
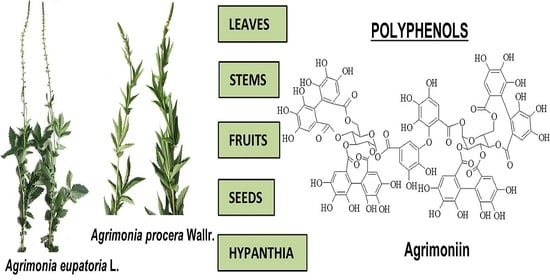
The Aerial Parts of Agrimonia procera Wallr. and Agrimonia eupatoria L. as a Source of Polyphenols, and Especially Agrimoniin and Flavonoids



Abstract
:
1. Introduction
2. Results and Discussion
2.1. Tentative Identification of Major Polyphenols in the Arial Parts of Common Agrimony and Fragrant Agrimony
2.2. The Content and Percentage Shares of Polyphenols in the Aerial Parts of Agrimonia Plants in the Main Stages of Plant Development
2.2.1. Agrimonia procera Wallr. Composition
2.2.2. Agrimonia eupatoria L. Composition
2.3. Differences in Polyphenol Content between the Studied Morphological Parts of Agrimonia Species
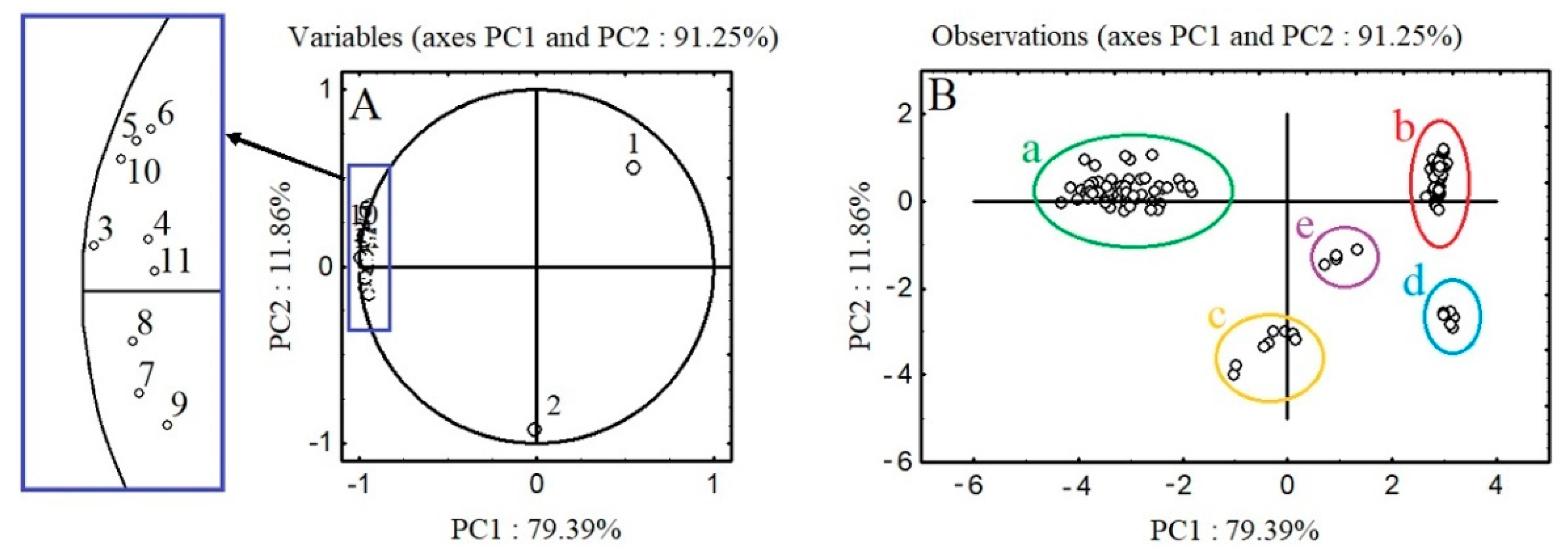
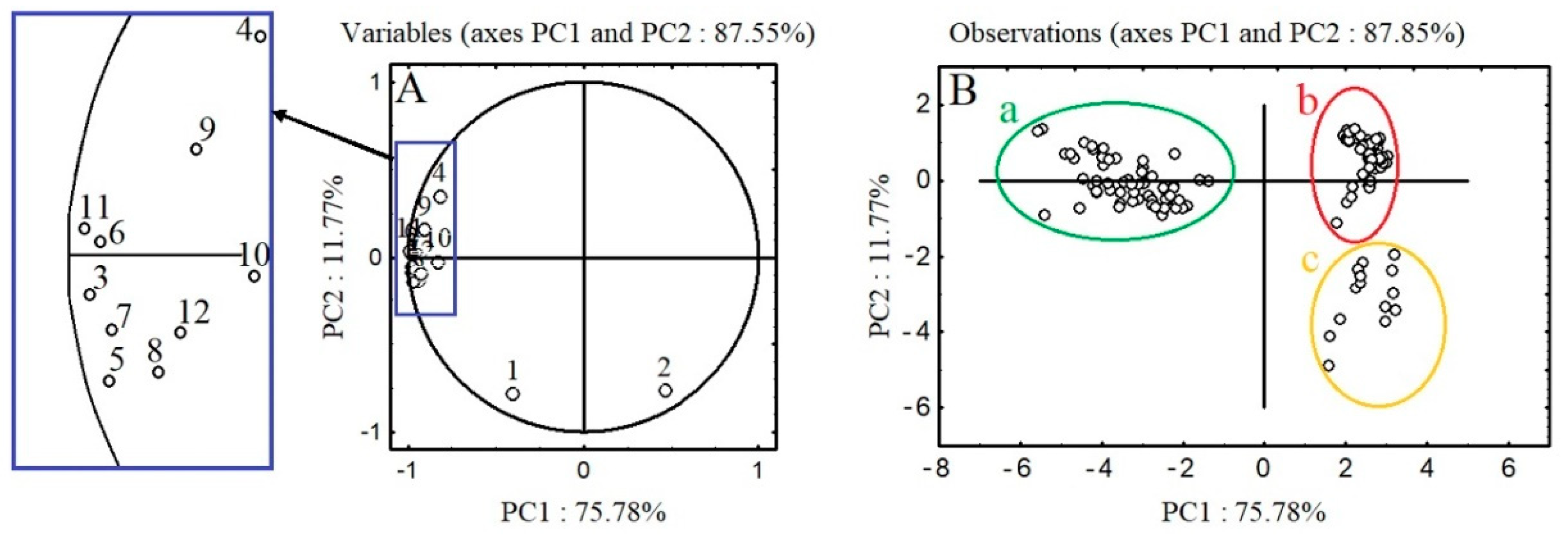
2.4. PCA Analysis of the Polyphenolic Profiles of A. procera and A. eupatoria
3. Materials and Methods
3.1. Plant Material
3.2. Course of the Experiment
3.2.1. Production of Seedlings
3.2.2. Cultivation, Fertilization and Tending Treatments during the Growing Season
3.2.3. Harvesting, Drying and Storage of the Raw Material
3.3. Polyphenol Extraction from the Stems and Seeds of A. procera and A. eupatoria
3.4. Polyphenol Extraction from the Leaves, Fruits, and Hypanthia of A. procera and A. eupatoria
3.5. Identification of Polyphenols
3.6. Quantitation of Polyphenols
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Alirezalu, A.; Salehi, P.; Ahmadi, N.; Sonboli, A.; Serena Aceto, S.; Hatami Maleki, H.; Ayyari, M. Flavonoids profile and antioxidant activity in flowers and leaves of hawthorn species (Crataegus spp.) from different regions of Iran. Int. J. Food Prop. 2018, 21, 452–470. [Google Scholar] [CrossRef] [Green Version]
- Gião, M.S.; Gomes, S.; Madureira, A.R.; Faria, A.; Pestana, D.; Calhau, C.; Pintado, M.E.; Azevedo, I.; Malcata, F.X. Effect of in vitro digestion upon the antioxidant capacity of aq. extracts of Agrimonia eupatoria, Rubus idaeus, Salvia sp. and Satureja montana. Food Chem. 2012, 131, 761–767. [Google Scholar] [CrossRef]
- Haggag, E.G.; Abdelhady, M.I.S.; Kamal, A.M. Phenolic content of Ruprechtia salicifolia leaf and its immunomodulatory, anti-inflammatory, anticancer and antibacterial activity. J. Pharm. Res. 2013, 6, 696–703. [Google Scholar] [CrossRef]
- Kosmala, M.; Zduńczyk, Z.; Juśkiewicz, J.; Jurgoński, A.; Karlińska, E.; Macierzyński, J.; Jańczak, R.; Rój, E. Chemical composition of defatted strawberry and raspberry seeds and the effect of these dietary ingredients on polyphenol metabolites, intestinal function, and selected serum parameters in rats. J. Agric. Food Chem. 2015, 63, 2989–2996. [Google Scholar] [CrossRef]
- Piwowarski, J.P.; Granica, S.; Stefańska, J.; Kiss, A.K. Differences in metabolism of ellagitannins by human gut microbiota ex vivo cultures. J. Nat. Prod. 2016, 79, 3022–3030. [Google Scholar] [CrossRef]
- Ivanova, D.; Vankova, D.; Nashar, M. Agrimonia eupatoria tea consumption in relation to markers of inflammation, oxidative status and lipid metabolism in healthy subjects. Arch. Physiol. Biochem. 2013, 119, 32–37. [Google Scholar] [CrossRef]
- Kuczmannová, A.; Gál, P.; Varinská, L.; Treml, J.; Kováč, I.; Novotný, M.; Vasilenko, T.; Dall’Acqua, S.; Nagy, M.; Mučaji, P. Agrimonia eupatoria L. and Cynara cardunculus L. water infusions: Phenolic profile and comparison of antioxidant activities. Molecules 2015, 20, 20538–20550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granica, S.; Kluge, H.; Horn, G.; Matkowski, A.; Kiss, A.K. The phytochemical investigation of Agrimonia eupatoria L. and Agrimonia procera Wallr. as valid sources of Agrimoniae herba—The pharmacopoeial plant material. J. Pharm. Biomed. Anal. 2015, 114, 272–279. [Google Scholar] [CrossRef]
