
Antibacterial Activities of Prenylated Isoflavones from Maclura tricuspidata against Fish Pathogenic Streptococcus: Their Structure-Activity Relationships and Extraction Optimization

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Plant Materials
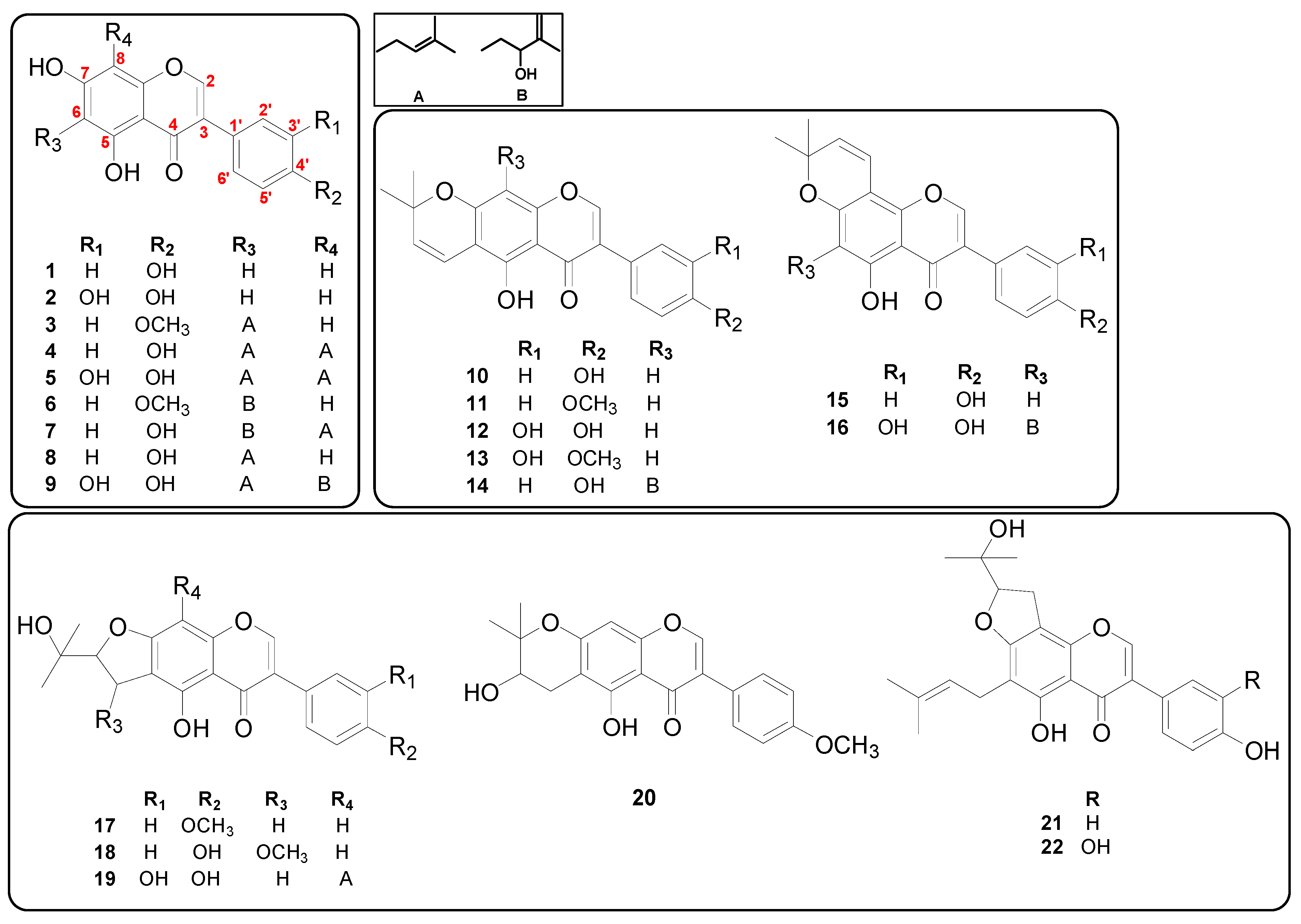
2.3. Extraction, Isolation, and Identification of Isoflavones from M. tricuspidata
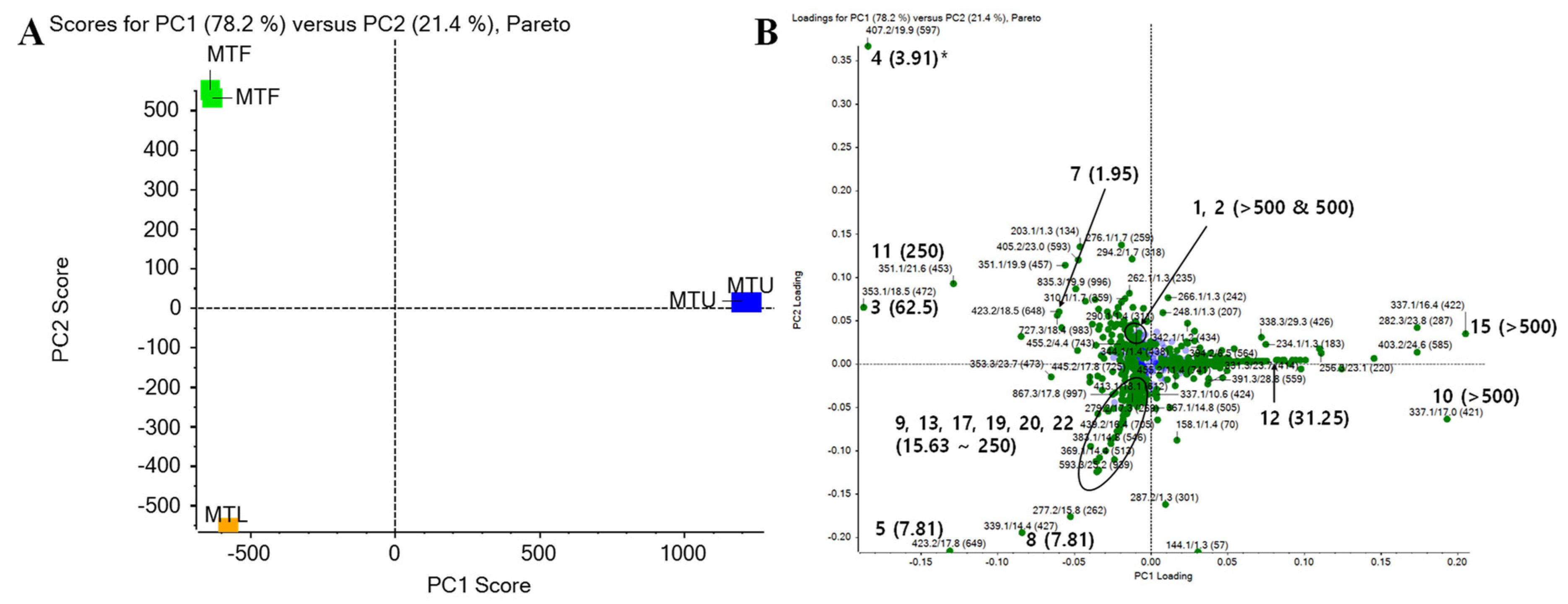
2.4. LC-Q-TOF MS Analysis of Isoflavones in Extracts of M. tricuspidata Leaves, Ripe Fruits, and Unripe Fruits with Principal Component Analyses (PCA)
2.5. Bacteria and Culture Conditions
2.6. Antibacterial Susceptibility Test
2.7. Time-Growth Curve and Scanning Electron Microscope (SEM) Analyses
2.8. Cytotoxicity Assay
2.9. Box-Behnken Design (BBD)
2.10. Quantification of 6,8-Diprenylgenistein (4) in OE-MTF and 15 Extracts from BBD
Method Validation
2.11. Statistical Analysis
3. Results and Discussion
3.1. Antibacterial Activities of Isoflavones from M. tricuspidata against S. iniae and Their Structure–Activity Relationships (SAR)
3.1.1. Effects of Prenyl Group—Addition
3.1.2. Effect of Cyclization of Prenyl Group
3.1.3. Effects of Other Substituents
3.2. Antibacterial Activities of Prenylated Isoflavones against Fish Pathogenic Clinical Strains of Streptococcus
3.3. Antibacterial Activities of Extracts from Each Part of M. tricuspidata against Fish Pathogenic Bacteria and Principal Component Analysis (PCA) Using LC-Q-TOF MS
3.4. Optimization Procedures
3.4.1. Statistical Analyses and Model Fitting of BBD
3.4.2. Multiple Responses Optimization of MTF Extracts and Predictive Capacities of Mathematical Models
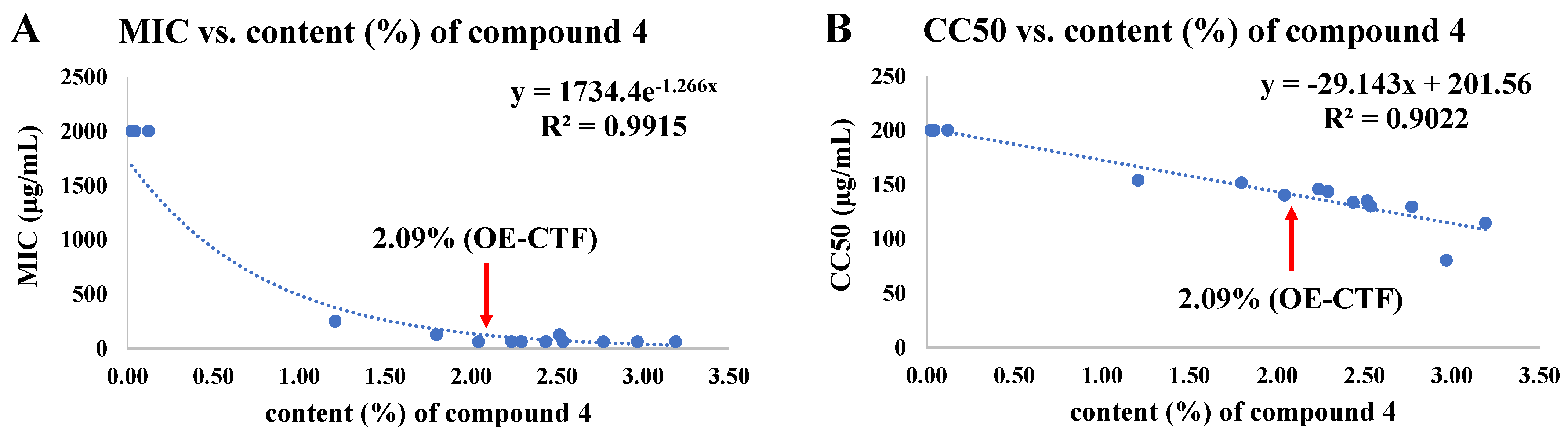
3.5. Correlations among 6,8-Diprenylgenistein (4) Content, Antibacterial Activity, and Cytotoxicity According to Extraction Conditions Based on BBD
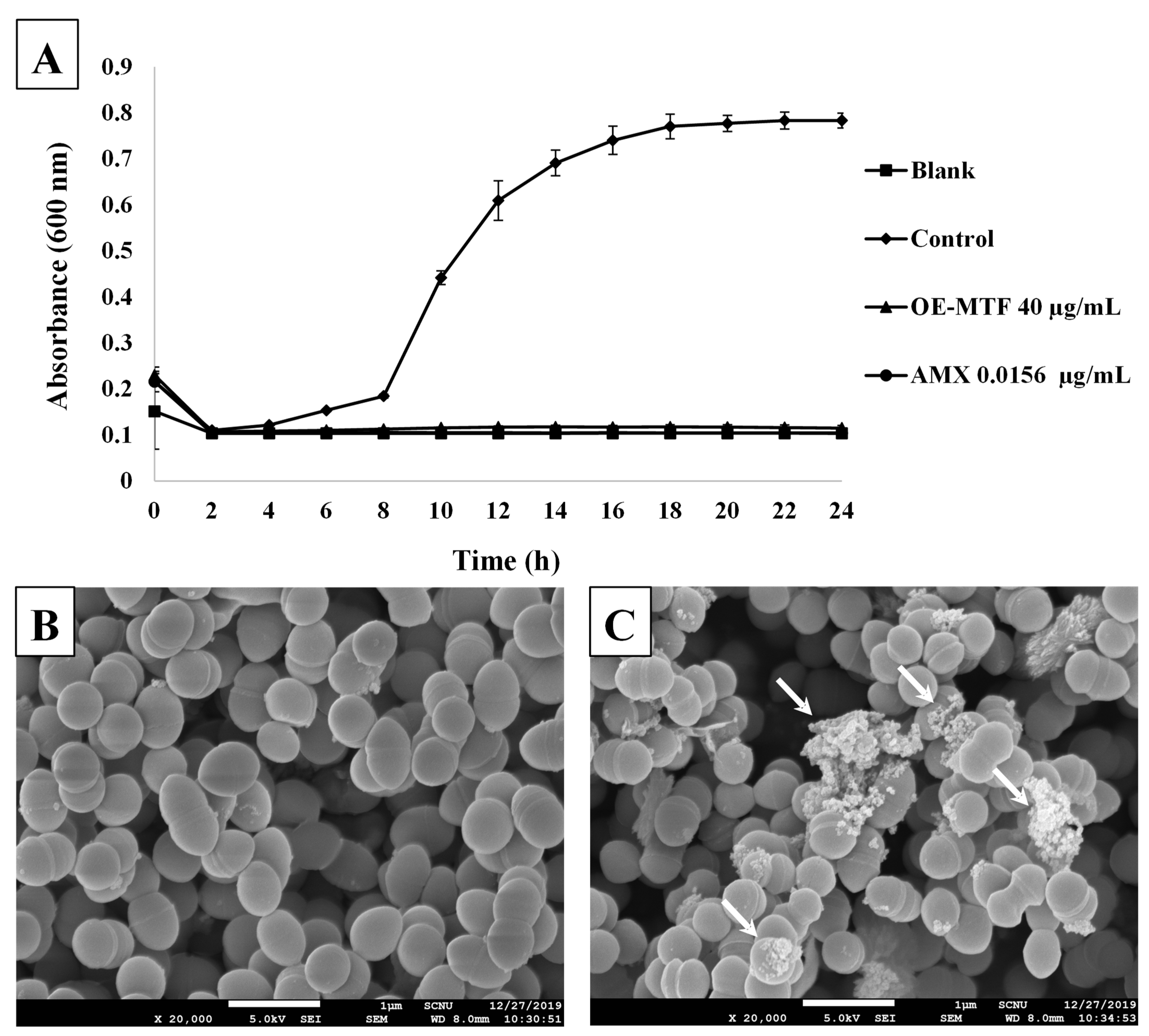
3.6. Effects of OE-MTF on the Growth and the Morphology of S. iniae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
