
Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.)

 ,
,
Abstract
:1. Introduction
2. Results
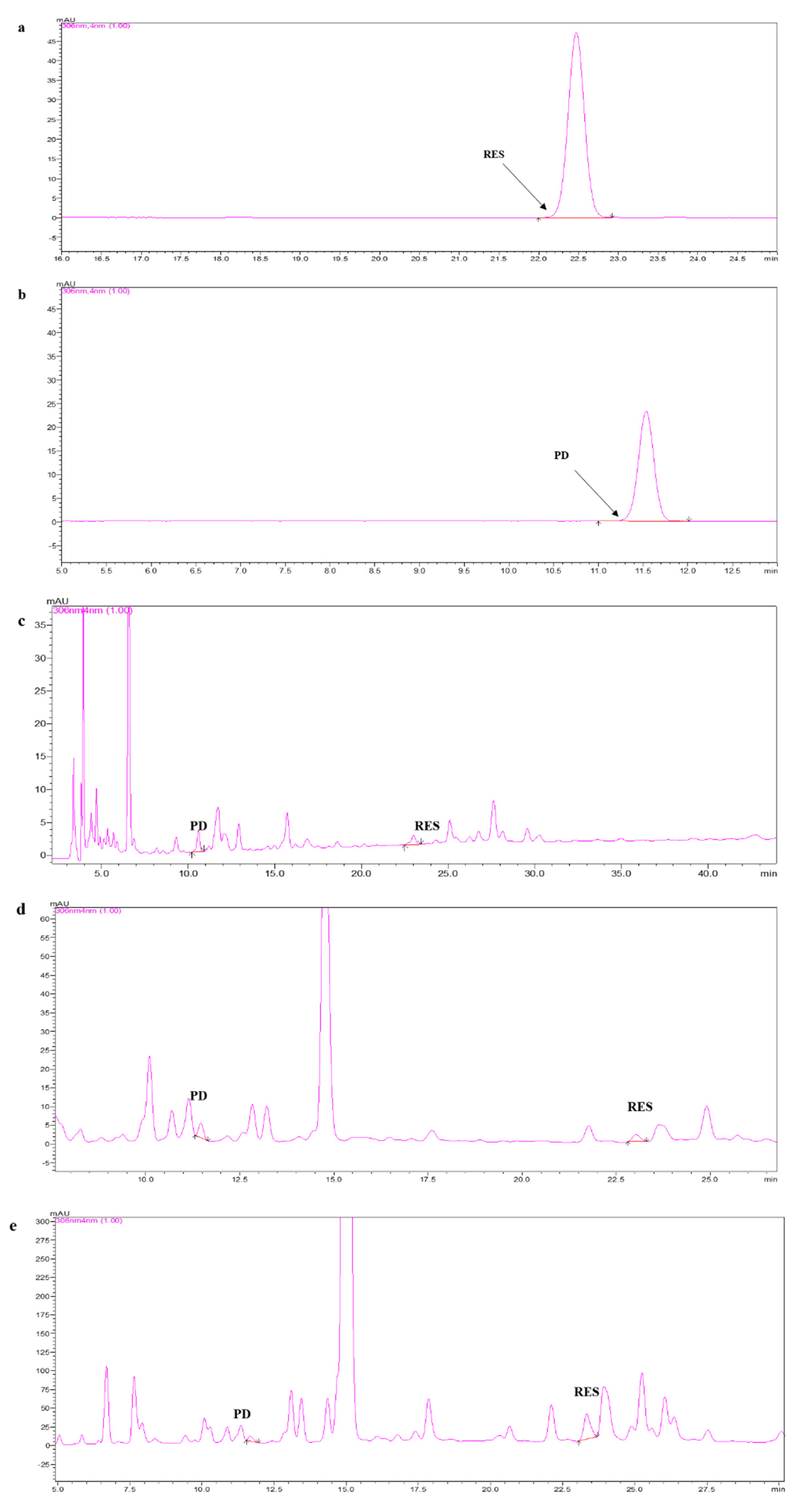
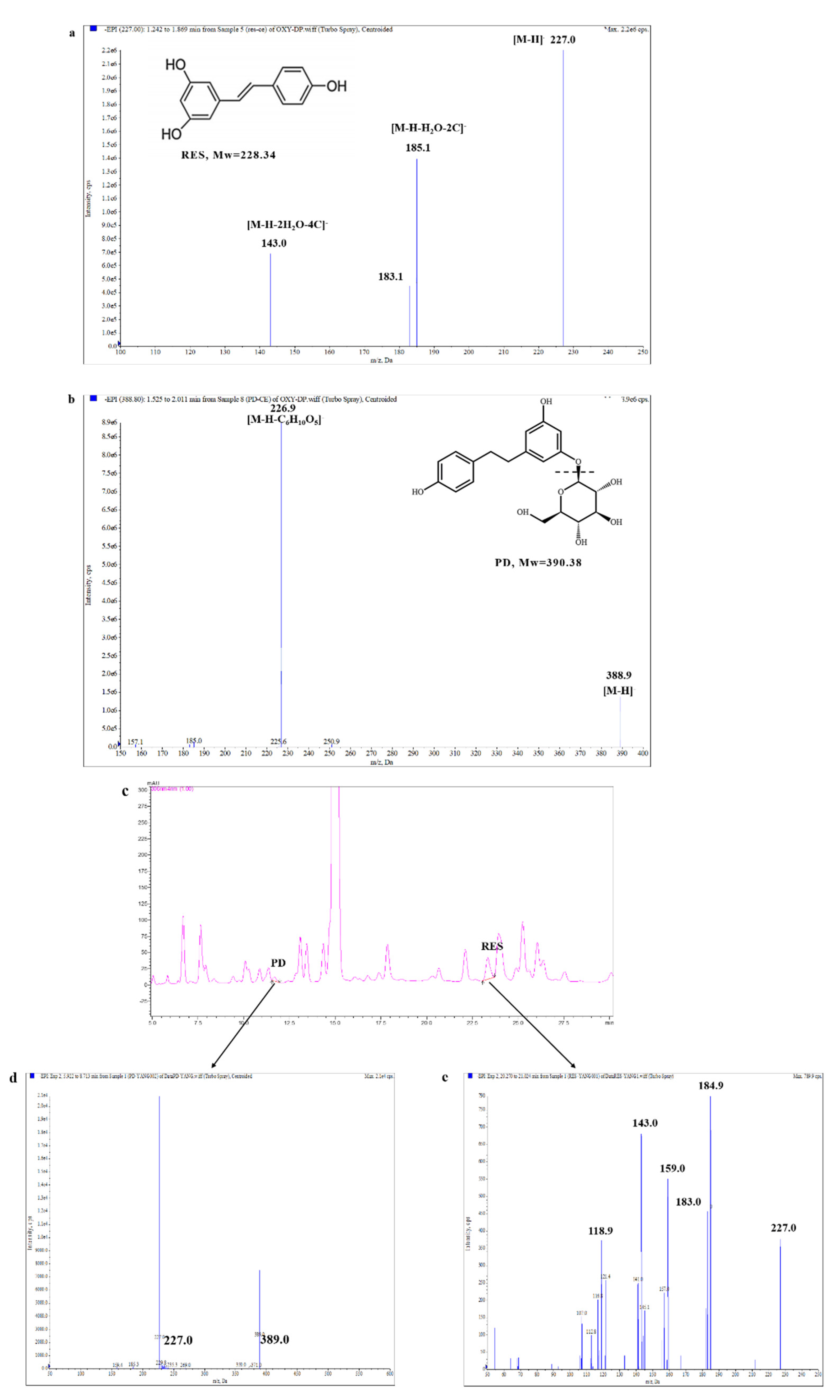
2.1. Separation and Identification of RES and PD
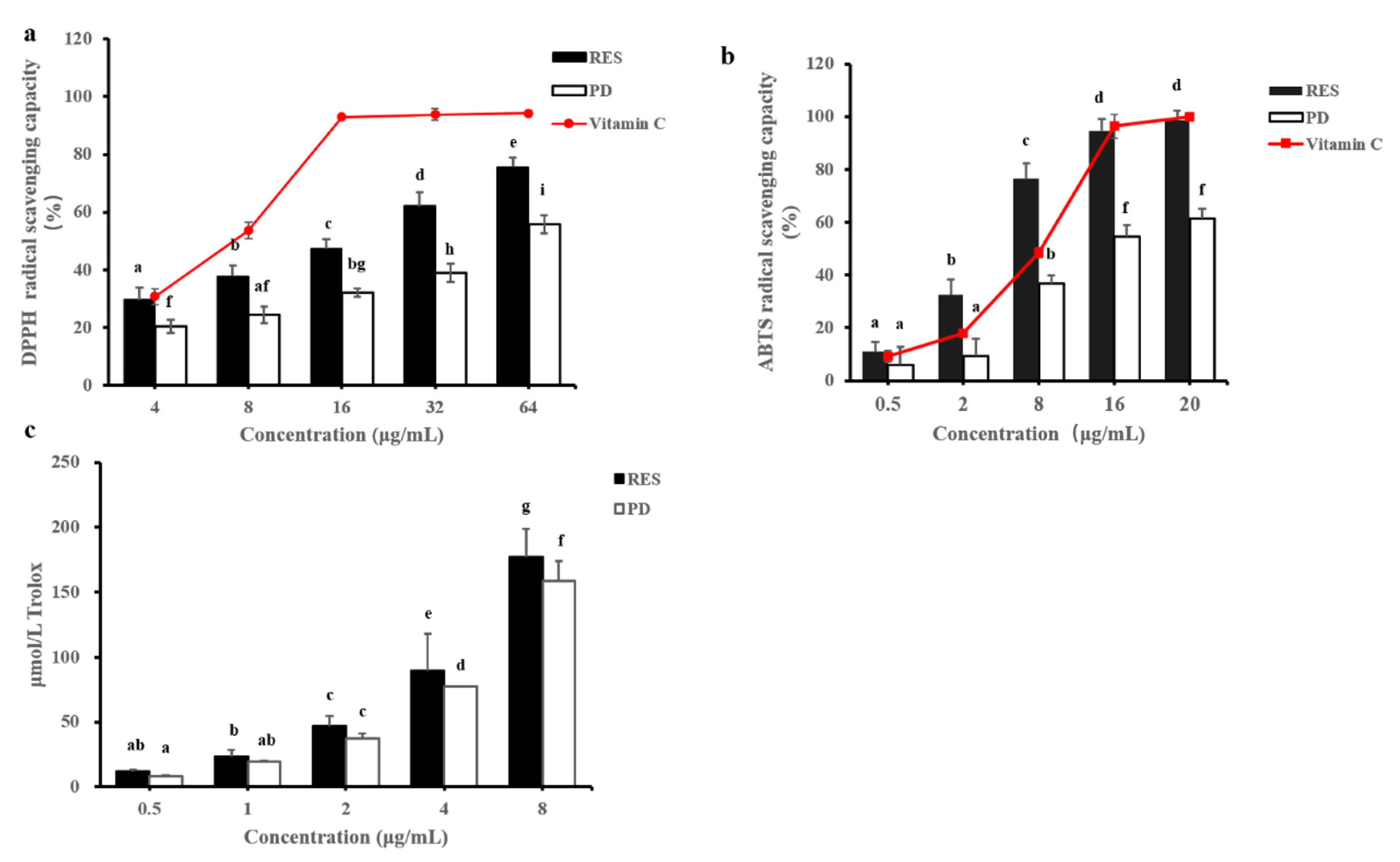
2.2. In Vitro Antioxidant Activities
2.2.1. DPPH Free Radical-Scavenging Activity
2.2.2. ABTS Free Radical-Scavenging Activity
2.2.3. ORAC (Oxygen Radical Absorption Capacity)
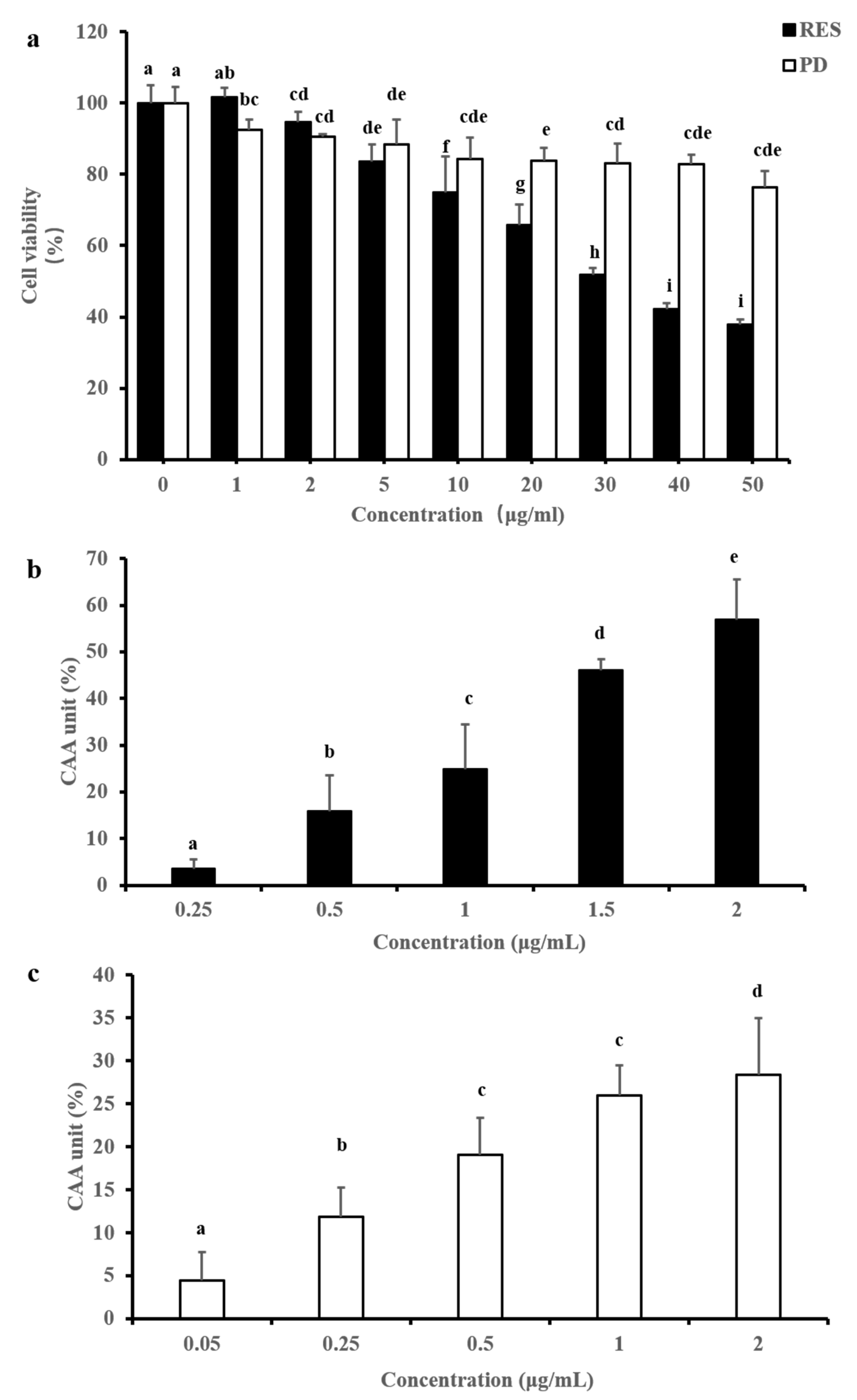
2.3. CAA
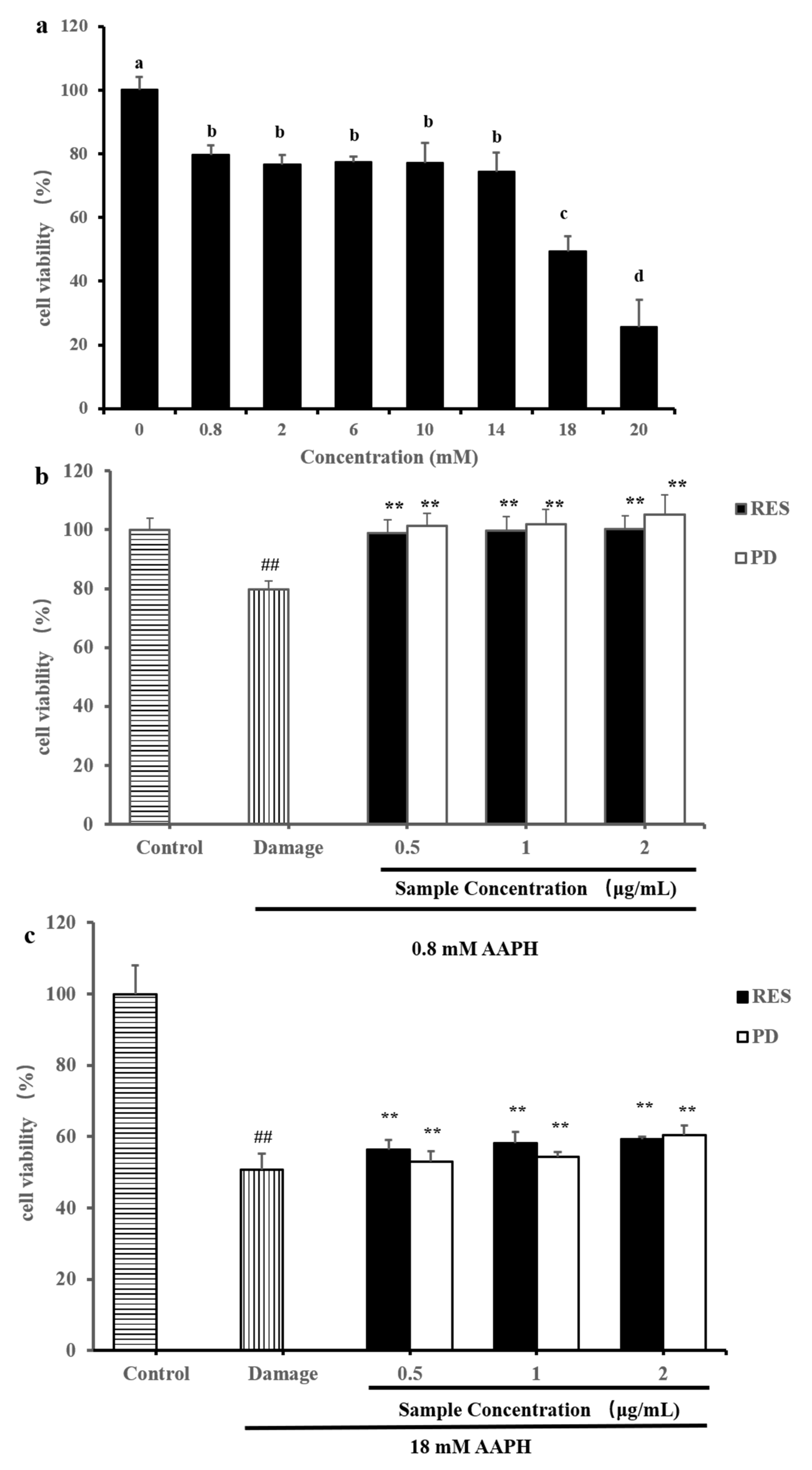
2.4. Protective Effects of RES and PD Compounds on HepG2 against AAPH-Induced Oxidative Stress
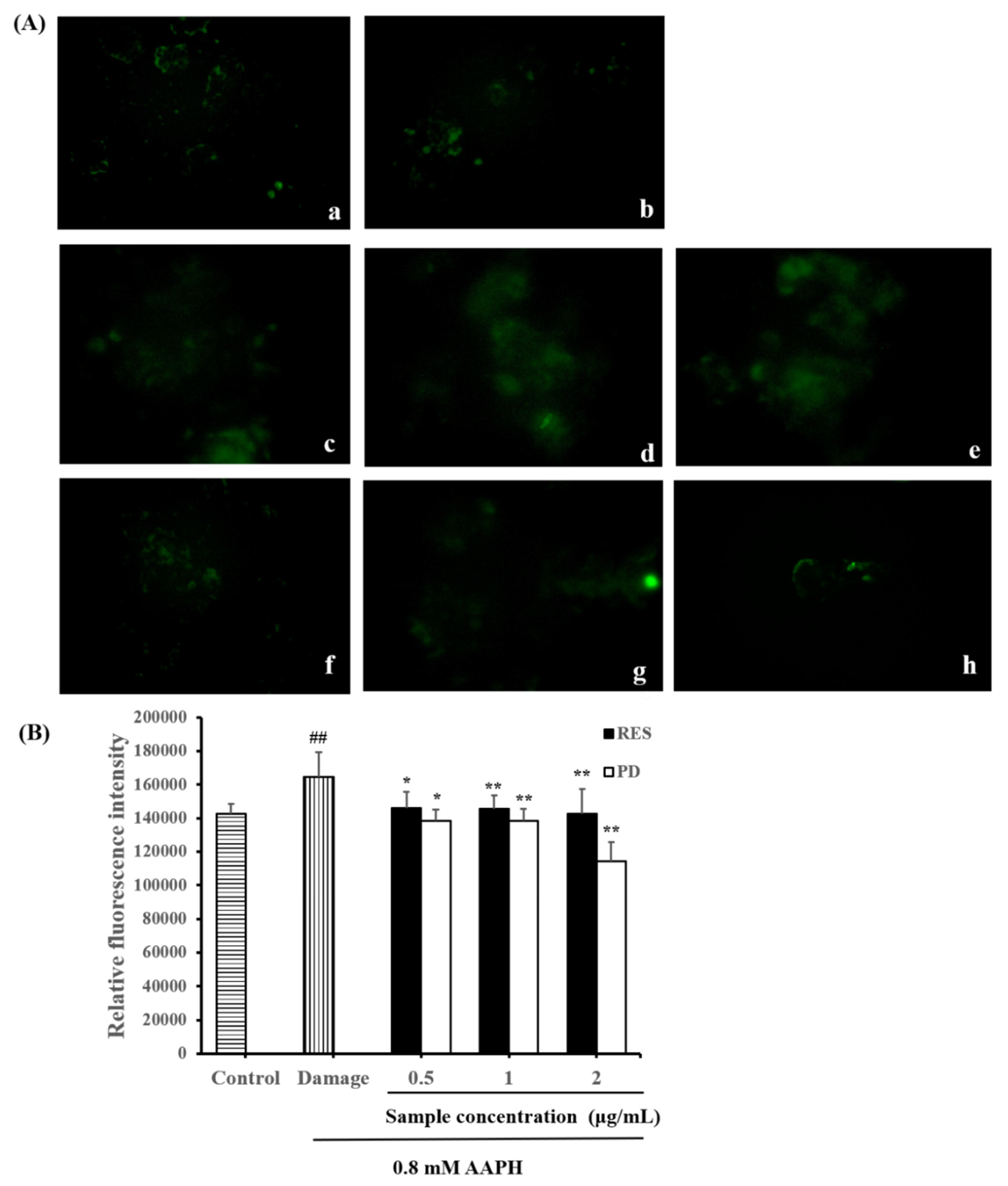
2.5. Effects of RES and PD on ROS Level in AAPH-Induced HepG2 Cell Damage
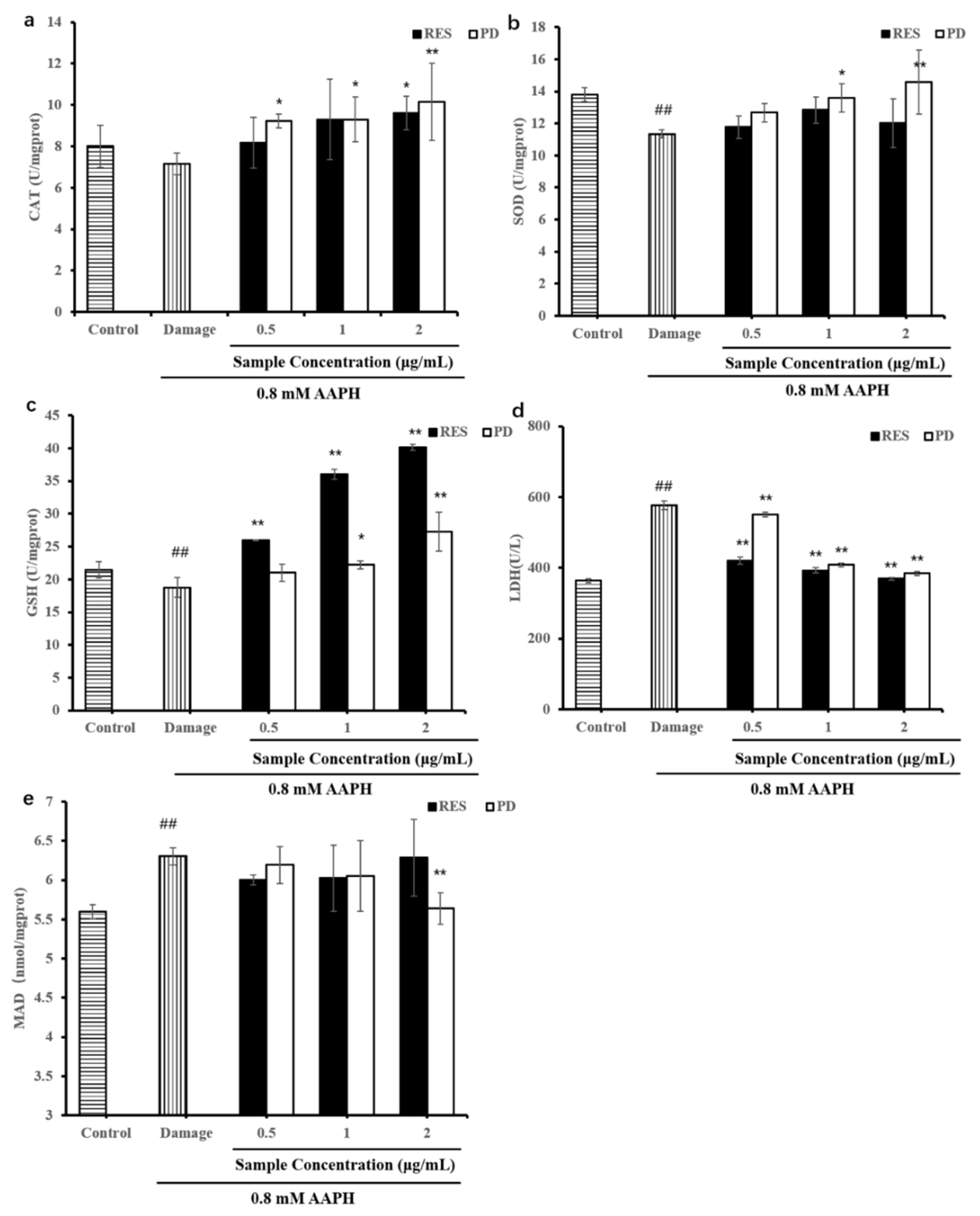
2.6. Levels of Antioxidant Enzymes (SOD, LDH, CAT, and GSH) and MDA
3. Materials and Methods
3.1. Chemicals and Materials
3.2. Extraction of RES and PD
3.3. Separation and Analysis of RES and PD
3.4. In Vitro Antioxidant Activities
3.4.1. DPPH Free Radical-Scavenging Activity
3.4.2. ABTS Free Radical-Scavenging Activity
3.4.3. ORAC Assays
3.5. Cells Lines and Culture Conditions
3.5.1. MTT Assay
3.5.2. CAA
3.6. Measurement of Protect Effect against AAPH-Induced Oxidative Stress
3.7. ROS Level
3.8. Measurement of GSH, MDA Levels and Cell Antioxidant Enzyme (LDH, SOD, and CAT) Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gupta, E.; Mishra, P. Functional Food with some Health Benefits are so Called as Superfood—A Review. Curr. Nutr. Food Sci. 2020, 16. [Google Scholar] [CrossRef]