- Rutkowski, L. Klucz do Oznaczania Roślin Naczyniowych Polski Niżowej; Wydawnictwo Naukowe PWN, SA.: Warsaw, Poland, 2018. [Google Scholar]
- Cybulska, H.; Janicka, H.; Karpała, Z.; Olesiński, A.; Rajkowski, Z.; Rumińska, A.; Tumiłowicz, H.; Wiszniewski, J. Uprawa i zbiór ziół, 3rd ed.; Państwowe Wydawnictwo Rolnicze i Leśne: Warsaw, Poland, 1956; pp. 495–496. [Google Scholar]
- Nowak, Z.T. Zioła z Polskich łąk; Wydawnictwo Aromat Słowa: Cracow, Poland, 2020; pp. 239–242. [Google Scholar]
- Correia, H.; González-Paramás, A.; Amaral, M.T.; Santos-Buelga, C.; Batista, M.T. Polyphenolic profile characterization of Agrimonia eupatoria L. by HPLC with different detection devices. Biomed. Chromat. 2006, 20, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Granica, S.; Krupa, K.; Kłębowska, A.; Kiss, A.K. Development and validation of HPLC-DAD-CAD-MS(3) method for qualitative and quantitative standardization of polyphenols in Agrimoniae eupatoriae herba (Ph. Eur). J. Pharm. Biomed. Anal. 2013, 86, 112–122. [Google Scholar] [CrossRef]
- Kubínová, R.; Jankovská, D.; Bauerová, V. Antioxidant and α-glucosidase inhibition activities and polyphenol content of five species of Agrimonia genus. Acta Fytotech. Zootech. 2012, 2, 38–41. [Google Scholar]
- Zhang, J.; Chen, Y. Studies on the lowering blood sugar substances from agrimony. Zhong Yao Cai 2009, 32, 1537–1539. [Google Scholar]
- Kool, M.M.; Comeskey, D.J.; Cooney, J.M.; McGhie, T.K. Structural identification of the main ellagitannins of a boysenberry (Rubus loganbaccus × baileyanus Britt.) extract by LC–ESI-MS/MS, MALDI-TOF-MS and NMR spectroscopy. Food Chem. 2010, 119, 1535–1543. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Kuwahara, M.; Memon, N.U.; Shingu, T. Tannins from Rosaceous medicinal plants. I. Structures of potentillin, agrimonic acid A and B, agrimoniin, a dimeric ellagitannin. Chem. Pharm. Bull. 1984, 32, 2165–2173. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Sarrias, A.; Gimez-Bastida, J.A.; Garcia-Conesa, M.T.; Gomez-Sanchez, M.B.; Garcia-Talavera, N.V.; Gil-Izquierdo, A.; Sanchez-Alvarez, C.; Fontana-Compiano, L.O.; Morga-Egea, J.P.; Pastor-Quirante, F.A.; et al. Occurrence of urolithins, gut microbiota ellagic acid metabolites and proliferation markers expression response in the human prostate gland upon consumption of walnuts and pomegranate juice. Mol. Nutr. Food Res. 2010, 54, 311–322. [Google Scholar] [CrossRef]
- Seeram, N.; Aronson, W.J.; Zhang, Y.; Henning, S.; Moro, A.; Lee, R.P.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegranate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef]
- Bazylko, A.; Piwowarski, J.P.; Filipek, A.; Bonarewicz, J.; Tomczyk, M. In vitro antioxidant and anti-inflammatory activities of extracts from Potentilla recta and its main ellagitannin, agrimoniin. J. Ethnopharmacol. 2013, 149, 222–227. [Google Scholar] [CrossRef]
