
Metabolomics Study of Flavonoids of Taxilluschinensis on Different Hosts Using UPLC-ESI-MS/MS

Abstract
:1. Introduction
2. Results and Analysis
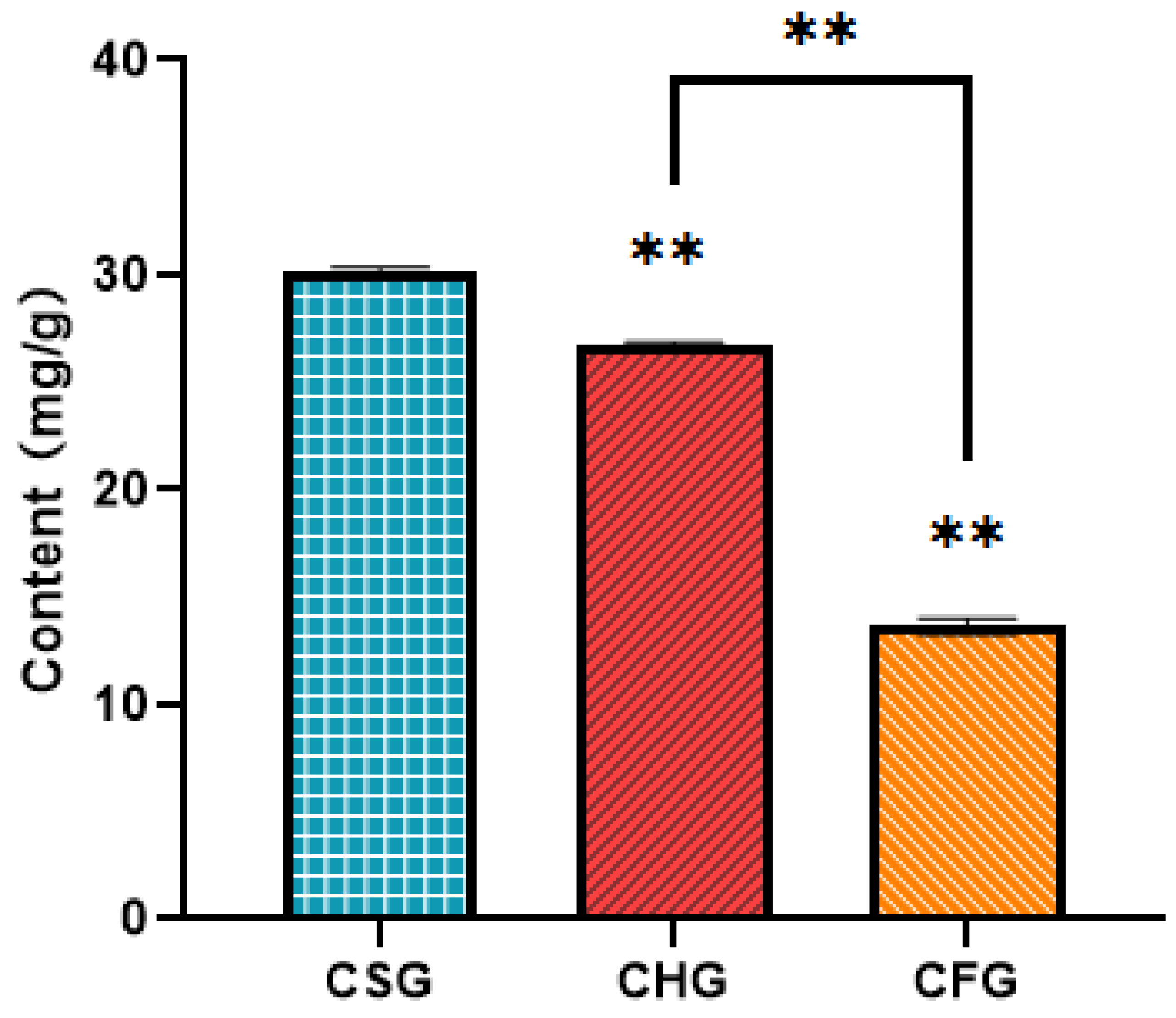