- Saini, P.; Kumar, N.; Kumar, S.; Mwaurah, P.W.; Panghal, A.; Attkan, A.K.; Singh, V.K.; Garg, M.K.; Singh, V. Bioactive compounds, nutritional benefits and food applications of colored wheat: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2020, 61, 3197–3210. [Google Scholar] [CrossRef]
- Rakić, S.; Povrenović, D.; Tešević, V.; Simić, M.; Maletić, R. Oak acorn, polyphenols and antioxidant activity in functional food. J. Food Eng. 2006, 74, 416–423. [Google Scholar] [CrossRef]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 3845–3925. [Google Scholar] [CrossRef]
- Machado, A.; Geraldi, M.V.; do Nascimento, R.P.; Moya, A.; Vezza, T.; Diez-Echave, P.; Galvez, J.J.; Cazarin, C.B.B.; Marostica Junior, M.R. Polyphenols from food by-products: An alternative or complementary therapy to IBD conventional treatments. Food Res. Int. 2021, 140, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Mal, G.; Sharma, D.; Sharma, R.; Antony, C.P.; Kalra, R.S. Gastrointestinal biotransformation of phytochemicals: Towards futuristic dietary therapeutics and functional foods. Trends. Food Sci. Technol. 2020, 106, 64–77. [Google Scholar] [CrossRef]
- Yu, Q.; Fang, C.; Ma, Y.; He, S.; Ajuwon, K.M.; He, J. Dietary resveratrol supplement improves carcass traits and meat quality of Pekin ducks. Poult Sci. 2021, 100, 1–8. [Google Scholar] [CrossRef]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety studies conducted on high-purity trans-resveratrol in experimental animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef]
- Abdollahi, S.; Salehi-Abargouei, A.; Toupchian, O.; Sheikhha, M.H.; Fallahzadeh, H.; Rahmanian, M.; Tabatabaie, M.; Mozaffari-Khosravi, H. The Effect of Resveratrol Supplementation on Cardio-Metabolic Risk Factors in Patients with Type 2 Diabetes: A Randomized, Double-Blind Controlled Trial. Phytother. Res. 2019, 33, 3153–3162. [Google Scholar] [CrossRef]