- Funatogawa, K.; Hayashi, S.; Shimomura, H.; Yoshida, T.; Hatano, T.; Ito, H.; Hirai, Y. Antibacterial activity of hydrolyzable tannins derived from medicinal plants against Helicobacter pylori. Microbiol. Immunol. 2004, 48, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Murayama, T.; Nomura, M.; Hatano, T.; Yoshida, T.; Furukawa, T.; Koshiura, R.; Okuda, T. Antitumor activity and Interleukin-1 induction by tannins. Anticancer Res. 1993, 13, 37–42. [Google Scholar]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Le Cornu, K.A.; Ryder, J.J.; Hall, W.L.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Rice-Evans, C.A. Luteolin-rich artichoke extract protects low density lipoprotein from oxidation in vitro. Free Radic. Res. 1998, 29, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Yamazaki, C.; Yamazaki, M. Inhibitory effect of Perilla leaf extract and luteolin on mouse skin tumor promotion. Biol. Pharm. Bull. 2003, 26, 560–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Q.; Zhao, Y.; Chen, X.; Zhao, K.; Zhai, Q.; Tu, F. Apigenin protects the brain against ischemia/reperfusion injury via caveolin-1/VEGF in vitro and in vivo. Oxid. Med. Cell. Longev. 2018, 2018, 7017204. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Xu, Y.Q.; Wu, J.C.; Hang, P.Z.; Wang, Y.; Wang, C.; Wu, J.W.; Qi, J.C.; Zhang, Y.; Du, Z.M. Vitexin protects against cardiac hypertrophy via inhibiting calcineurin and CaMKII signaling pathways. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 386, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Jing, Z.; Wang, Z.; Li, X.; Li, X.; Cao, T.; Bi, Y.; Zhou, J.; Chen, X.; Yu, D.; Zhu, L.; et al. Protective effect of quercetin on posttraumatic cardiac injury. Nature 2016, 6, 30812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhanpal, P.; Rai, D.K. Quercetin: A versatile flavonoid. Internet J. Medical Update 2007, 2, 22–37. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Pu, X.P. Neuroprotective effect of kaempferol against a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced mouse model of Parkinson’s disease. Biol. Pharm. Bull. 2011, 34, 1291–1296. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Schieber, A.; Bererdini, N.; Carle, R. Identification of flavonol and xanthone glycosides from mango (Mangifera indica L. Cv. “Tommy Atkins”) peels by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2003, 51, 5006–5011. [Google Scholar] [CrossRef]
- Karlińska, E.; Pecio, Ł.; Macierzyński, J.; Stochmal, A.; Kosmala, M. Structural elucidation of the ellagitannin with a molecular weight of 2038 isolated from strawberry fruit (Fragaria ananassa Duch.) and named fragariin A. Food Chem. 2019, 296, 109–115. [Google Scholar] [CrossRef]
- Lee, K.Y.; Hwang, L.; Jeong, E.J.; Kim, S.H.; Kim, Y.C.; Sung, S.H. Effect of neuroprotective flavonoids of Agrimonia eupatoria on glutamate-induced oxidative injury to HT22 hippocampal cells. Biosci. Biotechnol. Biochem. 2010, 74, 1704–1706. [Google Scholar] [CrossRef] [Green Version]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium, Julius Kühn-Institut: Quedlinburg, Germany, 2018; pp. 14–17. [Google Scholar] [CrossRef]
- Gräber, T.; Kluge, H.; Granica, S.; Horn, G.; Brandsch, C.; Stangl, G.I. Studies on the health impact of Agrimonia procera in piglets. BMC Vet. Res. 2014, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Fernández, M.; Ludwig, I.A.; De Peña, M.-P.; Cid, C. Bioaccessibility of Tudela artichoke (Cynara scolymus cv. Blanca de Tudela) (poly)phenols: The effects of heat treatment, simulated gastrointestinal digestion and human colonic microbiota. Food Funct. 2021, 12, 1996–2011. [Google Scholar] [CrossRef]