List of Abbreviations
| ANOVA | Analysis of variance |
| BBD | Box–Behnken design |
| BHIA | Brain heart infusion agar |
| BHIB | Brain heart infusion broth |
| CC50 | 50% cytotoxic concentration |
| DCM | Dichloromethane |
| DMSO | Dimethyl sulfoxide |
| EtOAc | Ethyl acetate |
| FBS | Fetal bovine serum |
| FHM | Fathead minnow cells |
| 1H-NMR | 1H-nuclear magnetic resonance |
| HPLC | High-performance liquid chromatography |
| IDA | Information-dependent acquisition |
| LC-Q-TOF MS | Liquid chromatography-quadrupole-time of flight mass spectrometry |
| LOD | Limit of detection |
| LOQ | Limit of quantification |
| MBC | Minimum bactericidal concentration |
| MeOH | Methanol |
| MIC | Minimum inhibitory concentration |
| MRM | Multiple reaction monitoring |
| MTF | Fresh ripe fruits of M. tricuspidata |
| MTL | Dried leaves of M. tricuspidata |
| MTU | Fresh unripe fruits of M. tricuspidata |
| n-BuOH | n-Butanol |
| NP-MPLC | Silica gel column chromatography |
| NR | Neutral red |
| OE-MTF | Optimized extract from MTF |
| PBS | Phosphate-buffered saline |
| PCA | Principal component analysis |
| RP-MPLC | Reverse phase column chromatography |
| RSD | Relative standard deviation |
| SAR | Structure–activity relationships |
| SEM | Scanning electron microscope |
| UV | Ultraviolet |
References
- FAO. Part 1. World review. In The State of World Fisheries and Aquaculture 2020; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2020; p. 2. [Google Scholar] [CrossRef]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Jee, B.Y.; Shin, K.W.; Lee, D.W.; Kim, Y.J.; Lee, M.K. Monitoring of the mortalities and medications in the inland farms of olive flounder, Paralichthys olivaceus, in South Korea. J. Fish Pathol. 2014, 27, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Lee, H.N.; Jee, B.Y.; Woo, S.H.; Kim, Y.J.; Lee, M.K. Monitoring of the mortalities in the aquaculture farms of South Korea. J. Fish Pathol. 2012, 25, 271–277. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, E.; He, Y.; Wang, K.; Yang, Q.; Wang, J.; Geng, Y.; Chen, D.; Huang, X.; Ouyang, P. Identification and screening of effective protective antigens for channel catfish against Streptococcus iniae. Oncotarget 2017, 8, 30793. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, M.R.; Litt, M.; Kertesz, D.A.; Wyper, P.; Rose, D.; Coulter, M.; McGeer, A.; Facklam, R.; Ostach, C.; Willey, B.M. Invasive infections due to a fish pathogen, Streptococcus iniae. N. Engl. J. Med. 1997, 337, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Björklund, H.; Bondestam, J.; Bylund, G. Residues of oxytetracycline in wild fish and sediments from fish farms. Aquaculture 1990, 86, 359–367. [Google Scholar] [CrossRef]
- Kemper, N. Veterinary antibiotics in the aquatic and terrestrial environment. Ecol. Indic. 2008, 8, 1–13. [Google Scholar] [CrossRef]
- Chakraborty, S.B.; Hancz, C. Application of phytochemicals as immunostimulant, antipathogenic and antistress agents in finfish culture. Rev. Aquac. 2011, 3, 103–119. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Sarter, S.; Sasal, P.; Caruso, D. Moving towards more sustainable aquaculture practices: A meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. 2021, 13, 537–555. [Google Scholar] [CrossRef]
- Jo, Y.H.; Kim, S.B.; Liu, Q.; Do, S.-G.; Hwang, B.Y.; Lee, M.K. Comparison of pancreatic lipase inhibitory isoflavonoids from unripe and ripe fruits of Cudrania tricuspidata. PLoS ONE 2017, 12, e0172069. [Google Scholar]
- Xin, L.-T.; Yue, S.-J.; Fan, Y.-C.; Wu, J.-S.; Yan, D.; Guan, H.-S.; Wang, C.-Y. Cudrania tricuspidata: An updated review on ethnomedicine, phytochemistry and pharmacology. RSC Adv. 2017, 7, 31807–31832. [Google Scholar] [CrossRef] [Green Version]
- Park, B.-H.; Back, K.-Y.; Lee, S.-I.; Kim, S.-D. Quality and antioxidative characteristics of Cudrania tricuspidata leaves tea. Korean J. Food Preserv. 2008, 15, 461–468. [Google Scholar]
