 ± 0.4 d |
| Bark | ND | ND | ND | 3.0 ± 0.5 ad | ND | 102.7 ± 5.8 a | ND | ND | 3.0 ± 0.5 c | ND | 2.7 ± 0.4 d |
| Flower | ND | ND | ND | 20.9 ± 3.2 a | ND | 2.3 ± 0.4 c | ND | ND | 10.7 ± 2.1 abc | ND | 4.2 ± 0.7 d |
| Fruit | ND | ND | ND | 1.3 ± 0.3 d | ND | ND | ND | ND | 2.9 ± 0.1 bc | ND | 6.3 ± 1.2 cd |
| Root | ND | ND | ND | 5.2 ± 0.9 abcd | ND | 17.0 ± 3.2 bc | ND | ND | 3.0 ± 0.3 bc | ND | 0.9 ± 0.1 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ai, Y.; Sun, Y.N.; Liu, L.; Yao, F.Y.; Zhang, Y.; Guo, F.Y.; Zhao, W.J.; Liu, J.L.; Zhang, N. Determination of Biogenic Amines in Different Parts of Lycium barbarum L. by HPLC with Precolumn Dansylation. Molecules 2021, 26, 1046. https://doi.org/10.3390/molecules26041046
Ai Y, Sun YN, Liu L, Yao FY, Zhang Y, Guo FY, Zhao WJ, Liu JL, Zhang N. Determination of Biogenic Amines in Different Parts of Lycium barbarum L. by HPLC with Precolumn Dansylation. Molecules. 2021; 26(4):1046. https://doi.org/10.3390/molecules26041046
Chicago/Turabian StyleAi, Yun, Yan Ni Sun, Li Liu, Fang Yuan Yao, Yan Zhang, Feng Yi Guo, Wen Jie Zhao, Jian Li Liu, and Ning Zhang. 2021. "Determination of Biogenic Amines in Different Parts of Lycium barbarum L. by HPLC with Precolumn Dansylation" Molecules 26, no. 4: 1046. https://doi.org/10.3390/molecules26041046