- Mirhadi, E.; Roufogalis, B.D.; Banach, M.; Barati, M.; Sahebkar, A. Resveratrol: Mechanistic and therapeutic perspectives in pulmonary arterial hypertension. Pharmacol. Res. 2021, 163, 1–11. [Google Scholar] [CrossRef]
- Huminiecki, L.; Atanasov, A.G.; Horbanczuk, J. Etiology of atherosclerosis informs choice of animal models and tissues for initial functional genomic studies of resveratrol. Pharmacol. Res. 2020, 156, 1–12. [Google Scholar] [CrossRef]
- Montanari, S.; Davani, L.; Tumiatti, V.; Dimilta, M.; Gaddi, A.V.; De Simone, A.; Andrisano, V. Development of an UHPLC-diode arrays detector (DAD) method for the analysis of polydatin in human plasma. J. Pharmaceut. Biomed. 2021, 198, 1–7. [Google Scholar] [CrossRef]
- Song, X.; Cui, L.; Li, J.; Yan, H.; Li, L.; Wen, L.; Geng, Y.; Wang, D. A novel bioreactor for highly efficient biotransformation of resveratrol from polydatin with high-speed counter-current chromatography. LWT-Food Sci. Technol. 2019, 103, 192–198. [Google Scholar] [CrossRef]
- Fan, X.-H.; Wang, L.-T.; An, J.-Y.; Zhang, S.-D.; Cai, Z.-H.; Niu, L.-L.; Kou, P.; Yang, Q.; Meng, D.; Fu, Y.-j. Magnetically immobilized edible Bacillus natto for the biotransformation of polydatin to resveratrol and its bioactivity assessment. Ind. Crops Prod. 2021, 161, 1–7. [Google Scholar] [CrossRef]
- Peng, Y.; Xu, J.; Zeng, Y.; Chen, L.; Xu, X.L. Polydatin attenuates atherosclerosis in apolipoprotein E-deficient mice: Role of reverse cholesterol transport. Phytomedicine 2019, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [Google Scholar] [CrossRef]