2.1. Quantitative Analysis of Total Flavonoid Content in T. chinensis from Different Hosts
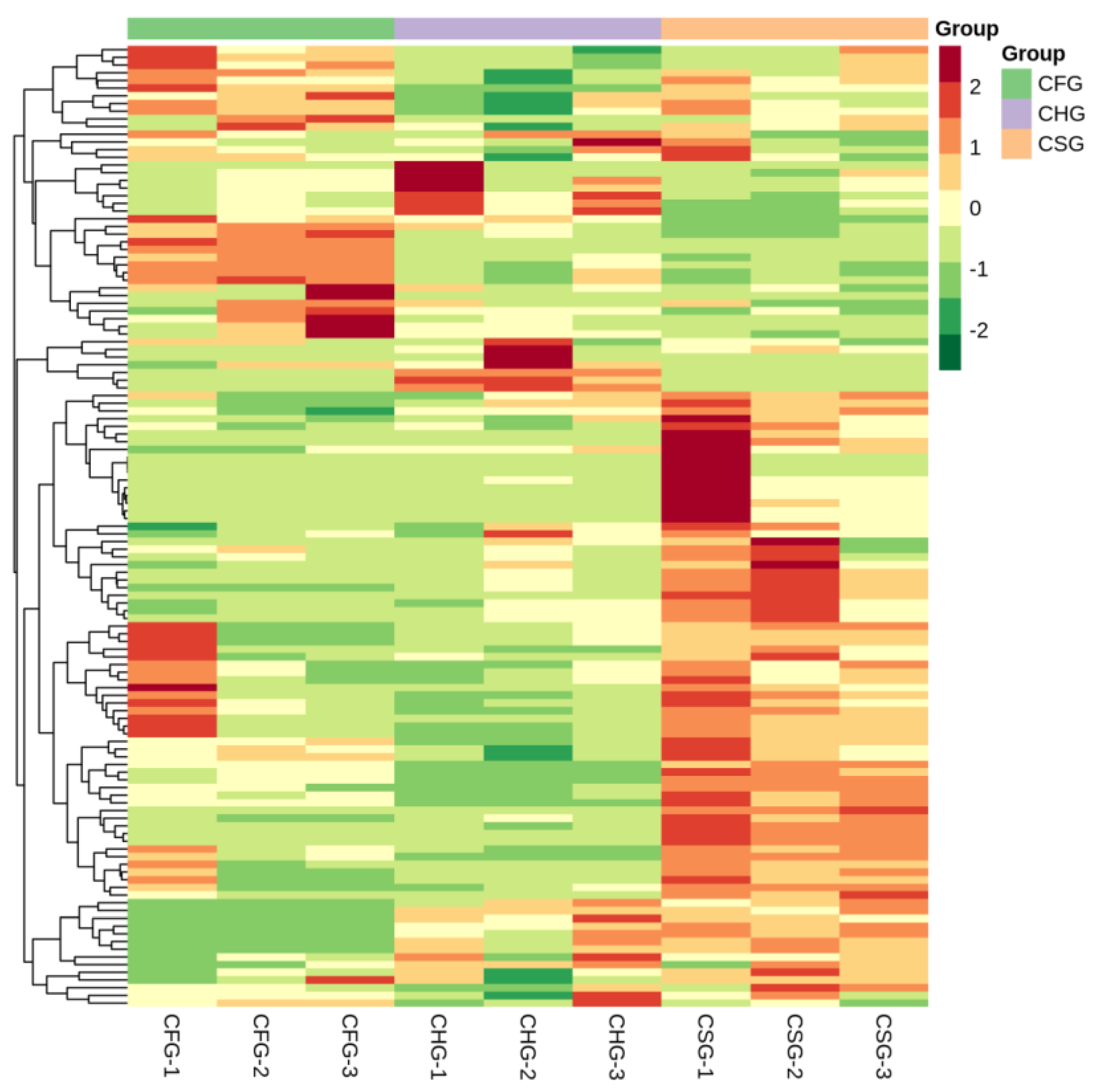
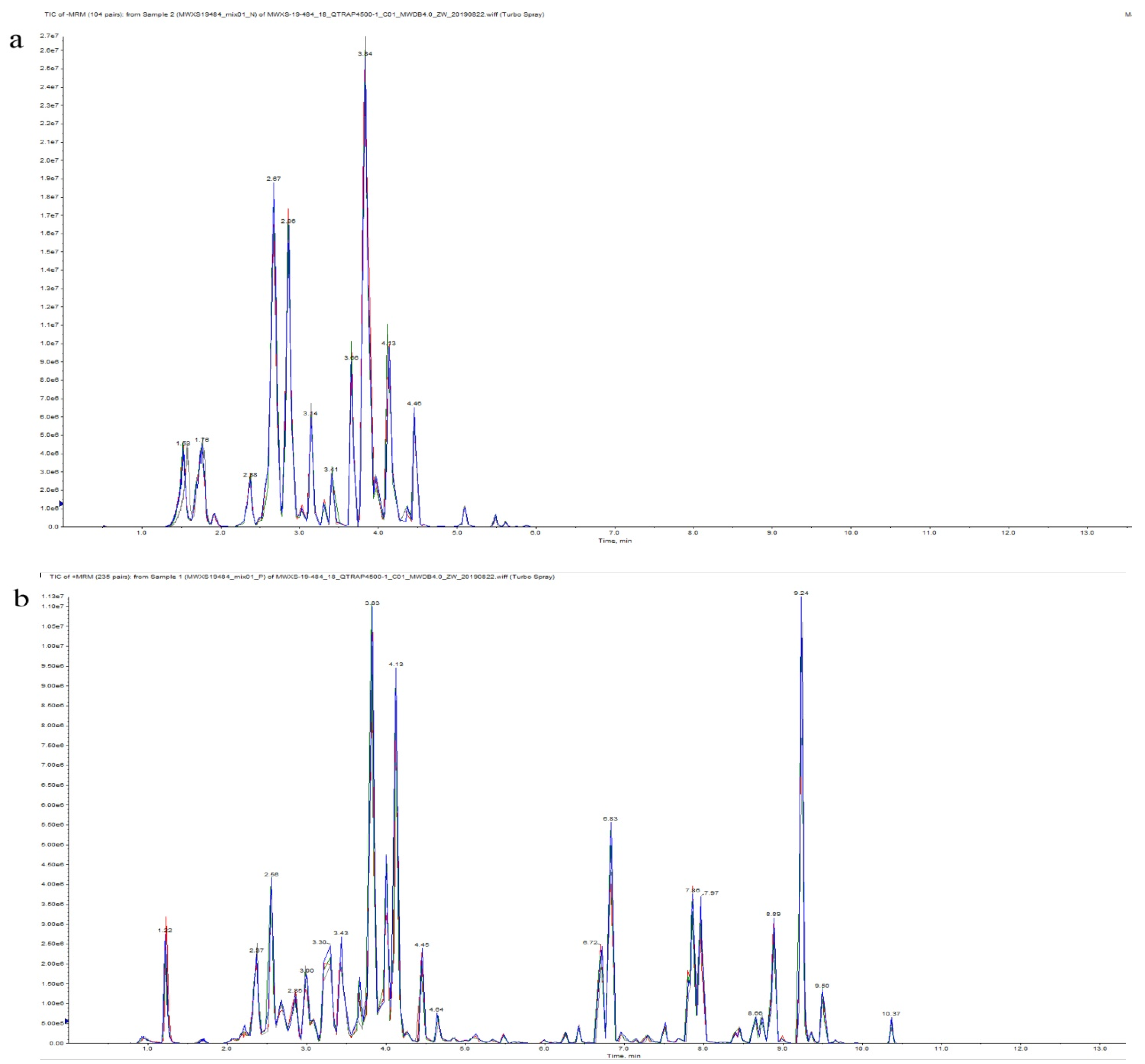
2.2. Comprehensive Analysis of Metabolite Composition