- Kim, D.K.; Son, D.M.; Chon, S.U.; Lee, K.D.; Rim, Y.S. Variation of vitexin and isovitexin contents in mungbean (Vigna radiata (L.) Wilczek) germplasms. Korean J. Plant. Resour. 2009, 22, 128–135. [Google Scholar]
- Sagaradze, V.A.; Babaeva, E.Y.; Ufimov, R.A.; Trusov, N.A.; Kalenikova, E.I. Study of the variability of rutin, vitexin, hyperoside, quercetin in “Crataegi folium cum flore” of hawthorn (Crataegus L.) species from Russian flora. J. Appl. Res. Med. Aroma. Plants 2019, 15, 100217. [Google Scholar] [CrossRef]
- Rusalepp, L.; Raal, A.; Püssa, T.; Maeorg, U. Comparison of chemical composition of Hypericum perforatum and H. maculatum in Estonia. Biochem. Syst. Ecol. 2017, 73, 41–46. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Guella, G.; Palmieri, L.; Martinatti, P.; Pojer, E.; Mattivi, F.; Vrhovsek, U. Evolution of ellagitannin content and profle during fruit ripening in Fragaria spp. J. Agric. Food Chem. 2013, 61, 8597–8607. [Google Scholar] [CrossRef] [Green Version]
- Karlińska, E.; Masny, A.; Cieślak, M.; Macierzyński, J.; Pecio, Ł.; Stochmal, A.; Kosmala, M. Ellagitannins in roots, leaves, and fruits of strawberry (Fragaria × ananassa Duch.) vary with developmental stage and cultivar. Sci. Hortic. 2021, 275, e109665. [Google Scholar] [CrossRef]
- Wittmann, W.; Strobl, W. Untersuchungen am Artenpaar Agrimonia eupatoria L. A. procera Wallr. Im Bundesland Salzburg (Österreich). Lizner Biol. Beitr. 1987, 19, 91–119. [Google Scholar]
- Saadati, N.; Abdullah, M.P.; Zakaria, Z.; Sany, S.B.T.; Rezayi, M.; Hassonizadeh, H. Limit of detection and limit of quantification development procedures for organochlorine pesticides analysis in water and sediment matrices. Chem. Cent. J. 2013, 63, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Peak No. | Compound | RT [min] | UV [nm] | MS Data [m/z] | MS/MS Data | Occurrence | Identification | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. procera Wallr. | A. eupatoria L. | ||||||||||
| L S F H | Se | L S | F H | Se | |||||||
| 1 | Quercetin arabinoglycoside a | 35.5 | 257, 355 | [595.14] −1 | 463, 445, 301 * | + | + | − | − | − | [3] |
| 2 | Agrimoniin | 36.3 | 260sh | [934.08] −2 | 1567, 1235, 1085, 935, 897, 783, 633, 301 | + | + | + | + | + | Standard, [8,13,33] |
| 3 | Apigenin 8-C-glucoside | 37.9 | 270, 353 | [431.07] −1 | 341, 311 | − | − | + | + | − | Standard, [8] |
| 4 | Quercetin 3-O-rhamnoglucoside | 38.3 | 257, 354 | [609.12] −1 | 463, 343, 301 | + | − | + | + | − | Standard, [8,13,14,15,34] |
| 5 | Apigenin 6-C-glucoside | 38.7 | 268, 350 | [431.08] −1 | 341, 311 | − | − | + | + | − | Standard, [8,12,13,14] |
| 6 | Ellagic acid | 38.8 | 254, 350 | [301.10] −1 | − | + | + | + | + | + | Standard, [8] |
| 7 | Quercetin 3-O-galactoside | 39.4 | 257, 353 | [463.08] −1 | 343, 301 | + | − | + | + | − | Standard, [12,13,14] |
| 8 | Kaempferol 3-O-glucoside | 40.0 | 267, 350 | [447.09] −1 | 327, 285, 269, 255, 151 | + | − | − | − | − | Standard, [12,13,15,34] |
| 9 | Luteolin 7-O-glucuronide a | 40.6 | 266, 349 | [461.07] −1 | 357, 327, 285, 175, 151, 113 | + | − | + | + | − | [7,8,13,34] |
| 10 | Luteolin 7-O-glucoside a | 42.5 | 267, 337 | [447.12] −1 | 327, 285, 175, 151, 113 | + | − | + | − | − | [7,8,34] |
| 11 | Apigenin 7-O-glucuronide a | 43.1 | 268, 339 | [445.07] −1 | 269, 175, 113 | + | − | + | + | + | [7,8,13,34] |