- Zhao, X.J.; Yu, H.W.; Yang, Y.Z.; Wu, W.Y.; Chen, T.Y.; Jia, K.K.; Kang, L.L.; Jiao, R.Q.; Kong, L.D. Polydatin prevents fructose-induced liver inflammation and lipid deposition through increasing miR-200a to regulate Keap1/Nrf2 pathway. Redox Biol. 2018, 18, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, C.H.; Jiang, Y.P.; Peng, C.; He, K.; Tang, J.Y.; Xin, H.L. Protective effects of polydatin from Polygonum cuspidatum against carbon tetrachloride-induced liver injury in mice. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Platella, C.; Raucci, U.; Rega, N.; D’Atri, S.; Levati, L.; Roviello, G.N.; Fuggetta, M.P.; Musumeci, D.; Montesarchio, D. Shedding light on the interaction of polydatin and resveratrol with G-quadruplex and duplex DNA: A biophysical, computational and biological approach. Int. J. Biol. Macromol. 2020, 151, 1163–1172. [Google Scholar] [CrossRef]
- Fleming, E.; Luo, Y. Co-delivery of synergistic antioxidants from food sources for the prevention of oxidative stress. J. Agric. Food Res. 2021, 3, 1–12. [Google Scholar] [CrossRef]
- Wang, L.; Sang, M.; Liu, E.; Banahene, P.O.; Zhang, Y.; Wang, T.; Han, L.; Gao, X. Rapid profiling and pharmacokinetic studies of major compounds in crude extract from Polygonum multiflorum by UHPLC-Q-TOF-MS and UPLC-MS/MS. J. Pharm. Biomed. Anal. 2017, 140, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Hou, X.; Gao, S.; Sun, M.; Lin, F.; Cai, G.; Xiao, K. Pharmacokinetics and bioavailability study of polydatin in rat plasma by using a LC–MS/MS method. Pak. J. Pharm. Sci. 2014, 27, 1931–1937. [Google Scholar]
- Irawan, C.; Hanafi; Sulistiawaty, L.; Rochaeni, H.; Lestari, P.S. Evaluation of DPPH free radical scavenging activity of Pometia pinnata from Indonesia. Pharma Innov. J. 2017, 6, 403–406. [Google Scholar]