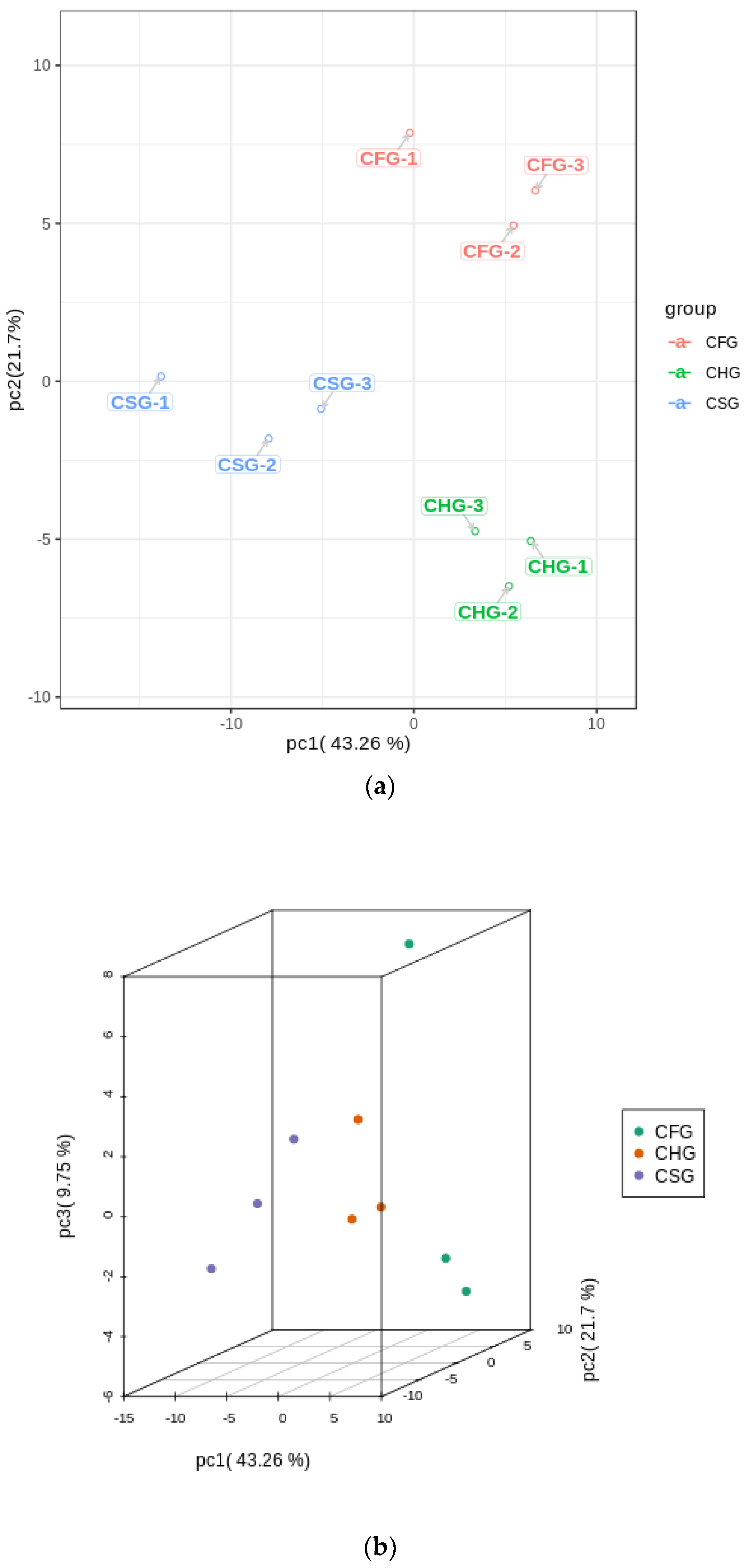
2.3. PCA Results
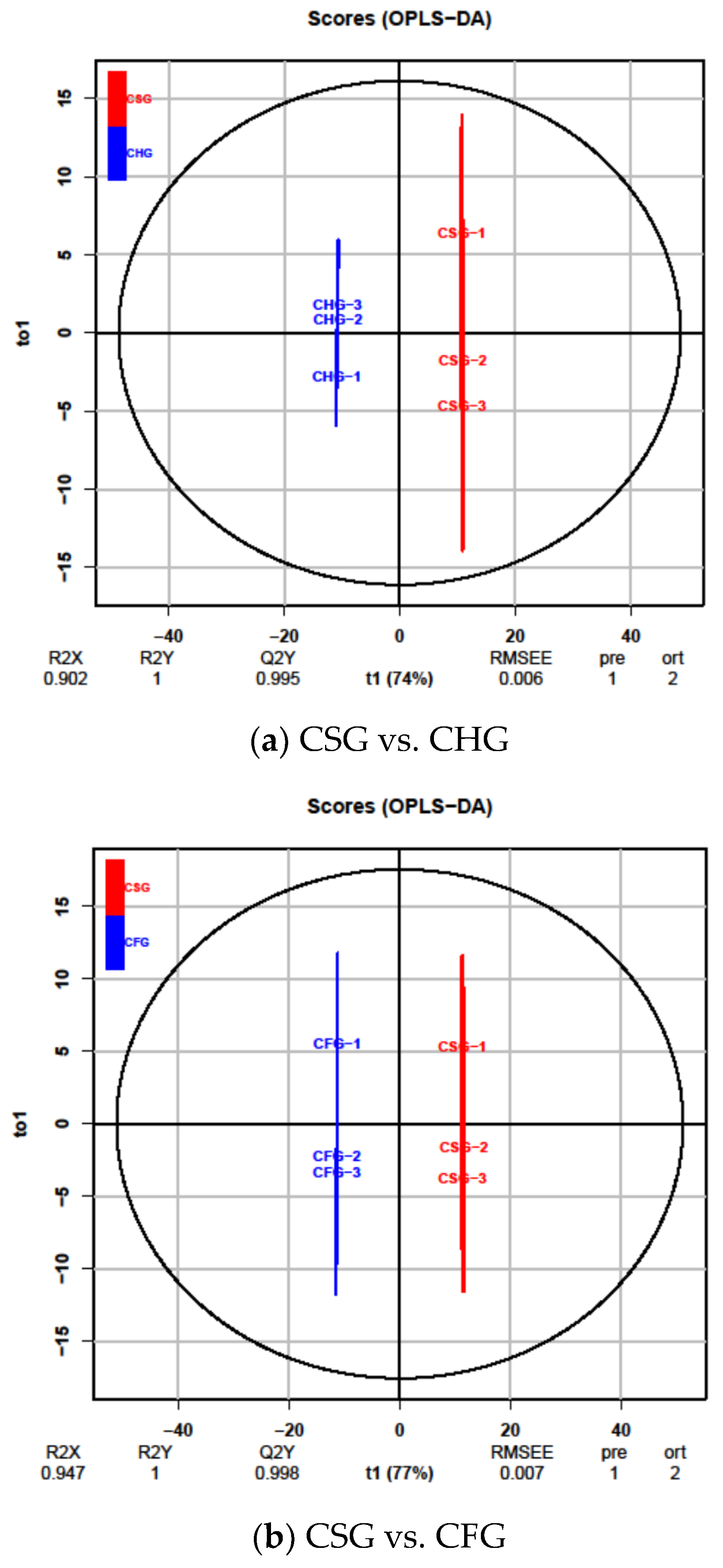
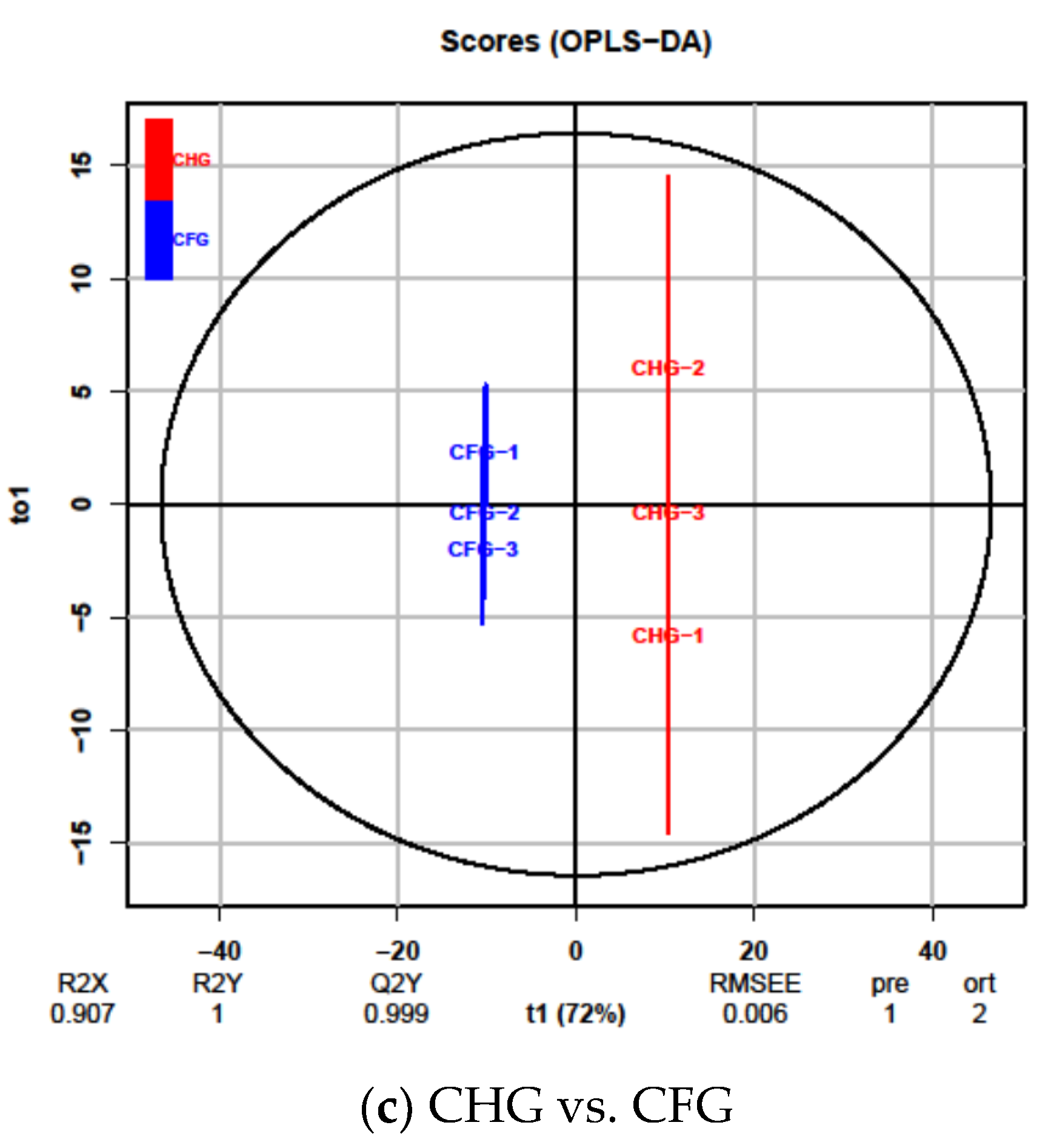
2.4. OPLS-DA Results