| 12 | Apigenin 7-O-glucoside a | 43.5 | 269, 343 | [431.08] −1 | 311, 327, 269, 175 | + | − | + | + | − | Standard, [8,13,15] |
| 13 | Quercetin 3-O-rhamnoside | 44.3 | 261, 350 | [447.09] −1 | 343, 301, 179 | − | − | + | + | − | Standard, [7,8,13,34] |
| 14 | KpCG * | 46.2 | 269, 315 | [593.12] −1 | 447, 307, 285 | + | + | + | + | + | Standard, [12,34] |
| 15 | KpCG * isomer | 46.7 | 269, 315 | [593.12] −1 | 447, 307, 285 | + | + | + | + | + | [12,34] |
| Agrimoniin | Ellagic Acid | Quercetin Arabinoglycoside | Quercetin 3-O-rhamnoglucoside | Quercetin 3-O-galactoside | Kaempferol 3-O-glucoside | Sum of KpCG * Isomer | Luteolin 7-O-glucuronide | Luteolin 7-O-glucoside | Apigenin 7-O-glucuronide | Apigenin 7-O-glucoside | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stage of Development | ||||||||||||
| LEAVES | ||||||||||||
| S1 | 2166.0 ± 402.0c (49.8) | 39.4 ± 7.1a (0.9) | 611.0 ± 27.5b (14.1) | 88.5 ± 5.3c (2.0) | 96.2 ± 16.3b (2.2) | 84.2 ± 4.2b (1.9) | 58.3 ± 5.3c (1.3) | 181.2 ± 7.7c (4.0) | 29.0 ± 4.1a (0.6) | 1015.1 ± 8.5c (22.6) | 113.4 ± 8.5b (2.5) | 4482.2 ± 356.8c |
| S2 | 2002.8 ± 306.7bc (45.9) | 49.7 ± 9.1c (1.1) | 653.4 ± 44.9c (15.0) | 83.2 ± 6.2c (1.9) | 105.6 ± 13.0bc (2.4) | 81.3 ± 4.7b (1.9) | 53.5 ± 6.6bc (1.2) | 176.3 ± 14.7c (4.0) | 32.5 ± 2.3a (0.7) | 1007.4 ± 90.1c (23.1) | 117.3 ± 8.5b (2.7) | 4362.7 ± 395.4c |
| S3 | 1872.1 ± 128.8b (49.1) | 38.2 ± 2.9a (1.0) | 551.5 ± 44.9a (14.5) | 66.0 ± 4.3b (1.7) | 81.9 ± 16.4a (2.1) | 65.9 ± 4.5a (1.7) | 59.6 ± 7.0c (1.6) | 141.0 ± 15.4ab (3.7) | 29.0 ± 5.5a (0.8) | 808.4 ± 104.7a (21.2) | 98.5 ± 15.9a (2.6) | 3812.2 ± 143.0b |
| S4 | 1583.6 ± 176.1a (45.8) | 41.4 ± 5.5ab (1.2) | 569.4 ± 67.7ab (16.5) | 66.4 ± 8.3b (1.9) | 101.8 ± 17.5bc (2.9) | 68.0 ± 9.5a (2.0) | 49.7 ± 11.4b (1.4) | 152.9 ± 18.5b (4.4) | 37.4 ± 5.4b (1.1) | 911.0 ± 101.7b (26.4) | 119.1 ± 13.8b (3.4) | 3455.6 ± 510.3a |
| S5 | 1595.6 ± 166.0a (44.1) | 47.0 ± 7.0bc (1.3) | 530.2 ± 27.2a (14.6) | 58.2 ± 5.6a (1.6) | 112.9 ± 12.3c (3.1) | 62.6 ± 5.5a (1.7) | 42.8 ± 4.2a (1.2) | 138.8 ± 8.4a (3.8) | 43.1 ± 3.5c (1.2) | 869.3 ± 43.4ab (24.0) | 120.5 ± 4.7b (3.3) | 3621.0 ± 134.7ab |
| Mean | 1855.6 ± 342.8 (46.8) | 43.9 ± 8.3 (1.1) | 594.7 ± 65.1 (15.0) | 74.2 ± 12.6 (1.9) | 100.8 ± 17.2 (2.5) | 73.9 ± 10.3 (1.9) | 52.8 ± 9.2 (1.3) | 161.5 ± 21.8 (4.1) | 34.1 ± 6.3 (0.9) | 939.0 ± 112.4 (23.6) | 114.9 ± 12.6 (2.9) | 3963.1 ± 539.1 |
| STEMS | ||||||||||||
| S1 | 2641.5 ± 457.6a (95.0) | 35.1 ± 5.8ab (1.3) | 61.5 ± 8.9a (2.2) | 9.3 ± 1.0a (0.3) | 11.0 ± 2.0a (0.4) | 3.1 ± 0.5a (0.1) | 8.0 ± 1.4c (0.3) | 3.4 ± 0.5a (0.1) | 1.9 ± 0.3a (0.1) | 4.7 ± 1.2a (0.2) | 1.5 ± 0.2a (0.1) | 2781.1 ± 466.5a |
| S2 | 2703.7 ± 588.4a (93.7) | 41.0 ± 10.8c (1.4) | 86.1 ± 18.4b (3.0) | 10.4 ± 1.8ab (0.4) | 14.4 ± 3.3b (0.5) | 4.3 ± 1.1b (0.1) | 6.8 ± 0.9b (0.8) | 5.2 ± 1.5b (0.2) | 2.7 ± 0.5b (0.1) | 9.5 ± 3.0b (0.3) | 2.8 ± 0.9b (0.1) | 2886.8 ± 621.8a |
| S3 | 3178.4 ± 84.3b (94.2) | 31.7 ± 1.1a (0.9) | 103.7 ± 7.3c (3.1) | 10.8 ± 0.8b (0.3) | 14.1 ± 1.2b (0.4) | 4.7 ± 0.6b (0.1) | 5.8 ± 0.3a (0.2) | 6.1 ± 0.7bc (0.2) | 3.1 ± 0.4bc (0.1) | 12.0 ± 2.4bc (0.4) | 3.5 ± 0.5b (0.1) | 3373.9 ± 87.3b |
| S4 | 3166.4 ± 493.7b (93.6) | 31.7 ± 5.9a (0.9) | 114.9 ± 27.5c (3.4) | 11.8 ± 2.3b (0.4) | 16.4 ± 3.8bc (0.5) | 5.1 ± 1.4bc (0.2) | 5.5 ± 1.0a (0.2) | 7.2 ± 2.0cd (0.2) | 3.6 ± 1.2c (0.1) | 16.9 ± 5.1d (0.5) | 4.3 ± 1.2bc (0.1) | 3383.8 ± 540.0b |