- Jeong, J.Y.; Jo, Y.H.; Lee, K.Y.; Do, S.-G.; Hwang, B.Y.; Lee, M.K. Optimization of pancreatic lipase inhibition by Cudrania tricuspidata fruits using response surface methodology. Bioorg. Med. Chem. Lett. 2014, 24, 2329–2333. [Google Scholar] [CrossRef]
- Jo, Y.H.; Choi, K.-M.; Liu, Q.; Kim, S.B.; Ji, H.-J.; Kim, M.; Shin, S.-K.; Do, S.-G.; Shin, E.; Jung, G. Anti-obesity effect of 6,8-diprenylgenistein, an isoflavonoid of Cudrania tricuspidata fruits in high-fat diet-induced obese mice. Nutrients 2015, 7, 10480–10490. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.J.; Kang, J.H.; Lee, C.H.; Song, K.B. Application of Cudrania tricuspidata leaf extract as a washing agent to inactivate Listeria monocytogenes on fresh-cut romaine lettuce and kale. Int. J. Food Sci. 2020, 55, 276–282. [Google Scholar] [CrossRef]
- Jo, Y.H.; Lee, S.; Yeon, S.W.; Turk, A.; Lee, J.H.; Hong, S.-M.; Han, Y.K.; Lee, K.Y.; Hwang, B.Y.; Kim, S.Y. Anti-diabetic potential of Masclura tricuspidata leaves: Prenylated isoflavonoids with α-glucosidase inhibitory and anti-glycation activity. Bioorg. Chem. 2021, 114, 105098. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; den Besten, H.M.; Aisyah, S.; Gruppen, H.; Vincken, J.-P. The position of prenylation of isoflavonoids and stilbenoids from legumes (Fabaceae) modulates the antimicrobial activity against Gram positive pathogens. Food Chem. 2017, 226, 193–201. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; Vincken, J.-P.; van de Schans, M.G.; Hageman, J.; Schaftenaar, G.; den Besten, H.M.; Gruppen, H. QSAR-based molecular signatures of prenylated (iso) flavonoids underlying antimicrobial potency against and membrane-disruption in Gram positive and Gram negative bacteria. Sci. Rep. 2018, 8, 9267. [Google Scholar] [CrossRef]
- Jang, J.Y.; Shin, H.; Lim, J.-W.; Ahn, J.H.; Jo, Y.H.; Lee, K.Y.; Hwang, B.Y.; Jung, S.-J.; Kang, S.Y.; Lee, M.K. Comparison of antibacterial activity and phenolic constituents of bark, lignum, leaves and fruit of Rhus verniciflua. PLoS ONE 2018, 13, e0200257. [Google Scholar] [CrossRef]