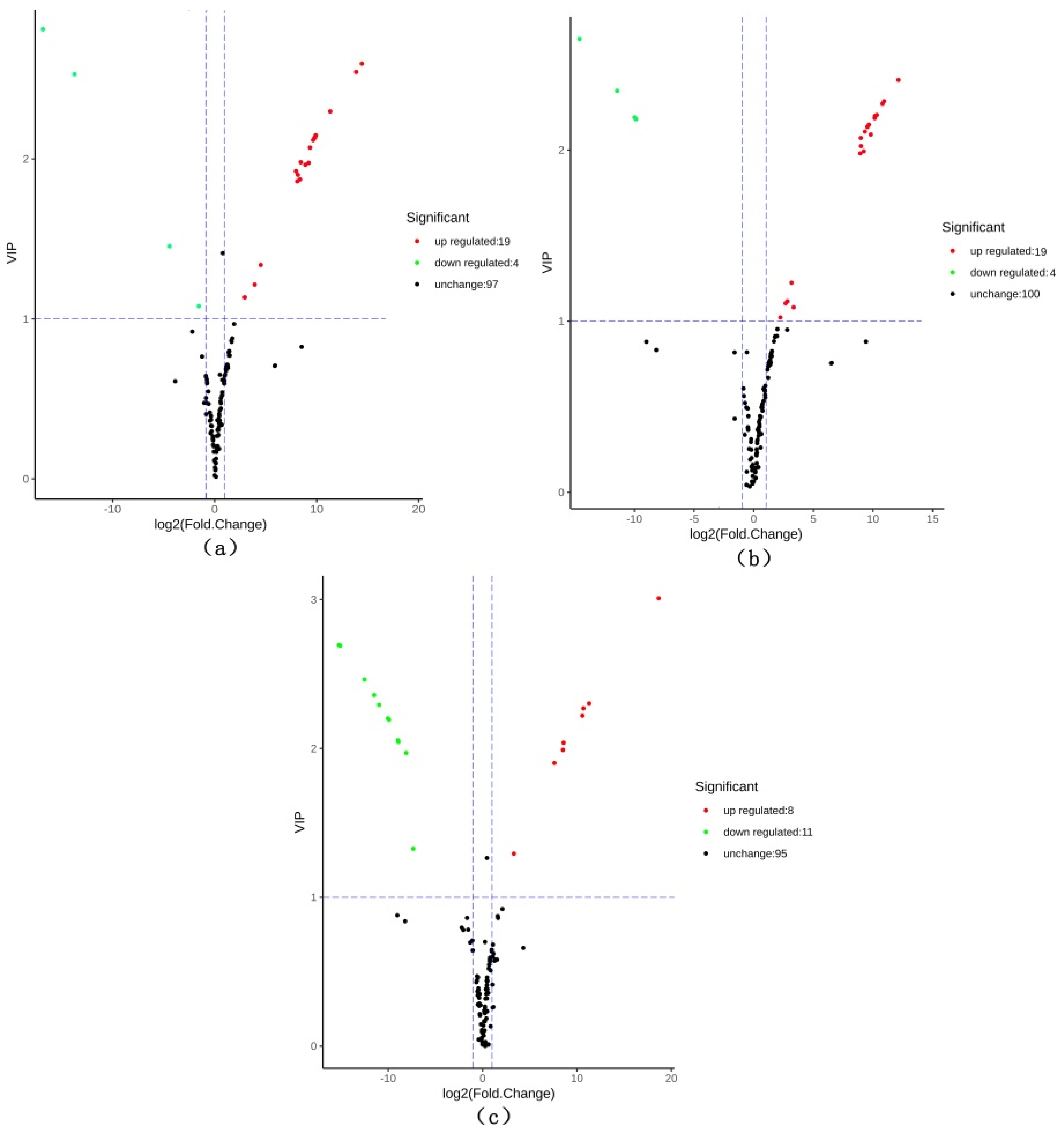
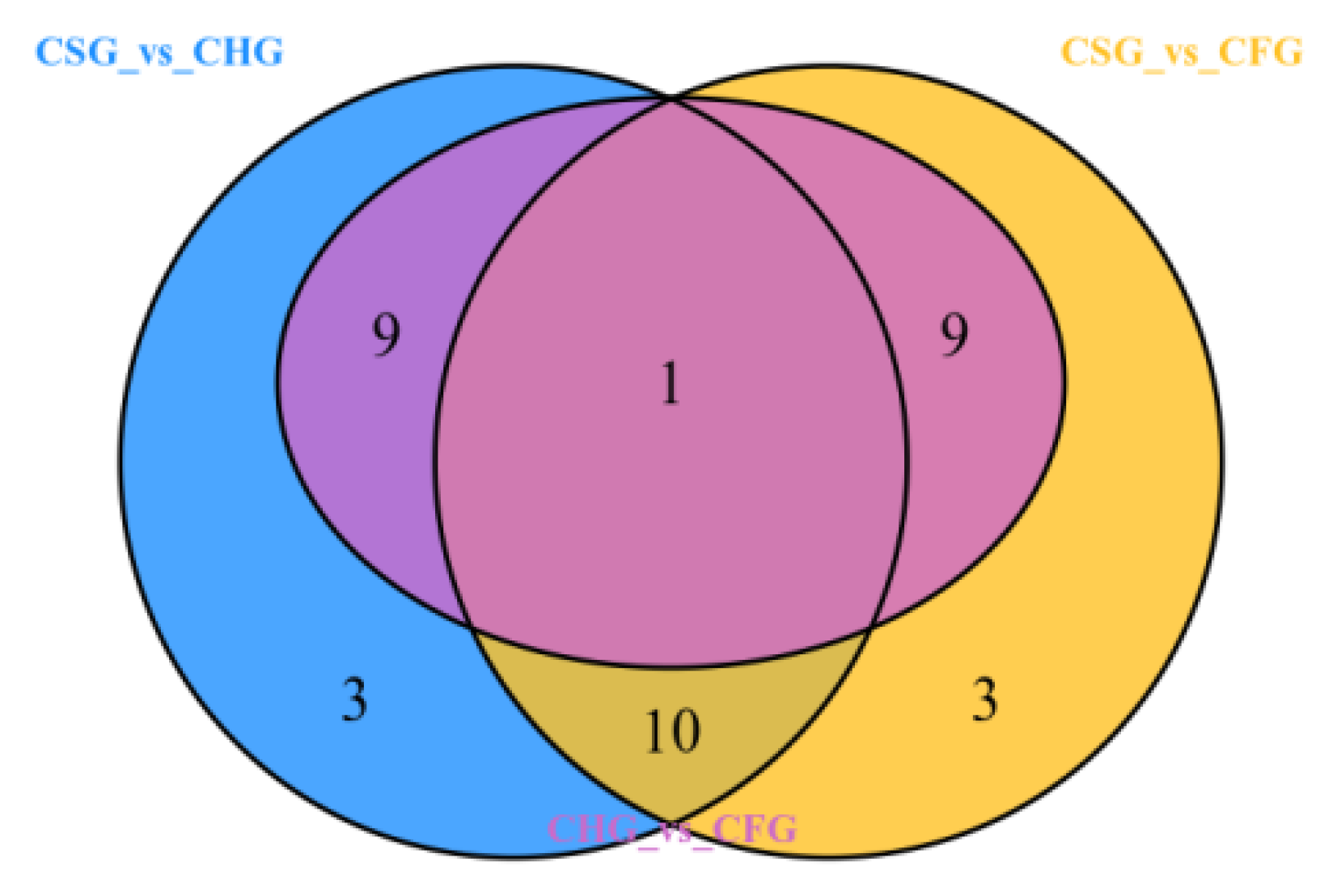
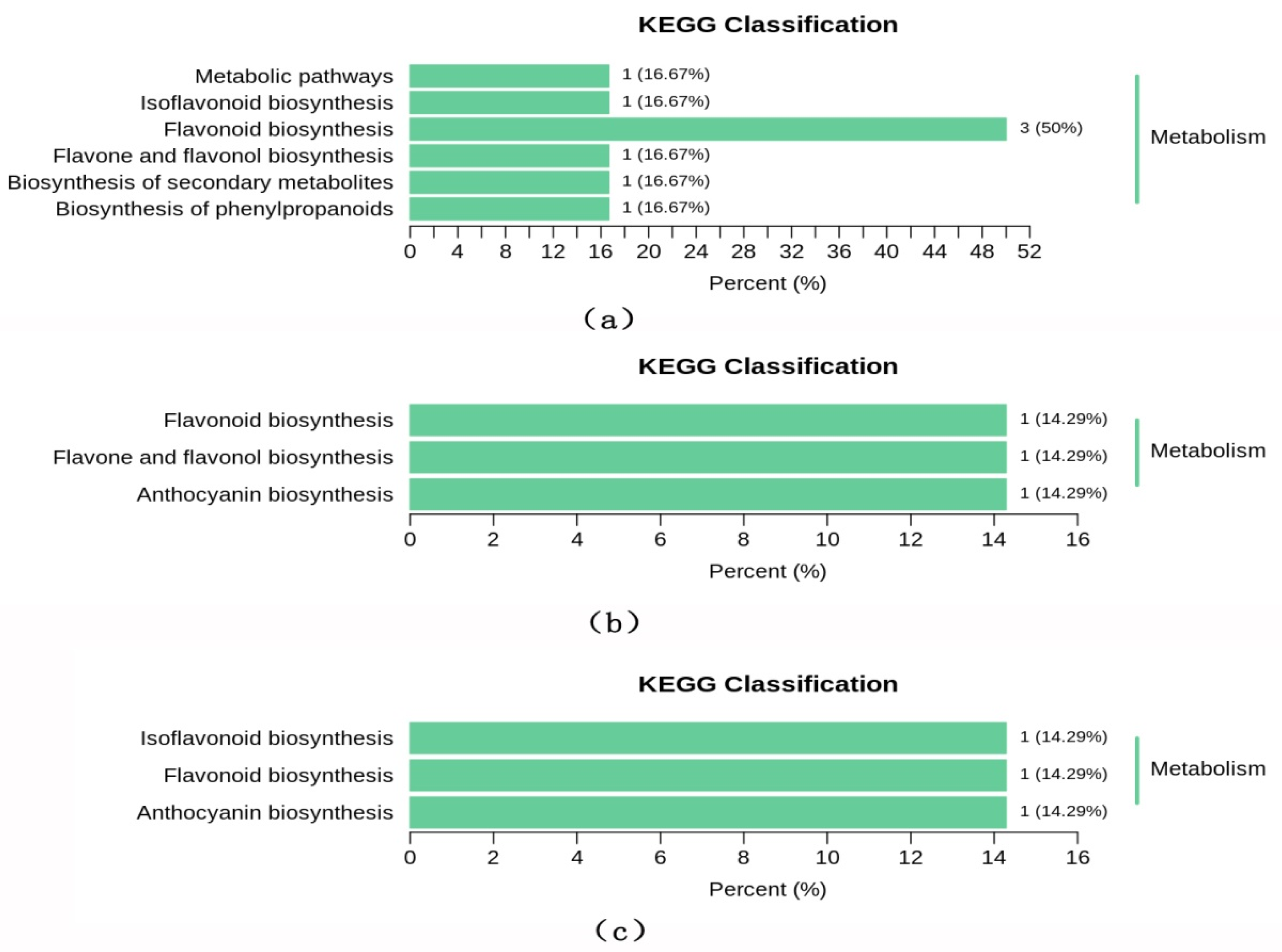
2.5. Differential Metabolite Identification and Screening
3. Discussion
4. Plant Materials and Treatments
4.1. Plant Materials
4.2. Metabolite Extraction
4.3. Calibration Curves
4.4. Determination of Total Flavonoid Content
4.5. LC-MS/MS Analysis of Metabolites
4.6. Quality Control
4.7. Metabolite Identification and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Chinese Pharmacopoeia Commission. Ch. P.; China Medical Science Press: Beijing, China, 2020; Volume I, p. 299.