| S5 | 3926.7 ± 420.7c (94.0) | 36.6 ± 2.6ab (0.9) | 141.8 ± 10.5d (3.4) | 13.3 ± 0.8c (0.3) | 17.6 ± 1.7c (0.4) | 5.8 ± 0.5c (0.1) | 5.1 ± 0.3a (0.1) | 7.7 ± 0.4d (0.2) | 4.3 ± 0.5d (0.1) | 14.6 ± 1.2cd (0.3) | 4.5 ± 0.5c (0.1) | 4178.1 ± 437.6c |
| Mean | 3038.8 ± 628.5 (94.0) | 35.9 ± 8.0 (1.1) | 98.9 ± 30.4 (3.1) | 11.0 ± 2.0 (0.3) | 14.7 ± 3.5 (0.5) | 4.5 ± 1.3 (0.1) | 6.3 ± 1.3 (0.4) | 5.9 ± 1.9 (0.2) | 3.1 ± 1.0 (0.1) | 11.5 ± 5.2 (0.4) | 3.1 ± 1.3 (0.1) | 3233.8 ± 669.4 |
| FRUITS | ||||||||||||
| S3 | 2082.1 ± 51.9b (59.1) | 161.8 ± 6.3b (4.6) | 672.5 ± 0.4b (19.1) | 74.8 ± 0.9c (2.1) | 141.2 ± 1.0c (4.0) | 25.5 ± 0.1c (0.7) | 38.4 ± 0.6b (1.1) | 83.9 ± 0.1b (2.4) | 15.5 ± 1.4a (0.4) | 108.1 ± 2.0b (3.1) | 12.5 ± 1.2b (0.4) | 3526.0 ± 71.8b |
| S4 | 1533.4 ± 83.8a (56.2) | 127.3 ± 7.3a (4.7) | 569.8 ± 43.6a ()20.9 | 64.2 ± 4.2b (2.4) | 100.1 ± 7.0b (3.7) | 19.1 ± 1.1b (0.7) | 23.9 ± 1.8a (0.9) | 74.1 ± 5.9b (2.7) | 13.4 ± 1.0a (0.5) | 102.4 ± 8.0b (3.8) | 6.3 ± 0.3a (0.2) | 2728.9 ± 170.8a |
| S5 | 1414.1 ± 17.0a ()57.3 | 127.4 ± 2.8a (5.2) | 498.0 ± 1.0a (20.2) | 51.4 ± 1.6a (2.1) | 77.9 ± 0.8a (3.2) | 14.5 ± 0.3a (0.6) | 35.0 ± 0.1b (1.4) | 61.7 ± 0.3a (2.5) | 12.9 ± 0.1a (0.5) | 83.9 ± 1.3a (3.4) | 5.3 ± 0.5a (0.2) | 2469.4 ± 19.0a |
| Mean | 1676.5 ± 321.8 (57.7) | 138.8 ± 18.3 (4.8) | 580.1 ± 80.9 (19.9) | 63.5 ± 10.6 (2.2) | 106.4 ± 28.9 (3.7) | 19.7 ± 5.0 (0.7) | 32.4 ± 6.9 (1.1) | 73.2 ± 10.3 (2.5) | 13.9 ± 1.5 (0.5) | 98.1 ± 11.9 (3.4) | 8.0 ± 3.5 (0.3) | 2908.1 ± 499.5 |
| SEEDS | ||||||||||||
| S3 | 2036.3 ± 71.7c (93.3) | 139.8 ± 5.2b (6.4) | 2.0 ± 0.2a (0.1) | nd | nd | nd | 4.8 ± 0.3b (0.2) | nd | nd | nd | nd | 2183.9 ± 67.4c |
| S4 | 1754.9 ± 59.4b (92.8) | 129.7 ± 8.7ab (6.9) | 2.3 ± 0.1a (0.1) | nd | nd | nd | 4.4 ± 0.5b (0.2) | nd | nd | nd | nd | 1891.3 ± 68.5b |
| S5 | 1189.0 ± 25.2a (87.9) | 115.1 ± 0.9a (8.5) | 8.3 ± 0.5b (0.6) | nd | nd | nd | 1.9 ± 0.1a (0.3) | nd | nd | nd | nd | 1353.9 ± 24.6a |
| Mean | 1660.1 ± 388.4 (91.8) | 128.2 ± 12.0 (7.1) | 4.2 ± 3.2 (0.2) | nd | nd | nd | 4.6 ± 0.4 (0.3) | nd | nd | nd | nd | 1809.1 ± 379.2 |
| HYPANTHIA | ||||||||||||
| S3 | 236.2 ± 13.3a (26.8) | 37.5 ± 1.9a (4.3) | 383.2 ± 2.9a (43.4) | 38.9 ± 0.2a (4.4) | 59.5 ± 0.3ab (6.7) | 9.7 ± 0.3a (1.1) | 8.8 ± 0.3a (1.0) | 44.2 ± 0.2a (5.0) | 7.2 ± 0.4a (0.9) | 52.1 ± 1.6a (5.9) | 4.7 ± 1.3a (0.5) | 882.2 ± 17.5a |
| S4 | 255.3 ± 57.0a (29.2) | 38.8 ± 7.5a (4.4) | 359.1 ± 94.3a (41.1) | 39.7 ± 10.3a (4.5) | 53.1 ± 12.8a (6.1) | 8.7 ± 1.8a (1.0) | 8.4 ± 2.3a (1.0) | 45.8 ± 10.4a (5.2) | 7.2 ± 1.8a (0.8) | 54.2 ± 13.8a (6.2) | 3.6 ± 0.3a (0.4) | 873.8 ± 211.7a |
| S5 | 525.6 ± 69.1b (33.5) | 110.6 ± 10.2b (7.0) | 572.4 ± 38.6b (36.4) | 60.8 ± 1.3b (3.9) | 78.6 ± 1.2b (5.0) | 13.5 ± 0.4b (0.9) | 32.6 ± 2.7b (2.1) | 72.3 ± 2.0b (4.6) | 17.4 ± 0.6b (1.1) | 78.1 ± 4.2a (5.0) | 9.1 ± 0.2b (0.6) | 1571.1 ± 130.2b |
| Mean | 339.0 ± 150.3 (30.6) | 62.3 ± 37.8 (5.6) | 438.2 ± 114.0 (39.5) | 46.5 ± 12.1 (4.2) | 63.7 ± 13.2 (5.7) | 10.7 ± 2.4 (1.0) | 16.6 ± 12.5 (1.5) | 54.1 ± 14.9 (4.9) | 10.6 ± 5.3 (1.0) | 61.5 ± 14.5 (5.5) | 5.8 ± 2.7 (0.5) | 1109.1 ± 374.9 |
| Agrimoniin | Ellagic Acid | Quercetin 3-O-rhamnoglucoside | Quercetin 3-O-galactoside | Quercetin 3-O-rhamnoside | Sum of KpCG*Isomer | Luteolin 7-O-glucuronide | Luteolin 7-O-glucoside | Apigenin 7-O-glucuronide | Apigenin 7-O-glucoside | Apigenin 8-C-glucoside | Apigenin 6-C-glucoside | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stage of Development | |||||||||||||