- Saman, R.B.A.; Mokhtar, R.A.M.; Iqbal, M. Identification of Bioactive Compounds, Quantitative Measurement of Phenolics and Flavonoids Content, and Radical Scavenging Activity of Lygodium circinnatum. Trans. Sci. Technol. 2017, 4, 354–359. [Google Scholar]
- MacDonald-Wicks, L.K.; Wood, L.G.; Garg, M.L. Methodology for the determination of biological antioxidant capacityin vitro: A review. J. Sci. Food Agric. 2006, 86, 2046–2056. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C. Review: Methods Used to Evaluate the Free Radical Scavenging Activity in Foods and Biological Systems. Food Sci. Technol. Int. 2002, 8, 121–137. [Google Scholar] [CrossRef]
- Das, A.K.; Singh, V. Antioxidative free and bound phenolic constituents in botanical fractions of Indian specialty maize (Zea mays L.) genotypes. Food Chem. 2016, 201, 298–306. [Google Scholar] [CrossRef]
- Sirichaiwetchakoon, K.; Lowe, G.M.; Eumkeb, G. The Free Radical Scavenging and Anti-Isolated Human LDL Oxidation Activities of Pluchea indica (L.) Less. Tea Compared to Green Tea (Camellia sinensis). Biomed. Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Bisby, R.H.; Brooke, R.; Navaratnam, S. Effect of antioxidant oxidation potential in the oxygen radical absorption capacity (ORAC) assay. Food Chem. 2008, 108, 1002–1007. [Google Scholar] [CrossRef]
- Dudonne, S.; Vitrac, X.; Ere, P.C.; Woillez, M.; Erillon, J.-M.M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar]