- Tang, S.W. Classified Materia Medica; Huaxia Publishing House: Beijing, China, 1993. [Google Scholar]
- Li, S.Z. Compendium of Materia Medica; People’s Medical Publishing House: Beijing, China, 1982. [Google Scholar]
- Zhang, B.; Zhou, H.H. Host Plants of Taxillus chinensi. Asia Pac. Tradit. Med. 2017, 13, 81–83. [Google Scholar]
- Wang, J. Sang Parasitic Research Overview. J. Pract. Tradit. Chin. Intern. Med. 2018, 32, 74–77. [Google Scholar]
- Li, Y.H.; Chen, S.L.; Lu, D.; Zhu, K.X.; Zhao, M.H.; Pei, H.H.; Ruan, J.L. Study on the Contents of Total Flavonoids of Herba Taxilli from Different Host-plants. Lishizhen Med. Mater. Med. Res. 2009, 20, 3009–3010. [Google Scholar]
- Li, L.L.; Wang, J.; Cui, Y.; Wen, P.; Guan, J.; Yang, S.; Ma, K. Antirheumatic substance and meridian tropism of Loranthus parasiticus based on “syndrome-efficacy-analysis of biological samples”. China J. Chin. Mater. Med. 2016, 41, 1933–1939. [Google Scholar]
- Zhao, H.W.; Wang, J.; Cui, Y.; Wang, H.J.; Wang, J.M.; Ma, K. A Research of Effective Chemicals of Taxillus chinensis Danser on Osteoporosis and Channel Tropism Based on Disease-Effect-Bioanalysis. World Sci. Technol./Mod. Tradit. Chin. Medwinc Mater. Med. 2016, 18, 626–631. [Google Scholar]
- Li, L.L.; Guan, J.; Feng, J.; Ma, K.; Cui, Y. Study on the anti-rheumatism effective substances and meridian tropism of the total flavonoids of Taxillus chinensis (DC.) based on ‘syndrome-efficacy-analysis of biological samples’. CJTCMP 2018, 33, 2203–2208. [Google Scholar]
- Park, G.H.; Song, H.M.; Su, B.P.; Ji, H.P.; Shin, M.S.; Son, H.J.; Um, Y.; Jin, B.J. Anti-Proliferative Activity of Ethanol Extracts from Taxilli Ramulus (Taxillus chinensis (DC.) Danser) Through Cyclin D1 Proteasomal Degradation in Human Colorectal Cancer Cells. Korean J. Plant Resour. 2017, 30, 640–646. [Google Scholar]
- Zhang, J.; Zhou, X.X.; Ling, Y.; Su, D. Study on in vitro anti-leukemia effects of different extracts of Taxillus chinensis. Shizhen Med. Mater. Med. Res. 2011, 22, 2452–2454. [Google Scholar]
- Wang, Y.; Zhang, S.Y.; Ma, X.F.; Tian, W.X. Potent inhibition of fatty acid synthase by parasitic loranthus [Taxillus chinensis (dc.) danser] and its constituent avicularin. J. Enzym. Inhib. Med. Chem. 2006, 21, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Sundar, R.K.; Sang, C.J.; Narsimha, R.; Trevor, B.; Longvah, T. Immunomodulatory activities of polysaccharides isolated from Taxillus chinensis and Uncaria rhyncophylla. Carbohydr. Polym. 2013, 98, 1458–1465. [Google Scholar]
- Ahn, S.J.; Baek, J.M.; Cheon, Y.H.; Park, S.H.; Kim, J.Y.; Lee, M.S.; Oh, J.; Min, K.C. Inhibitory Effect of Taxillus chinensis Extract on Osteoclast Differentiation and Bone Resorption. PLoS ONE 2014, 7, e51750. [Google Scholar]
- Zhang, H.; Huang, F.Y.; Liu, R.Y.; Lu, H.L.; Su, B.W.; Zhu, K.X.; Li, Y.H. Effects of Taxilli Herba from different hosts on lowering blood pressure of spontaneously hypertensive rats. Chin. Tradit. Pat. Med. 2018, 40, 249–254. [Google Scholar]
- Yoo, J.M.; Yang, J.H.; Kim, Y.S.; Cho, W.K.; Ma, J.Y. Inhibitory Effect of Loranthus parasiticus on IgE-Mediated Allergic Responses in RBL-2H3 Cells. Mediat. Inflamm. 2016, 2016, 8742562. [Google Scholar] [CrossRef] [Green Version]
- Weon, J.B.; Lee, J.; Eom, M.R.; Jung, Y.S.; Ma, C.J. The Effects of Loranthus parasiticus on Scopolamine-Induced Memory Impairment in Mice. Evid. Based Complement. Altern. Med. 2014, 2014, 860180. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.Z.H.; Kadir, H.A.; Lee, C.L.; Hing, G.B. Neuroprotective properties of Loranthus parasiticus aqueous fraction against oxidative stress-induced damage in NG108-15 cells. J. Nat. Med. 2012, 66, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.H.; Huang, J.J.; Zhu, K.X.; Su, B.W.; Zhao, M.H.; Zhang, M.F. Prospect of the Precious Genuine Traditional Chinese Medicine Taxilli Herba. China Pharm. 2016, 25, 13–16. [Google Scholar]
- Li, B.; Lu, R.M.; Chen, H.; Miao, W.S. Determination of total flavonoids in Taxillus chinensis from two different hosts. Asia-Pac. Tradit. Med. 2010, 6, 37–38. [Google Scholar]
- Huang, F.Y.; Liu, R.Y.; Lu, H.L.; Li, Y.H. Effects of different medicinal herbs on the content of total flavonoids in Taxillus chinensis. J. Guangxi Univ. Chin. Med. 2017, 20, 34–36. [Google Scholar]
- Qing, Y.R.; Li, H.S.; Liu, X.H.; Lin, S.H.; Chen, F.J.; Luo, Z.Y. Seasonal differences of total flavonoids content in different hosts of Loranthaceae in northwest Guangxi. J. Hechi Univ. 2014, 34, 1–7. [Google Scholar]
- Jia, N.F.; Tan, W.W.; Li, B.; Chen, S.F.; Lu, R.M. The content of total flavonoids Taxillus chinensis in different hosts was determined by ultraviolet spectrophotometry. Guangxi J. Tradit. Chin. Med. 2013, 36, 79–80. [Google Scholar]