| LEAVES | |||||||||||||
| S1 | 417.3 ± 45.5a (30.0) | 12.9 ± 3.4d (0.9) | 23.0 ± 4.5a (1.7) | 85.1 ± 18.5a (10.7) | 102.9 ± 17.2b (7.4) | 47.5 ± 8.7c (3.4) | 145.4 ± 23.8b (10.5) | 11.7 ± 2.2a (0.8) | 222.8 ± 25.8b (16.0) | 55.8 ± 7.6ab (4.0) | 53.0 ± 6.5b (3.8) | 149.9 ± 14.9ab (10.8) | 1327.4 ± 109.6a |
| S2 | 419.9 ± 71.4a (31.5) | 9.6 ± 3.5c (0.7) | 28.6 ± 6.4ab (2.2) | 98.6 ± 22.4a (7.4) | 103.1 ± 13.5b (7.7) | 51.4 ± 9.6c (3.9) | 130.2 ± 18.3b (9.8) | 13.8 ± 2.8ab (1.0) | 221.0 ± 18.8b (16.6) | 56.3 ± 6.5ab (4.2) | 53.8 ± 4.1b (4.0) | 144.7 ± 15.3ab (10.9) | 1331.0 ± 80.1a |
| S3 | 491.2 ± 65.4c (39.0) | 6.1 ± 2.1ab (0.5) | 22.6 ± 3.3a (1.8) | 81.5 ± 7.3a (6.5) | 87.1 ± 12.6a (6.9) | 35.2 ± 4.1ab (2.8) | 98.8 ± 14.6a (7.8) | 12.6 ± 1.9a (1.0) | 185.5 ± 22.1a (14.7) | 52.7 ± 6.4a (4.2) | 49.9 ± 1.7b (4.0) | 137.3 ± 11.6a (10.9) | 1260.7 ± 60.9a |
| S4 | 420.0 ± 70.9a (33.2) | 7.5 ± 2.8bc (0.6) | 34.0 ± 16.3b (2.7) | 98.9 ± 39.0a (7.8) | 90.7 ± 18.2ab (7.2) | 32.5 ± 5.3a (2.6) | 104.3 ± 18.6a (8.2) | 18.3 ± 10.1b (1.4) | 188.7 ± 33.5a (14.9) | 64.2 ± 16.9bc (5.1) | 41.2 ± 7.7a (3.3) | 165.2 ± 47.6b (13.0) | 1265.6 ± 146.3a |
| S5 | 401.0 ± 43.1a (30.1) | 4.6 ± 0.6a (0.3) | 47.1 ± 5.6c (3.5) | 126.8 ± 18.9b (9.5) | 103.1 ± 6.9a (7.7) | 40.2 ± 4.6b (3.0) | 112.8 ± 6.0a (8.5) | 25.7 ± 4.5c (1.9) | 202.8 ± 13.6ab (15.2) | 70.4 ± 9.1c (5.3) | 50.8 ± 4.6b (3.8) | 146.7 ± 25.5ab (11.0) | 1332.0 ± 91.7a |
| Mean | 426.7 ± 666.8 (32.4) | 8.5 ± 3.9 (0.6) | 30.7 ± 11.9 (2.3) | 98.3 ± 24.0 (8.3) | 97.9 ± 15.7 (7.4) | 42.4 ± 10.6 (3.2) | 119.9 ± 24.0 (9.1) | 16.0 ± 7.1 (1.2) | 206.2 ± 28.3 (15.7) | 59.6 ± 11.7 (4.5) | 49.7 ± 7.3 (3.8) | 149.8 ± 28.4 (11.4) | 1305.9 ± 130.6 |
| STEMS | |||||||||||||
| S1 | 175.5 ± 31.8a (74.5) | 16.4 ± 5.3cd (6.9) | 2.9 ± 1.1a (1.2) | 5.3 ± 1.52a (2.2) | 5.1 ± 1.2a (2.2) | 10.6 ± 2.2c (4.5) | 3.2 ± 0.8a (1.4) | 0.9 ± 0.3a (0.4) | 2.1 ± 0.5a (0.9) | 1.1 ± 0.2a (0.5) | 0.7 ± 0.2a (0.3) | 11.7 ± 3.6bc (5.0) | 235.6 ± 35.9a |
| S2 | 161.4 ± 24.9a (68.1) | 17.8 ± 3.7d (7.5) | 5.9 ± 1.7b (2.5) | 10.0 ± 3.0ab (4.2) | 7.1 ± 2.2ab (3.0) | 9.3 ± 2.2bc (3.9) | 5.0 ± 1.3b (2.1) | 1.4 ± 0.3b (0.6) | 3.0 ± 0.5b (1.3) | 1.7 ± 0.6b (0.7) | 1.2 ± 0.2b (0.5) | 13.2 ± 2.4c (5.6) | 237.1 ± 29.5a |
| S3 | 246.2 ± 37.3b (76.6) | 14.3 ± 1.4c (4.5) | 5.9 ± 2.2b (1.8) | 11.6 ± 4.3b (3.6) | 8.6 ± 2.5b (2.7) | 8.7 ± 1.2ab (2.7) | 5.6 ± 2.0bc (1.8) | 1.4 ± 0.3b (0.4) | 3.9 ± 0.4c (1.2) | 1.8 ± 0.3b (0.6) | 1.7 ± 0.2c (0.5) | 11.7 ± 1.1bc (3.6) | 321.4 ± 46.1b |
| S4 | 330.4 ± 101.9c (80.9) | 10.6 ± 2.9b (2.6) | 7.7 ± 3.3b (1.9) | 17.7 ± 9.5c (4.3) | 8.3 ± 2.9b (2.0) | 7.1 ± 1.8a (1.7) | 6.9 ± 3.1cd (1.7) | 1.6 ± 0.6bc (0.4) | 4.1 ± 0.5c (1.0) | 2.0 ± 0.9b (0.5) | 2.6 ± 0.5e (0.6) | 9.7 ± 2.1ab (2.4) | 408.7 ± 102.8c |
| S5 | 236.9 ± 35.1b (72.5) | 6.7 ± 1.9a (2.0) | 12.3 ± 4.4c (3.8) | 22.7 ± 6.4c (7.0) | 11.4 ± 2.0c (3.5) | 8.8 ± 1.6ab (2.7) | 8.6 ± 2.2d (2.6) | 1.8 ± 0.6c (0.6) | 4.2 ± 0.3c (1.3) | 3.1 ± 0.7c (0.9) | 2.2 ± 0.1d (0.7) | 8.0 ± 4.1a (2.4) | 326.7 ± 39.9b |
| Mean | 226.8 ± 91.2 (75.0) | 13.8 ± 5.2 (4.6) | 6.7 ± 3.7 (2.2) | 13.0 ± 7.9 (4.3) | 7.8 ± 2.9 (3.0) | 8.9 ± 2.2 (2.9) | 5.7 ± 2.6 (1.9) | 1.4 ± 0.5 (0.5) | 3.4 ± 0.9 (1.1) | 1.9 ± 0.8 (0.6) | 1.7 ± 0.8 (0.6) | 11.2 ± 3.3 (3.7) | 302.3 ± 95.2 |
| FRUITS | |||||||||||||