- do Nascimento, K.S.; Gasparotto Sattler, J.A.; Lauer Macedo, L.F.; Serna González, C.V.; Pereira de Melo, I.L.; da Silva Araújo, E.; Granato, D.; Sattler, A.; de Almeida-Muradian, L.B. Phenolic compounds, antioxidant capacity and physicochemical properties of Brazilian Apis mellifera honeys. LWT-Food Sci. Technol. 2018, 91, 85–94. [Google Scholar] [CrossRef]
- Ganogpichayagrai, A.; Suksaard, C. Proximate composition, vitamin and mineral composition, antioxidant capacity, and anticancer activity of Acanthopanax trifoliatus. J. Adv. Pharm. Technol. Res. 2020, 11, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Li, B.; Zhang, Q.; Gao, N.; Zhang, X.; Meng, X. Effect of in vitro-simulated gastrointestinal digestion on the stability and antioxidant activity of blueberry polyphenols and their cellular antioxidant activity towards HepG2 cells. Int. J. Food Sci. Technol. 2018, 53, 61–71. [Google Scholar] [CrossRef]
- Murador, D.C.; Mercadante, A.Z.; de Rosso, V.V. Cooking techniques improve the levels of bioactive compounds and antioxidant activity in kale and red cabbage. Food Chem. 2016, 196, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Huo, J.; Huang, M.; Zhao, M.; Luo, X.; Sun, B. Structural Characterization of a Tetrapeptide from Sesame Flavor-Type Baijiu and Its Preventive Effects against AAPH-Induced Oxidative Stress in HepG2 Cells. J. Agric Food Chem. 2017, 65, 10495–10504. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.P.; Mittal, S.P.; Devasagayam, T.P.; Pal, J.K. Oxidative stress perturbs cell proliferation in human K562 cells by modulating protein synthesis and cell cycle. Free Radic Res. 2009, 43, 1090–1100. [Google Scholar] [CrossRef]