- Zhang, X.J.; Zhu, K.X.; Zhao, M.H.; Pei, H.H.; Li, Y.H. Content Analysis of Quercitrin and Quercetin in Herba Taxilli from Different Host-Plants. Lishizhen Med. Mater. Med. Res. 2011, 22, 1604–1606. [Google Scholar]
- Hanieh, H.; Hairul, I.V.I.; Saravanan, S.; Chellappandian, M.; Ragul, K.; Durga, A.; Venugopal, K.; Senthilkumar, V.; Senthilkumar, P.; Thirugnanasambantham, K. Pinocembrin, a novel histidine decarboxylase inhibitor with anti-allergic potential in in vitro. Eur. J. Pharmacol. 2017, 814, 178–186. [Google Scholar] [CrossRef]
- Tao, J.; Shen, C.; Sun, Y.; Chen, W.; Yan, G.F. Neuroprotective effects of pinocembrin on ischemia/reperfusion-induced brain injury by inhibiting autophagy. Biomed. Pharmacother. 2018, 106, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Gong, H. Pinocembrin suppresses proliferation and enhances apoptosis in lung cancer cells in vitro by restraining autophagy. Bioengineered 2021, 12, 6035–6044. [Google Scholar] [CrossRef]
- Innajak, S.; Tangchirakhaphan, S.; Nilwarangoon, S.; Tanjapatkul, N.; Mahabusarkam, W.; Watanapokasin, R. Anti-Proliferation and Apoptosis Induction in Epidermoid Carcinoma A431 Cells by Artonin E. J. Med. Assoc. Thai 2017, 100, 54. [Google Scholar]
- Christopher, E.I.; Rasedee, A.; Arifah, K.; Mohd, H.N.; Keong, Y.S.; Umar, I.M.; Faiqah, R.; Ibrahim, M.; Lim, L.K.; Ubong, E. The molecular mechanism of the anticancer effect of Artonin E in MDA-MB 231 triple negative breast cancer cells. PLoS ONE 2017, 12, e0182357. [Google Scholar]
- Wang, Y.H. Studies on the Bioactive Constituents of Two Moraceous Medicinal Plants; Fudan University: Shanghai, China, 2004. [Google Scholar]
- Chen, K.F.; Ou, Y.S.; Wen, H.L.; Shen, Z.J.; Xie, P. Newly discovered flavonoids and their bioactivity during 2004–2008 of Artocarpus. Jiangxi J. Tradit. Chin. Med. 2009, 40, 71–73. [Google Scholar]
- Wang, Y.; Wei, H.; Yang, G.; Zhang, M. Effect of sanggenon D on the growth of tumor cells and transplantation tumors. Nat. Prod. Res. Dev. 2020, 32, 1057–1064. [Google Scholar]
- Nam, M.S.; Jung, D.B.; Seo, K.H.; Kim, B.I.; Kim, J.H.; Kim, J.H.; Kim, B.; Baek, N.I.; Kim, S.H. Apoptotic Effect of Sanggenol L via Caspase Activation and Inhibition of NF-κB Signaling in Ovarian Cancer Cells. Phytother. Res. 2016, 30, 90–96. [Google Scholar] [CrossRef]
- Won, Y.S.; Seo, K.I. Sanggenol L Promotes apoptotic cell death in melanoma skin cancer cells through activation of caspase cascades and apoptosis-inducing factor—ScienceDirect. Food Chem. Toxicol. 2020, 138, 111221. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T.; Hano, Y. Constituents of the Chinese crude drug “Sang-Bai-Pi” (Morus root bark). Planta Medica 1983, 47, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Nanjing University of Chinese Medicine. Dictionary of Traditional Chinese Medicine, 2nd ed.; Scientific and Technical Publishers: Shanghai, China, 2006; Volume 2. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Fraga, C.G.; Clowers, B.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography—Mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Rt (min) | Q1 | Q3 | Molecular Weight | Ionization Model | Formula | Compounds |
|---|---|---|---|---|---|---|---|
| pmp000595 | 3.80 | 611.16 | 287.05 | 610.13 | [M+H]+ | C27H30O16 | Luteolin-7,3′-Di-O-beta-d-glucoside |
| mws0789 | 6.00 | 255.10 | 107.01 | 256.06 | [M−H]− | C15H12O4 | Pinocembrin (dihydrochrysin) |
| pmp000836 | 7.40 | 455.20 | 379.12 | 454.14 | [M+H]+ | C25H26O8 | Nigrasins A |
| pmp000821 | 8.00 | 421.20 | 309.04 | 420.14 | [M+H]+ | C25H24O6 | Artonin E2 |
| pmp000815 | 8.60 | 407.20 | 165.02 | 406.16 | [M+H]+ | C25H26O5 | Kuwanon S2 |
| pmb0663 | 4.20 | 611.10 | 383.08 | 610.15 | [M+H]+ | C27H30O16 | 8-C-hexosyl-luteolin O-hexoside |
| pmp000809 | 6.00 | 355.10 | 229.05 | 354.10 | [M+H]+ | C20H18O6 | Sanggenon F/H |
| pmp000829 | 6.70 | 437.20 | 363.09 | 436.13 | [M+H]+ | C25H24O7 | Sanggenon M |
| pmp000825 | 8.00 | 423.20 | 153.02 | 422.15 | [M+H]+ | C25H26O6 | Kuwanon D |
| pmp000822 | 7.00 | 423.20 | 311.07 | 422.15 | [M+H]+ | C25H26O6 | Sanggenol L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Teng, J.; Zhu, Y.; Xie, F.; Hou, J.; Ling, Y.; Zhu, H. Metabolomics Study of Flavonoids of Taxilluschinensis on Different Hosts Using UPLC-ESI-MS/MS. Molecules 2021, 26, 7681. https://doi.org/10.3390/molecules26247681
Li L, Teng J, Zhu Y, Xie F, Hou J, Ling Y, Zhu H. Metabolomics Study of Flavonoids of Taxilluschinensis on Different Hosts Using UPLC-ESI-MS/MS. Molecules. 2021; 26(24):7681. https://doi.org/10.3390/molecules26247681
Chicago/Turabian StyleLi, Li, Jianbei Teng, Yilin Zhu, Fengfeng Xie, Jing Hou, Yuan Ling, and Hua Zhu. 2021. "Metabolomics Study of Flavonoids of Taxilluschinensis on Different Hosts Using UPLC-ESI-MS/MS" Molecules 26, no. 24: 7681. https://doi.org/10.3390/molecules26247681