
Discovery of Novel Cyclic Ethers with Synergistic Antiplasmodial Activity in Combination with Valinomycin

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
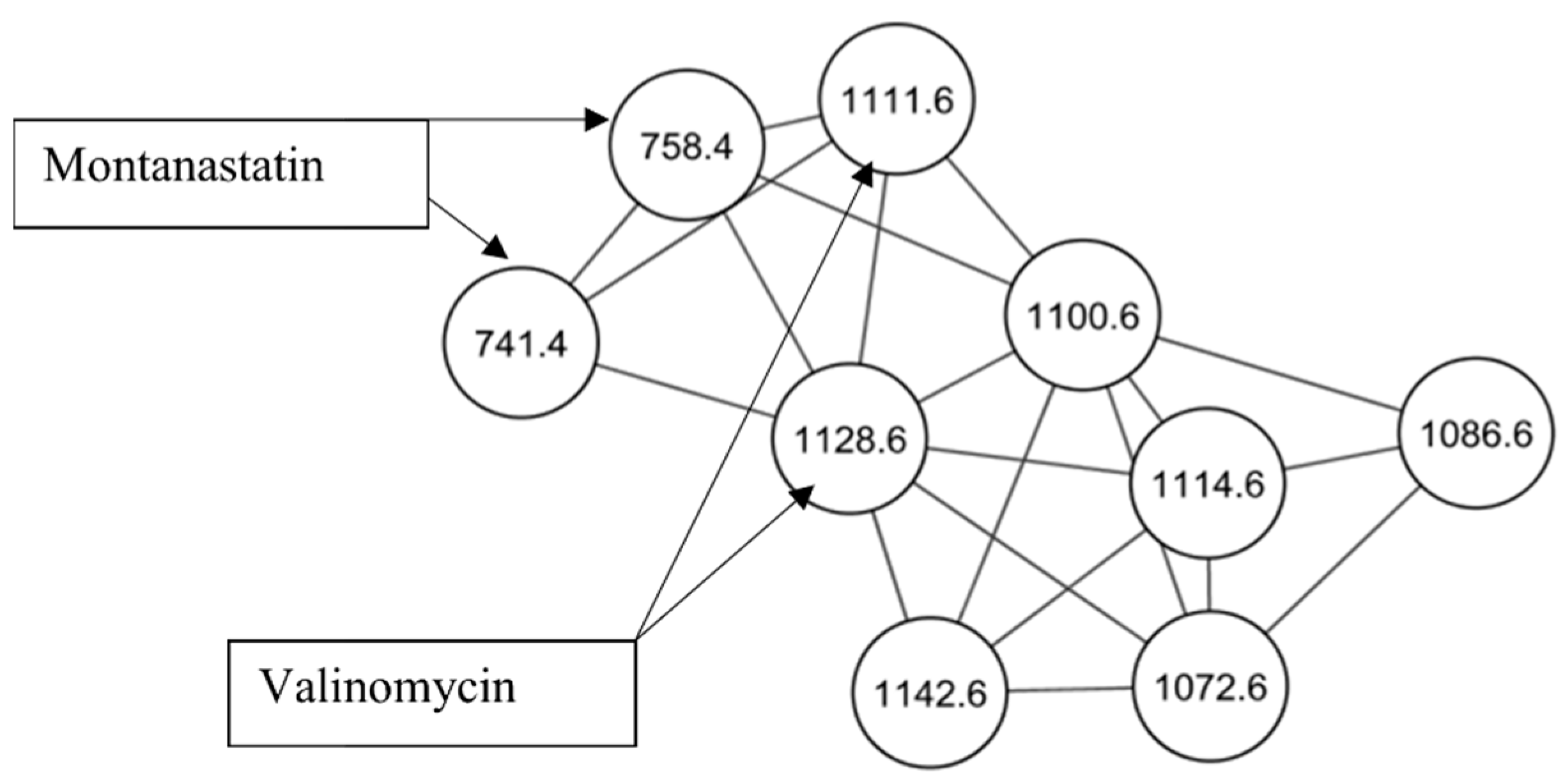
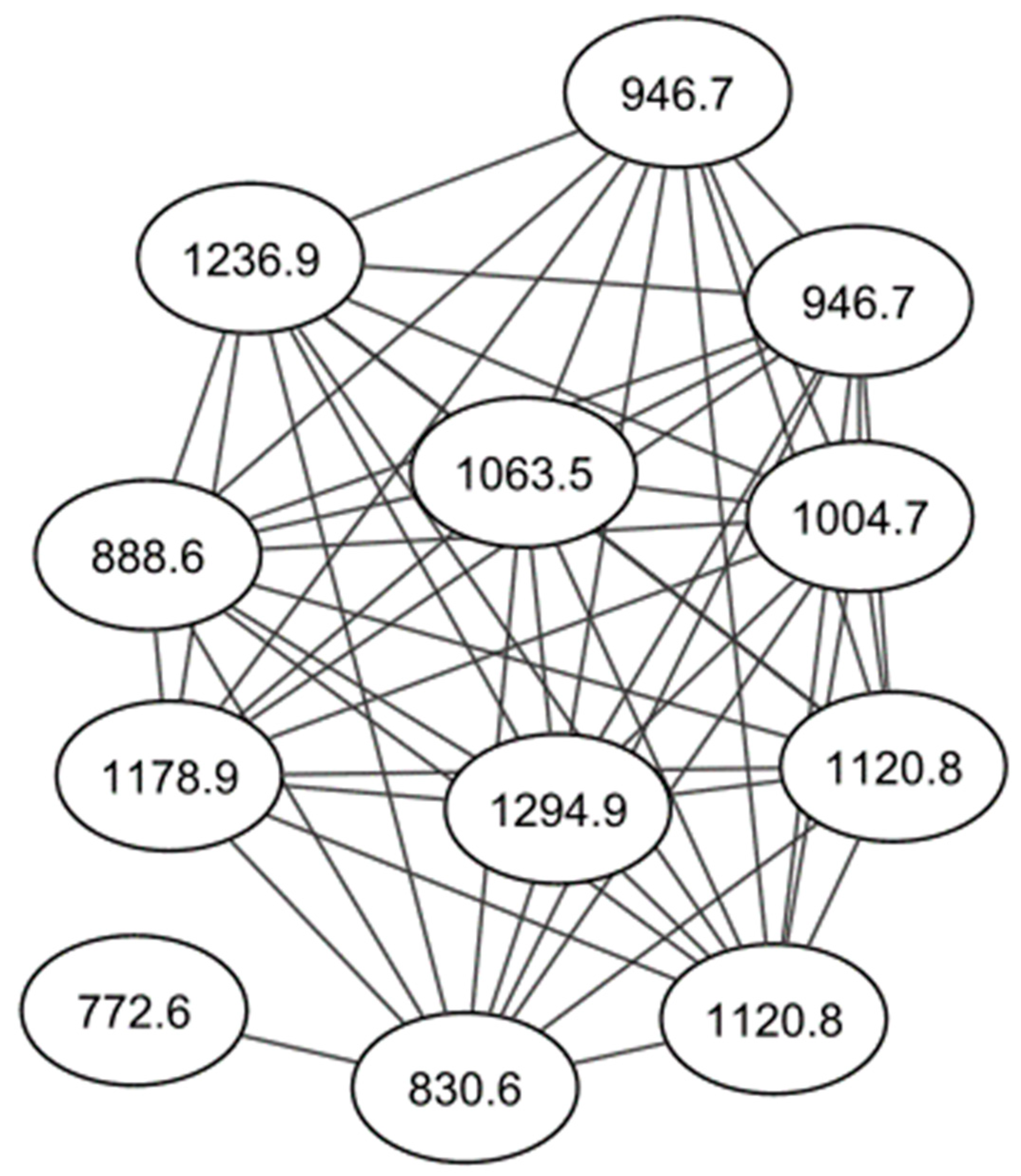
2.1. Identification of Cyclodepsipeptides
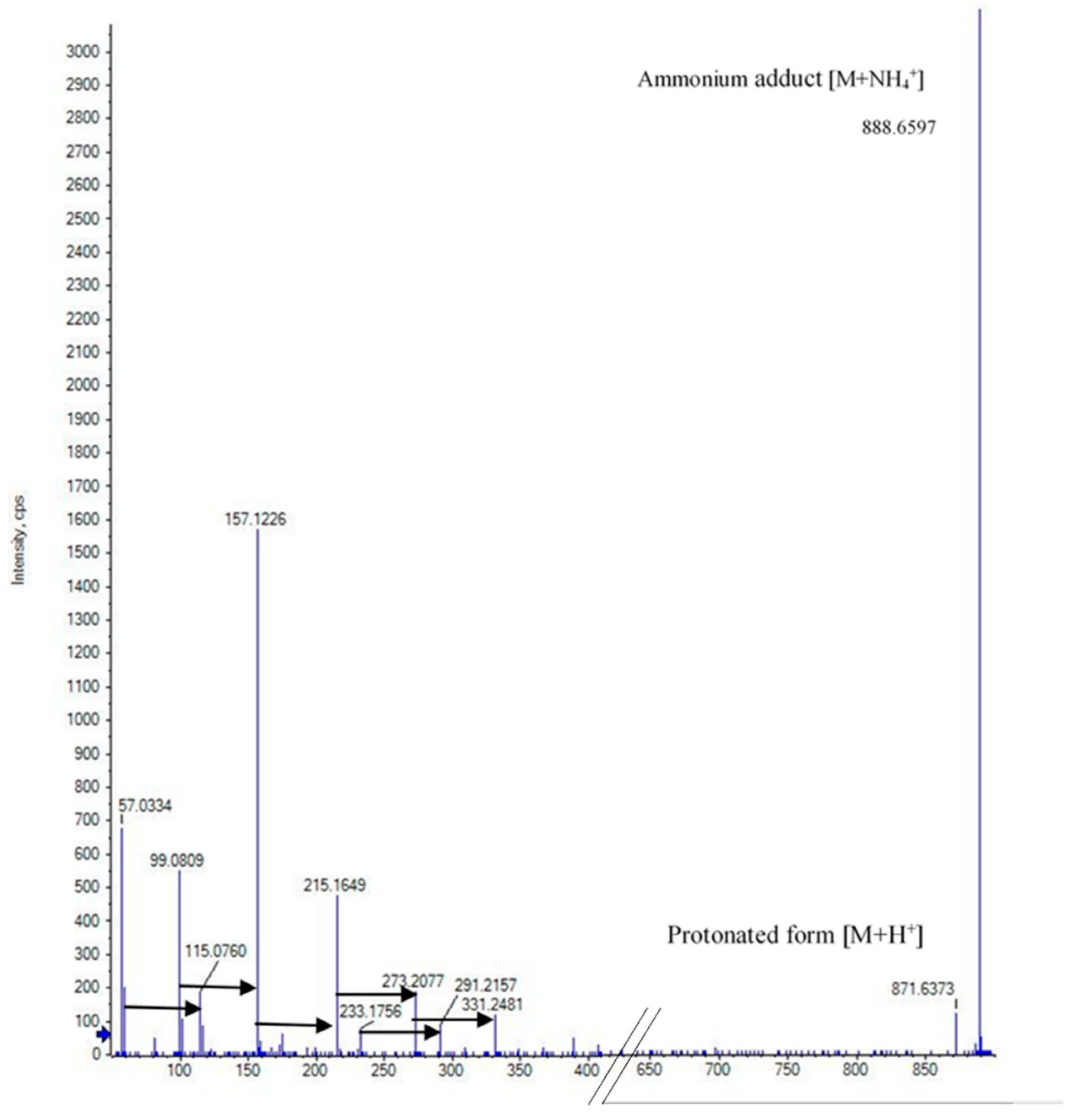
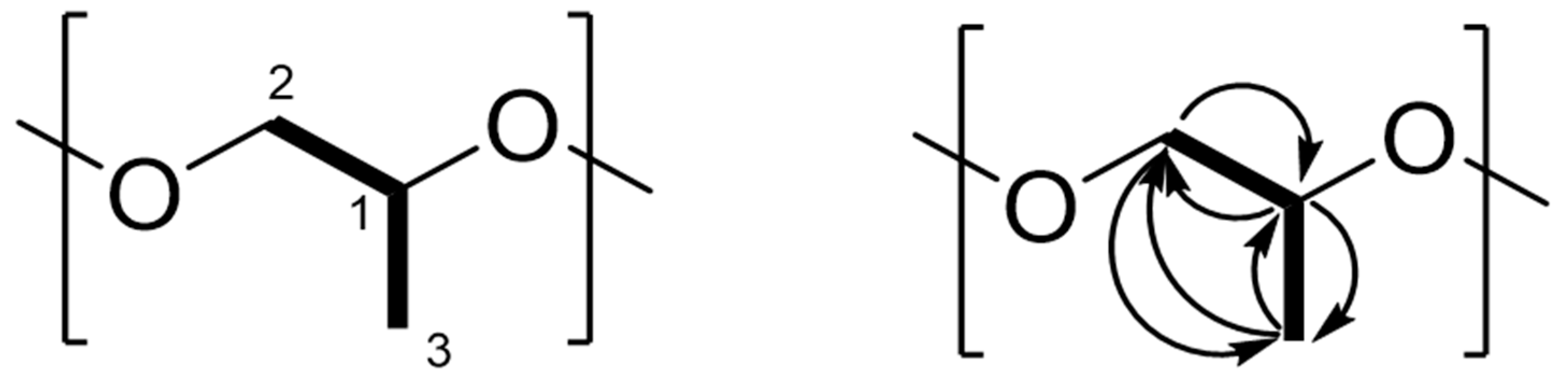
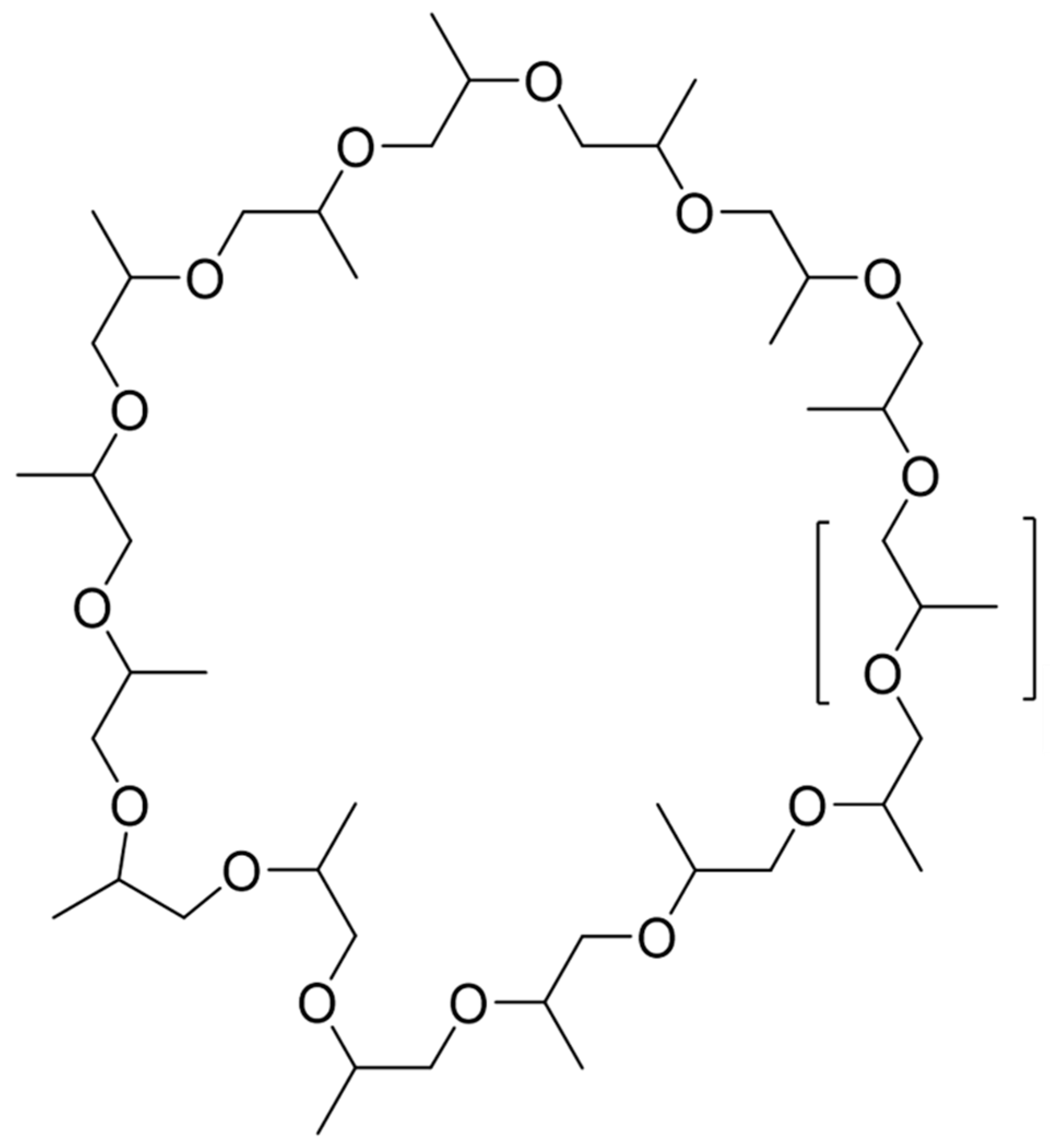
2.2. Elucidation of the Cyclic Polypropylene Glycol
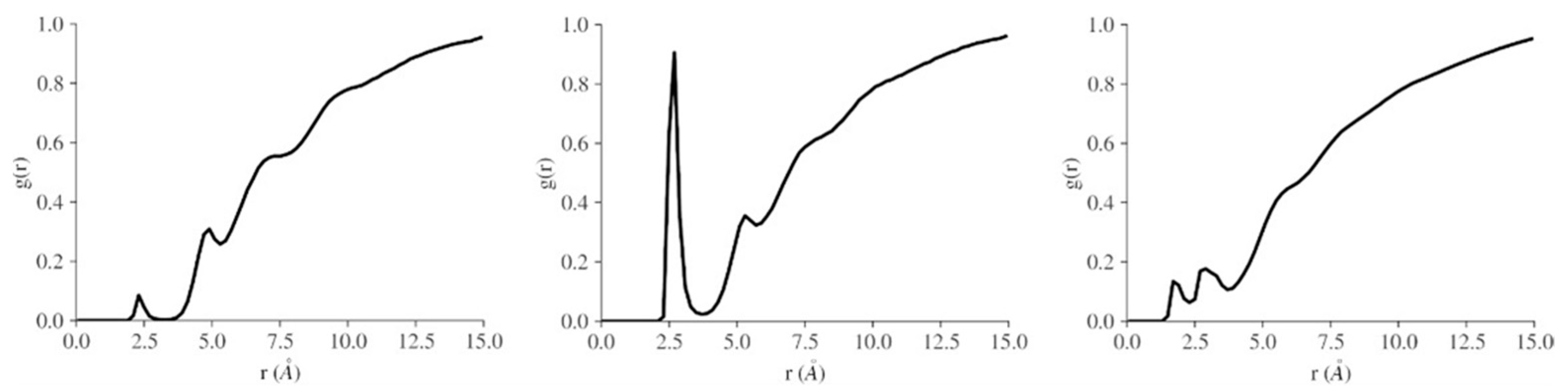
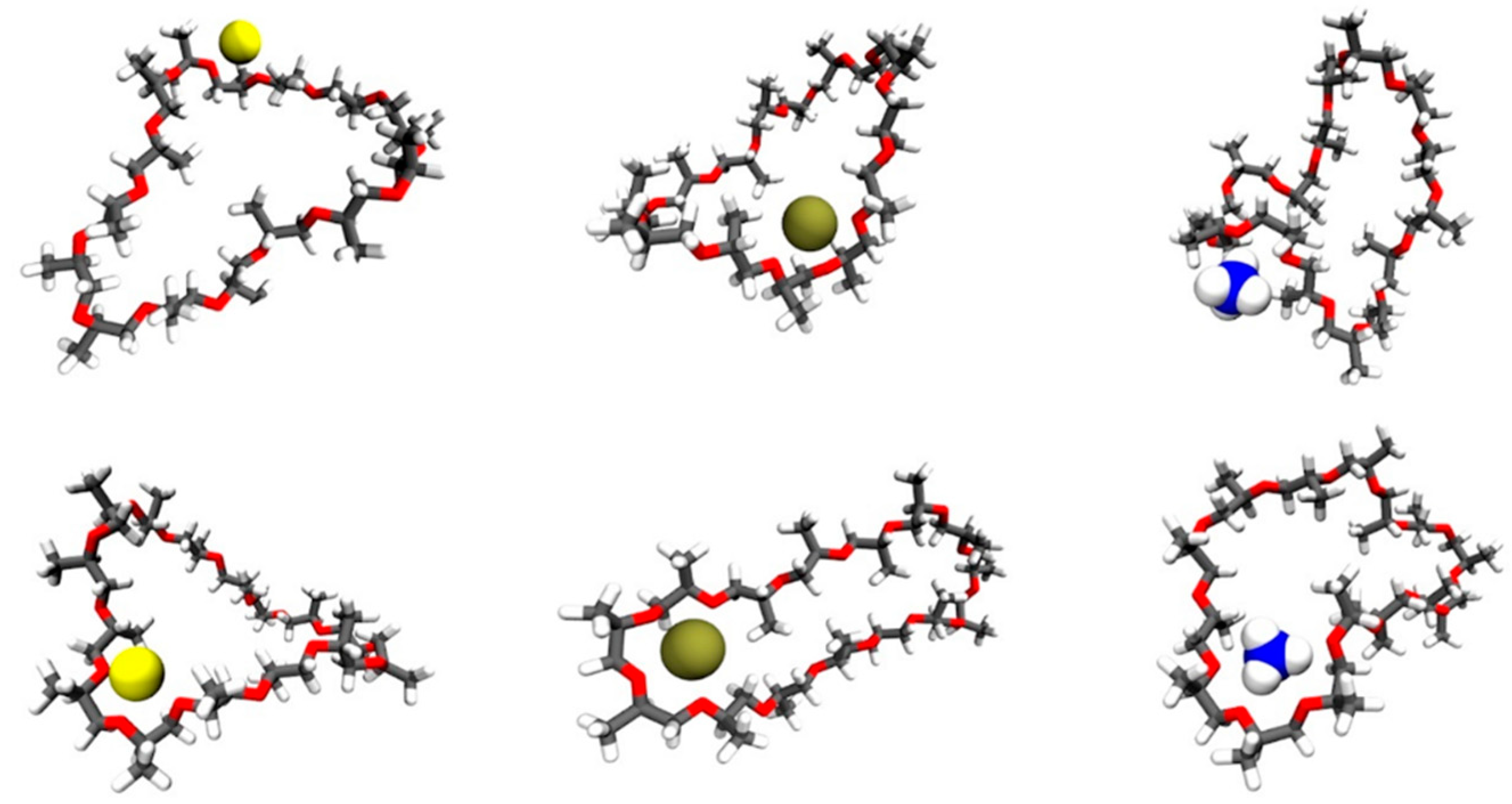
3. Computational Studies
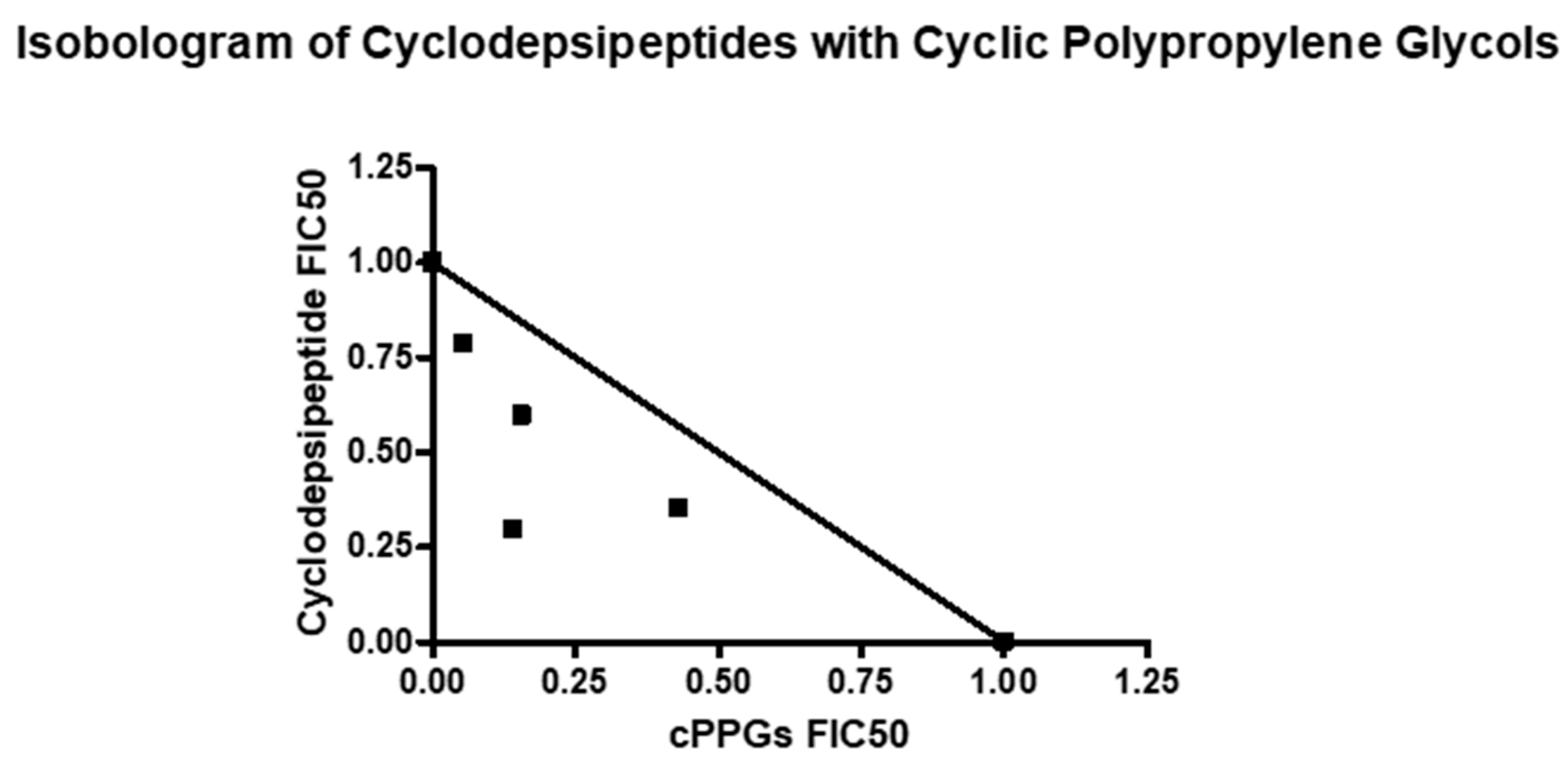
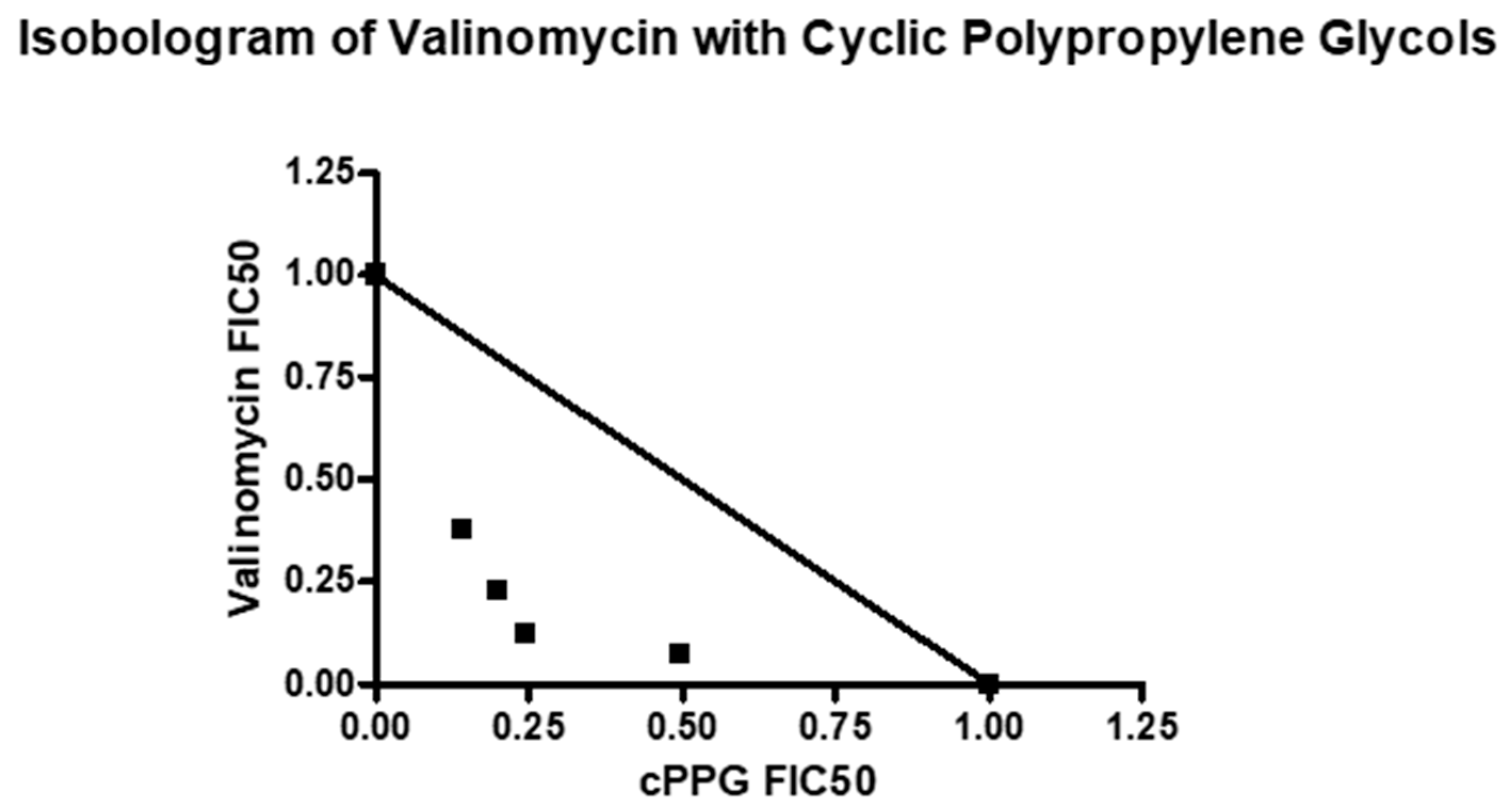
4. Combination Studies