- Liao, W.; Ning, Z.; Chen, L.; Wei, Q.; Yuan, E.; Yang, J.; Ren, J. Intracellular antioxidant detoxifying effects of diosmetin on 2,2-azobis(2-amidinopropane) dihydrochloride (AAPH)-induced oxidative stress through inhibition of reactive oxygen species generation. J. Agric Food Chem. 2014, 62, 8648–8654. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Lee, S.H.; Han, J.S.; Heo, S.J.; Hwang, J.Y.; Jeon, Y.J. Protective effects of dieckol isolated from Ecklonia cava against high glucose-induced oxidative stress in human umbilical vein endothelial cells. Toxicol. In Vitro 2010, 24, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Grauzdyte, D.; Pukalskas, A.; Viranaicken, W.; El Kalamouni, C.; Venskutonis, P.R. Protective effects of Phyllanthus phillyreifolius extracts against hydrogen peroxide induced oxidative stress in HEK293 cells. PLoS ONE 2018, 13, e0207672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, E.H.; Grant, M.H. The effect of the flavonoids, quercetin, myricetin and epicatechin on the growth and enzyme activities of MCF7 human breast cancer cells. Chem.-Biol. Interact. 1998, 116, 213–228. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Rupa, P.; Matsui, T.; Tanaka, M.; Konishi, T.; Sauchi, Y.; Sato, K.; Ono, S.; Mine, Y. In vitro and ex vivo uptake of glutathione (GSH) across the intestinal epithelium and fate of oral GSH after in vivo supplementation. J. Agric. Food Chem. 2014, 62, 9499–9506. [Google Scholar] [CrossRef]
- Yang, L.; Xu, F.; Zhang, M.; Shang, X.Y.; Xie, X.; Fu, T.; Li, J.P.; Li, H.L. Role of LncRNA MALAT-1 in hypoxia-induced PC12 cell injury via regulating p38MAPK signaling pathway. Neurosci. Lett. 2018, 670, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.-Y.; Yan, X.-P.; Li, S.-X.; Zhou, J.; Cai, G.X.; Zheng, Q.Y.; Huang, D. Isolation and Identification of Oxyresveratrol in Mulberry Branch. Sci. Seric. 2011, 37, 0948–0951. [Google Scholar] [CrossRef]