- Bizjak, J.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R. Changes in primary metabolites and polyphenols in the peel of “Braeburn” apples (Malus domestica Borkh.) during advanced maturation. J. Agric. Food Chem. 2013, 61, 10283–10292. [Google Scholar] [CrossRef]
- Karacabey, E.; Mazza, G. Optimisation of antioxidant activity of grape cane extracts using response surface methodology. Food Chem. 2010, 119, 343–348. [Google Scholar] [CrossRef]
- Liu, F.F.; Ang, C.Y.; Springer, D. Optimization of extraction conditions for active components in Hypericum perforatum using response surface methodology. J. Agric. Food Chem. 2000, 48, 3364–3371. [Google Scholar] [CrossRef]
- Uslu, M.E.; Erdoğan, İ.; Bayraktar, O.; Ateş, M. Optimization of extraction conditions for active components in Equisetum arvense extract. Rom. Biotechnol. Lett. 2013, 18, 8115–8131. [Google Scholar]
- CLSI. 8. Broth Dilution Procedures (Macrodilution and Microdilution). In Methods for Broth Dilution Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Approved Guideline M49-A; Clinical and Laboratory Standards Institute (CLSI): Wayne, NJ, USA, 2006; Volume 30, pp. 10–11. [Google Scholar]
- Lim, J.W.; Jung, M.H.; Jung, S.J.; Kim, D.H.; Park, K.; Kang, S. The efficacy of amoxicillin sodium against streptococcosis in cultured olive flounder Paralichthys olivaceus and its pharmacokinetics. J. Vet. Pharmacol. Ther. 2017, 40, 77–87. [Google Scholar] [CrossRef]
- Lim, J.-W.; Shin, S.-M.; Jung, S.-J.; Lee, M.K.; Kang, S.Y. Optimization of antibacterial extract from lacquer tree (Rhus verniciflua Stokes) using response surface methodology and its efficacy in controlling edwardsiellosis of olive flounder (Paralichthys olivaceus). Aquaculture 2019, 502, 40–47. [Google Scholar] [CrossRef]