5. Cytotoxicity Studies
6. Conclusions
7. Methods
7.1. Fermentation and Extraction
7.2. Fractionation
7.3. P. falciparum, NF54 Parasite Lactate Dehydrogenase Assay
7.4. Spectroscopic Analysis
7.5. Global Natural Products Social Molecular Networking
7.6. Molecular Dynamics Setup and Analysis
7.7. Cytotoxicity Assays
7.8. Fixed-Ratio Isobologram
8. Data Availability
8.1. Molecular Networks
8.2. Dereplicator
8.3. Varquest
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fong, C.W. Physiology of ionophore transport of potassium and sodium ions across cell membranes: Valinomycin and 18-Crown-6 Ether. Int. J. Comput. Biol. Drug Des. 2016, 9, 228–246. [Google Scholar] [CrossRef]
- Bharti, H.; Singal, A.; Raza, M.; Ghosh, P.; Nag, A. Ionophores as Potent Anti-malarials: A Miracle in the Making. Curr. Top. Med. Chem. 2019, 18, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Park, C.N.; Lee, J.M.; Lee, D.; Kim, B.S. Antifungal Activity of Valinomycin, a Peptide Antibiotic Produced by Streptomyces sp. Strain M10 Antagonistic to Botrytis Cinerea. J. Microbiol. Biotechnol. 2008, 18, 880–884. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18633285 (accessed on 7 September 2021).
- Kaushik, V.; Yakisich, J.S.; Kumar, A.; Azad, N.; Iyer, A.K.V. Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers. Cancers 2018, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Kunimoto, S.; Takahashi, Y.; Naganawa, H.; Sakaue, M.; Inoue, S.; Ohno, T.; Takeuchi, T. Inhibitory effects of polyethers on human immunodeficiency virus replication. Antimicrob. Agents Chemother. 1992, 36, 492–494. [Google Scholar] [CrossRef] [Green Version]
- Gumila, C.; Ancelin, M.L.; Delort, A.M.; Jeminet, G.; Vial, H.J. Characterization of the Potent In Vitro and In Vivo Antimalarial Activities of Ionophore Compounds. Antimicrob. Agents Chemother. 1997, 41, 523–529. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9055986 (accessed on 28 June 2021). [CrossRef] [Green Version]
- Gumila, C.; Ancelin, M.L.; Jeminet, G.; Delort, A.M.; Miquel, G.; Vial, H.J. Differential In Vitro Activities of Ionophore Compounds against Plasmodium falciparum and Mammalian Cells. Antimicrob. Agents Chemother. 1996, 40, 602–608. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8851578 (accessed on 28 June 2021). [CrossRef] [Green Version]
- Maron, M.I.; Magle, C.T.; Czesny, B.; Turturice, B.; Huang, R.; Zheng, W.; Vaidya, A.B.; Williamson, K.C. Maduramicin Rapidly Eliminates Malaria Parasites and Potentiates the Gametocytocidal Activity of the Pyrazoleamide PA21A050. Antimicrob. Agents Chemother. 2015, 60, 1492–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, A.B.; Morrisey, J.M.; Zhang, Z.; Das, S.; Daly, T.M.; Otto, T.; Spillman, N.; Wyvratt, M.; Siegl, P.; Marfurt, J.; et al. Pyrazoleamide compounds are potent antimalarials that target Na+ homeostasis in intraerythrocytic Plasmodium falciparum. Nat. Commun. 2014, 5, 5521. [Google Scholar] [CrossRef] [Green Version]
- Otoguro, K.; Kohana, A.; Manabe, C.; Ishiyama, A.; Ui, H.; Shiomi, K.; Yamada, H.; Omura, S. Potent Antimalarial Activities of Polyether Antibiotic, X-206. J. Antibiot. 2001, 54, 658–663. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. World Malaria Report 2020. 2020. Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 25 June 2021).