- Guo, Y.X.; Bai, H.D.; Zhang, Z.Q. The Resveratrol Withdraws in the Medicine Mulberry. Sichuan Food Ferment. 2007, 43, 24–27. [Google Scholar]
- Miao, J.; Liao, W.; Kang, M.; Jia, Y.; Wang, Q.; Duan, S.; Xiao, S.; Cao, Y.; Ji, H. Anti-fatigue and anti-oxidant activities of oyster (Ostrea rivularis) hydrolysate prepared by compound protease. Food Funct. 2018, 9, 6577–6585. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Tsai, M.S.; Lu, H.Y.; Chang, C.K.; Cheng, K.C.; Jhan, M.H.; Hsieh, C.W. Enzymatic hydrolysates obtained from Trametes versicolor polysaccharopeptides protect human skin keratinocyte against AAPH-induced oxidative stress and inflammatory. J. Cosmet Dermatol. 2019, 18, 2011–2018. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Stilbenes | Mulberry Fruits (μg/g) | Mulberry Branches (μg/g) | Mulberry Roots (μg/g) |
|---|---|---|---|
| RES | 0.48 ± 0.0030 a | 5.70 ± 0.34 c | 32.45 ± 1.72 d |
| PD | 0.0020 ± 0.0028 b | 0.33 ± 0.028 a | 3.15 ± 1.52 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chen, X.; Liu, G.; Li, J.; Zhang, J.; Cao, Y.; Miao, J. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.). Molecules 2021, 26, 7574. https://doi.org/10.3390/molecules26247574
Li Z, Chen X, Liu G, Li J, Zhang J, Cao Y, Miao J. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.). Molecules. 2021; 26(24):7574. https://doi.org/10.3390/molecules26247574
Chicago/Turabian StyleLi, Ziwei, Xiaoman Chen, Guo Liu, Jun Li, Jinglin Zhang, Yong Cao, and Jianyin Miao. 2021. "Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.)" Molecules 26, no. 24: 7574. https://doi.org/10.3390/molecules26247574