- Wald-Dickler, N.; Holtom, P.; Spellberg, B. Busting the myth of “static vs cidal”: A systemic literature review. Clin. Infect. Dis. 2018, 66, 1470–1474. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Kim, J.D.; Choi, H.C.; Lee, C.W. Antibacterial activity of CNT-Ag and GO-Ag nanocomposites against gram-negative and gram-positive bacteria. Bull. Korean Chem. Soc. 2013, 34, 3261–3264. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.D. Antiviral activity of Viracea® against acyclovir susceptible and acyclovir resistant strains of herpes simplex virus. Antivir. Res. 1998, 39, 55–61. [Google Scholar] [CrossRef]
- Kang, S.Y.; Kang, J.-Y.; Oh, M.-J. Antiviral activities of flavonoids isolated from the bark of Rhus verniciflua stokes against fish pathogenic viruses In Vitro. J. Microbiol. 2012, 50, 293–300. [Google Scholar] [CrossRef]
- Lim, J.W.; Seo, J.K.; Jung, S.J.; Kang, S.Y. Efficacy of an optimized extract from licorice roots (Glycyrrhiza uralensis fischer) against viral hemorrhagic septicemia virus in olive flounder (Paralichthys olivaceus). Aquac. Res. 2021, 52, 2609–2621. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure–activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef]
- Kuete, V.; Fozing, D.; Kapche, W.; Mbaveng, A.; Kuiate, J.; Ngadjui, B.; Abegaz, B. Antimicrobial activity of the methanolic extract and compounds from Morus mesozygia stem bark. J. Ethnopharmacol. 2009, 124, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Curtis-Long, M.J.; Yuk, H.J.; Kim, D.W.; Tan, X.F.; Park, K.H. Bacterial neuraminidase inhibitory effects of prenylated isoflavones from roots of Flemingia philippinensis. Bioorg. Med. Chem. 2013, 21, 6398–6404. [Google Scholar] [CrossRef] [PubMed]
- Dhayakaran, R.P.A.; Neethirajan, S.; Xue, J.; Shi, J. Characterization of antimicrobial efficacy of soy isoflavones against pathogenic biofilms. LWT—Food Sci. Technol. 2015, 63, 859–865. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Oliveira, T.B.; Khumalo, G.P.; van Vuuren, S.F.; van Wyk, B.-E. Antimicrobial Isoflavones and Derivatives from Erythrina (Fabaceae): Structure Activity Perspective (Sar & Qsar) on Experimental and Mined Values Against Staphylococcus aureus. Antibiotics 2020, 9, 223. [Google Scholar]