- Roberts, L. Drug-resistant malaria advances in Mekong. Science 2017, 358, 155–156. [Google Scholar] [CrossRef]
- Noedl, H.; Se, Y.; Schaecher, K.; Smith, B.L.; Socheat, D.; Fukuda, M.M. Evidence of artemisinin-resistant malaria in western Cambodia. N. Engl. J. Med. 2008, 359, 2619–2620. [Google Scholar] [CrossRef]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsburg, H. Transport pathways in the malaria-infected erythrocyte. Their characterization and their use as potential targets for chemotherapy. Biochem. Pharmacol. 1994, 48, 1847–1856. [Google Scholar] [CrossRef]
- Watson, D. Screening of Actinobacteria for Novel Antimalarial Compounds. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2020. [Google Scholar]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulo, B.S.; Sigrist, R.; Angolini, C.F.F.; De Oliveira, L.G. New Cyclodepsipeptide Derivatives Revealed by Genome Mining and Molecular Networking. ChemistrySelect 2019, 4, 7785–7790. [Google Scholar] [CrossRef]
- Singh, V.P.; Sharma, R.; Sharma, V.; Raina, C.; Kapoor, K.K.; Kumar, A.; Chaubey, A.; Singh, D.; Vishwakarma, R.A. Isolation of depsipeptides and optimization for enhanced production of valinomycin from the North-Western Himalayan cold desert strain Streptomyces lavendulae. J. Antibiot. 2019, 72, 617–624. [Google Scholar] [CrossRef]
- Wibowo, J.T.; Kellermann, M.Y.; Versluis, D.; Putra, M.Y.; Murniasih, T.; Mohr, K.I.; Wink, J.; Engelmann, M.; Praditya, D.F.; Steinmann, E.; et al. Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia. Mar. Drugs 2019, 17, 635. [Google Scholar] [CrossRef] [Green Version]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of microbial metabolites through database search of mass spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Mikheenko, A.; Shlemov, A.; Korobeynikov, A.; Mohimani, H.; Pevzner, P.A. Increased diversity of peptidic natural products revealed by modification-tolerant database search of mass spectra. Nat. Microbiol. 2018, 3, 319–327. [Google Scholar] [CrossRef]
- Wibowo, J.; Kellermann, M.; Köck, M.; Putra, M.; Murniasih, T.; Mohr, K.; Wink, J.; Praditya, D.; Steinmann, E.; Schupp, P. Anti-Infective and Antiviral Activity of Valinomycin and Its Analogues from a Sea Cucumber-Associated Bacterium, Streptomyces sp. SV 21. Mar. Drugs 2021, 19, 81. [Google Scholar] [CrossRef]
- Rindelaub, J.D.; Baird, Z.; Lindner, B.A.; Strantz, A.A. Identifying extractable profiles from 3D printed medical devices. PLoS ONE 2019, 14, e0217137. [Google Scholar] [CrossRef]
- Gavril, G.-L.; Wrona, M.; Bertella, A.; Świeca, M.; Râpă, M.; Salafranca, J.; Nerín, C. Influence of medicinal and aromatic plants into risk assessment of a new bioactive packaging based on polylactic acid (PLA). Food Chem. Toxicol. 2019, 132, 110662. [Google Scholar] [CrossRef] [PubMed]
- Arenaz, P.; Bitticks, L.; Pannell, K.; Garcia, S. Genotoxic potential of crown ethers in mammalian cells: Induction of sister-chromatid exchanges. Mutat. Res. Toxicol. 1992, 280, 109–115. [Google Scholar] [CrossRef]
- Hayvalı, Z.; Güler, H.; Öğütcü, H.; Sarı, N. Novel bis-crown ethers and their sodium complexes as antimicrobial agent: Synthesis and spectroscopic characterizations. Med. Chem. Res. 2014, 23, 3652–3661. [Google Scholar] [CrossRef]
- Michaud-Agrawal, N.; Denning, E.J.; Woolf, T.B.; Beckstein, O. MDAnalysis: A toolkit for the analysis of molecular dynamics simulations. J. Comput. Chem. 2011, 32, 2319–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Tubiana, T.; Carvaillo, J.-C.; Boulard, Y.; Bressanelli, S. TTClust: A Versatile Molecular Simulation Trajectory Clustering Program with Graphical Summaries. J. Chem. Inf. Model. 2018, 58, 2178–2182. [Google Scholar] [CrossRef]
- Weller, M.G. A unifying review of bioassay-guided fractionation, effect-directed analysis and related techniques. Sensors 2012, 12, 9181–9209. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adovelande, J.; Schrével, J. Carboxylic ionophores in malaria chemotherapy: The effects of monensin and nigericin on Plasmodium falciparum in vitro and Plasmodium vinckei petteri in vivo. Life Sci. 1996, 59, PL309–PL315. [Google Scholar] [CrossRef]
- Matthews, H.; Deakin, J.; Rajab, M.; Idris-Usman, M.; Nirmalan, N.J. Investigating antimalarial drug interactions of emetine dihydrochloride hydrate using CalcuSyn-based interactivity calculations. PLoS ONE 2017, 12, e0173303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fivelman, Q.L.; Adagu, I.S.; Warhurst, D.C. Modified Fixed-Ratio Isobologram Method for Studying In Vitro Interactions between Atovaquone and Proguanil or Dihydroartemisinin against Drug-Resistant Strains of Plasmodium falciparum. Antimicrob. Agents Chemother. 2004, 48, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, S.; Viljoen, A. Plant-Based Antimicrobial Studies–Methods and Approaches to Study the Interaction between Natural Products. Planta Med. 2011, 77, 1168–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-Y.; Jan, J.-T.; Ma, S.-H.; Kuo, C.-J.; Juan, H.-F.; Cheng, Y.-S.E.; Hsu, H.-H.; Huang, H.-C.; Wu, D.; Brik, A.; et al. Small molecules targeting severe acute respiratory syndrome human coronavirus. Proc. Natl. Acad. Sci. USA 2004, 101, 10012–10017. [Google Scholar] [CrossRef] [Green Version]
- Pink, R.; Hudson, A.; Mouriès, M.-A.; Bendig, M. Opportunities and Challenges in Antiparasitic Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 727–741. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Yuan, G.; Wang, Y.; Qu, Y.; Zhou, M. Synergistic combination of two antimicrobial agents closing each other’s mutant selection windows to prevent antimicrobial resistance. Sci. Rep. 2018, 8, 7237. [Google Scholar] [CrossRef]
- Ara, I.; Kudo, T. Sphaerosporangium gen. nov., a new member of the family Streptosporangiaceae, with descriptions of three new species as Sphaerosporangium melleum sp. nov., Sphaerosporangium rubeum sp. nov. and Sphaerosporangium cinnabarinum sp. nov., and transfer of Streptosporangium viridialbum Nonomura and Ohara 1960 to Sphaerosporangium viridialbum comb. nov. Actinomycetologica 2007, 21, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Makler, M.T.; Ries, J.M.; Williams, J.A.; Bancroft, J.E.; Piper, R.C.; Gibbins, B.L.; Hinrichs, D.J. Parasite lactate dehydrogenase as an assay for Plasmodium falciparum drug sensitivity. Am. J. Trop. Med. Hyg. 1993, 48, 739–741. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Case, P.A.K.D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cisneros, G.A.; Cruzeiro, V.W.D.; Darden, T.A.; et al. Amber Tools; San Fran: 2021. Available online: https://www.researchgate.net/publication/37460688_AMBER_10_University_of_California_San_Francisco (accessed on 9 December 2021).