| S3 | 1004.7 ± 80.3c (77.4) | 49.9 ± 5.4a (3.8) | 6.7 ± 0.3c (0.5) | 28.6 ± 0.1c (2.2) | 7.9 ± 0.6c (0.6) | 20.3 ± 0.1b (1.6) | 4.2 ± 0.4b (0.3) | nd | 77.3 ± 3.0b (5.9) | 6.0 ± 0.1c (0.5) | 8.3 ± 0.1b (0.6) | 63.7 ± 0.9c (4.9) | 1297.7 ± 84.8b |
| S4 | 639.6 ± 10.9b (75.8) | 46.3 ± 0.4a (5.5) | 4.3 ± 0.2a (0.5) | 17.6 ± 0.1a (2.1) | 4.7 ± 0.2a (0.6) | 13.8 ± 3.2a (1.6) | 2.9 ± 0.1a (0.3) | nd | 59.4 ± 1.6a (7.0) | 2.9 ± 0.1a (0.3) | 7.4 ± 0.1a (0.9) | 39.4 ± 0.5a (4.7) | 843.5 ± 17.2a |
| S5 | 470.3 ± 52.2a (67.3) | 49.6 ± 1.7a (7.5) | 5.1 ± 0.1b (0.7) | 25.5 ± 0.5b (3.7) | 6.7 ± 0.2b (1.0) | 13.6 ± 0.1a (2.0) | 3.2 ± 0.0a (0.5) | nd | 62.6 ± 2.2a (9.0) | 3.7 ± 0.2b (0.5) | 7.6 ± 0.1a (1.1) | 47.7 ± 0.5b (6.8) | 698.4 ± 54.5a |
| Mean | 704.8 ± 248.0 (74.5) | 48.6 ± 3.1 (5.1) | 5.4 ± 1.1 (0.6) | 23.9 ± 5.1 (2.5) | 6.4 ± 1.5 (0.7) | 15.9 ± 3.7 (1.7) | 3.4 ± 0.6 (0.4) | nd | 66.3 ± 8.6 (7.0) | 4.2 ± 1.4 (0.4) | 7.7 ± 0.4 (0.8) | 50.3 ± 11.1 (5.3) | 946.5 ± 283.3 |
| SEEDS | |||||||||||||
| S3 | 720.5 ± 37.0c (92.6) | 52.3 ± 2.8ab (6.7) | nd | nd | nd | 2.6 ± 0.3a (0.3) | nd | nd | 3.1 ± 0.1b (0.4) | nd | nd | nd | 778.5 ± 40.4c |
| S4 | 549.9 ± 21.1b (89.8) | 59.9 ± 5.7b (9.8) | nd | nd | nd | 2.4 ± 0.6a (0.4) | nd | nd | 2.4 ± 0.2a (0.4) | nd | nd | nd | 614.7 ± 27.6b |
| S5 | 307.3 ± 68.7a (85.8) | 46.9 ± 0.5a (13.1) | nd | nd | nd | 1.6 ± 0.4a (0.5) | nd | nd | 2.4 ± 0.3a (0.7) | nd | nd | nd | 358.2 ± 68.5a |
| Mean | 529.9 ± 189.2 (90.1) | 53.0 ± 6.5 (9.1) | nd | nd | nd | 2.2 ± 0.6 (0.4) | nd | nd | 2.6 ± 0.4 (0.4) | nd | nd | nd | 583.8 ± 193.2 |
| HYPANTHIA | |||||||||||||
| S3 | 164.4 ± 18.8a (47.8) | 21.7 ± 0.9a (6.3) | 4.5 ± 0.4a (1.3) | 16.6 ± 2.8a (4.8) | 4.0 ± 0.4a (1.2) | 8.5 ± 0.6a (2.5) | 2.9 ± 0.1b (0.9) | nd | 70.8 ± 9.3a (20.6) | 3.1 ± 0.3a (0.9) | 6.8 ± 0.8a (2.0) | 40.4 ± 6.2a (11.8) | 343.9 ± 40.4a |
| S4 | 285.7 ± 52.4ab (50.3) | 33.1 ± 5.4a (5.8) | 10.9 ± 2.1b (1.9) | 32.6 ± 6.3a (5.7) | 8.1 ± 1.2a (1.4) | 9.7 ± 1.5ab (1.7) | 5.8 ± 1.1c (1.0) | nd | 100.5 ± 16.5a (17.7) | 8.7 ± 1.3b (1.5) | 9.9 ± 1.8a (1.7) | 63.2 ± 12.4a (11.1) | 568.3 ± 102.1ab |
| S5 | 383.2 ± 55.7b (52.6) | 76.0 ± 12.0b (10.4) | 11.7 ± 3.1b (1.6) | 41.2 ± 12.9a (5.7) | 18.1 ± 4.3b (2.5) | 16.6 ± 3.4b (2.3) | 2.1 ± 0.1a (0.3) | nd | 92.9 ± 25.1a (12.7) | 7.8 ± 1.4b (1.1) | 11.5 ± 2.7a (1.6) | 69.7 ± 22.8a (9.6) | 728.6 ± 143.7b |
| Mean | 277.8 ± 104.2 (50.8) | 46.3 ± 26.3 (8.0) | 9.0 ± 3.9 (1.7) | 30.1 ± 13.0 (5.5) | 10.1 ± 6.8 (1.8) | 11.6 ± 4.3 (2.1) | 3.6 ± 2.1 (0.6) | nd | 88.1 ± 19.7 (16.1) | 6.6 ± 2.8 (1.2) | 9.4 ± 2.6 (1.7) | 57.8 ± 18.2 (10.6) | 546.9 ± 190.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karlińska, E.; Romanowska, B.; Kosmala, M. The Aerial Parts of Agrimonia procera Wallr. and Agrimonia eupatoria L. as a Source of Polyphenols, and Especially Agrimoniin and Flavonoids. Molecules 2021, 26, 7706. https://doi.org/10.3390/molecules26247706
Karlińska E, Romanowska B, Kosmala M. The Aerial Parts of Agrimonia procera Wallr. and Agrimonia eupatoria L. as a Source of Polyphenols, and Especially Agrimoniin and Flavonoids. Molecules. 2021; 26(24):7706. https://doi.org/10.3390/molecules26247706
Chicago/Turabian StyleKarlińska, Elżbieta, Beata Romanowska, and Monika Kosmala. 2021. "The Aerial Parts of Agrimonia procera Wallr. and Agrimonia eupatoria L. as a Source of Polyphenols, and Especially Agrimoniin and Flavonoids" Molecules 26, no. 24: 7706. https://doi.org/10.3390/molecules26247706