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Eastman, P.; Swails, J.; Chodera, J.D.; McGibbon, R.T.; Zhao, Y.; Beauchamp, K.A.; Wang, L.-P.; Simmonett, A.C.; Harrigan, M.P.; Stern, C.D.; et al. OpenMM 7: Rapid development of high performance algorithms for molecular dynamics. PLoS Comput. Biol. 2017, 13, e1005659. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Antiplasmodial Activity against P. falciparum, NF54 IC50 (ng/mL) |
|---|---|
| Crude Extract | 497 ± 14 |
| Reverse Phase SPE Fractions | |
| #1 Water | >5000 |
| #2 Water/MeOH (1:1) | >5000 |
| #3 MeOH | 40.0 ± 1.5 |
| Normal Phase SPE Fractions | |
| #4 Hexane | >5000 |
| #5 Hexane/EtAc (8:1) | 150.0 ± 53 |
| #6 Hexane/EtAc (7:1) | 31.3 ± 4.2 |
| #7 Hexane/EtAc (6:1) | 63.5 ± 17.0 |
| #8 Hexane/EtAc (5:1) | 287.0 ± 14.0 |
| #9 Hexane/EtAc (4:1) | >5000 |
| #10 Hexane/EtAc (3:1) | >5000 |
| #11 Hexane/EtAc (2:1) | >5000 |
| #12 Hexane/EtAc (1:1) | >5000 |
| #13 EtAc | 2105.0 ± 146.0 |
| #14 EtAc/MeOH (4:1) | >5000 |
| #15 EtAc/MeOH (2:1) | >5000 |
| #16 MeOH | >5000 |
| Positive Controls | |
| Chloroquine | 7.1 ± 2.1 |
| Artemisinin | 3.3 ± 1.3 |
| Position | 13C | 1H mult | 13C-1H COSY | 13C-1H HMBC |
|---|---|---|---|---|
| 1 | 75.3, CH | 3.63 m | 2,3 | 2,3 |
| 2 | 72.87, CH2 | 3.55 3.48 m | 1 | 1,3 |
| 3 | 16.22, CH3 | 1.16 m | 2 | 1,2 |
| Sample | Cytotoxicity against CHO, IC50 (µg/mL) | Antiplasmodial Activity against P. falciparum, NF54 IC50 (ng/mL) | Selectivity Index (SI) * |
|---|---|---|---|
| Valinomycin | 12.40 ± 1.1 | 3.75 ± 0.77 | 3306 |
| Fixed Ratio 4:1 | 60.80 ± 6.3 | 1.86 ± 0.05 | 32,688 |
| Fixed Ratio 3:2 | 57.88 ± 6.1 | 0.90 ± 0.20 | 64,311 |
| Fixed Ratio 2:3 | 65.25 ± 1.3 | 0.75 ± 0.08 | 87,000 |
| Fixed Ratio 1:4 | 58.43 ± 6.1 | 0.53 ± 0.1 | 110,245 |
| Cyclic Polpropylene Glycols (Fraction #13) | >100 ± ND | 1792 ± 547 | >55 |
| Emetine | 0.03 ± 0.002 | ND | ND |
| Ratio Number | Fixed Ratio | Cyclodepsipeptides Concentration (µg/mL) | Cyclic Polypropylene Glycols Concentration (µg/mL) |
|---|---|---|---|
| 1 | 5:0 | 1.6 | 0 |
| 2 | 4:1 | 1.28 | 8 |
| 3 | 3:2 | 0.96 | 16 |
| 4 | 2:3 | 0.60 | 24 |
| 5 | 1:4 | 0.32 | 32 |
| 6 | 0:5 | 0 | 40 |
| Ratio Number | Fixed Ratio | Valinomycin Concentration (ng/mL) | Cyclic Polypropylene Glycols Concentration (µg/mL) |
|---|---|---|---|
| 1 | 5:0 | 160 | 0 |
| 2 | 4:1 | 128 | 8 |
| 3 | 3:2 | 96 | 16 |
| 4 | 2:3 | 64 | 24 |
| 5 | 1:4 | 32 | 32 |
| 6 | 0:5 | 0 | 40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watson, D.J.; Meyers, P.R.; Acquah, K.S.; Dziwornu, G.A.; Barnett, C.B.; Wiesner, L. Discovery of Novel Cyclic Ethers with Synergistic Antiplasmodial Activity in Combination with Valinomycin. Molecules 2021, 26, 7494. https://doi.org/10.3390/molecules26247494
Watson DJ, Meyers PR, Acquah KS, Dziwornu GA, Barnett CB, Wiesner L. Discovery of Novel Cyclic Ethers with Synergistic Antiplasmodial Activity in Combination with Valinomycin. Molecules. 2021; 26(24):7494. https://doi.org/10.3390/molecules26247494
Chicago/Turabian StyleWatson, Daniel J., Paul R. Meyers, Kojo Sekyi Acquah, Godwin A. Dziwornu, Christopher Bevan Barnett, and Lubbe Wiesner. 2021. "Discovery of Novel Cyclic Ethers with Synergistic Antiplasmodial Activity in Combination with Valinomycin" Molecules 26, no. 24: 7494. https://doi.org/10.3390/molecules26247494