- Ahmadi, A.; Gandomi, H.; Derakhshandeh, A.; Misaghi, A.; Noori, N. Phytochemical composition and in vitro safety evaluation of Ziziphora clinopodioides Lam. ethanolic extract: Cytotoxicity, genotoxicity and mutagenicity assessment. J. Ethnopharmacol. 2021, 266, 113428. [Google Scholar] [CrossRef]
- Ho, C.-T. Phenolic Compounds in Food: An overview. In Phenolic Compounds in Food and Their Effects on Health; Huang, M.-T., Ho, C.-T., Lee, C.Y., Eds.; American Chemical Society: Washington, DC, USA, 1992; Volume 506, pp. 2–7. [Google Scholar]
- Kim, M.-H.; Kim, M.-C.; Park, J.-S.; Kim, J.-W.; Lee, J.-O. The antioxidative effects of the water-soluble extracts of plants used as tea materials. Korean J. Food Sci. Technol. 2001, 33, 12–18. [Google Scholar]
- Cacace, J.; Mazza, G. Optimization of extraction of anthocyanins from black currants with aqueous ethanol. J. Food Sci. 2003, 68, 240–248. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Chemical Formula (M.W. a) | Retention Time (min) | Detected Parts from LC-Q-TOF MS b | S.iniae KCTC3657 |
|---|---|---|---|---|
| µg/mL | ||||
| Genistein (1) | C15H10O5 (270.2) | 7.73 | L, F, U | >500 |
| Orobol (2) | C15H10O6 (286.2) | 5.78 | L, F, U | 500 |
| Gancaonin A (3) | C21H20O5 (352.4) | 18.36 | L, F | 62.5 c (500 d) |
| 6,8-Diprenylgenistein (4) | C25H26O5 (406.5) | 19.83 | L, F | 3.91 (7.81) |
| 6,8-Diprenylorobol (5) | C25H26O6 (422.5) | 17.78 | L, F, U | 7.81 (31.25) |
| 5,7-Dihydroxy-6-(2″-hydroxy-3″-methylbut-3″-enyl)-4′-methoxylisoflavone (6) | C21H20O6 (368.4) | - | Not detected | 31.25 (125) |
| Isoerysenegalensein E (7) | C25H26O6 (422.5) | 18.45 | L, F | 1.95 (3.91) |
| Wighteone (8) | C20H18O5 (338.4) | 14.42 | L, F | 7.81 (15.63) |
| Millewanin H (9) | C25H26O7 (438.5) | 15.92 | L | 15.63 (62.5) |
| Alpinumisoflavone (10) | C20H16O5 (336.3) | 16.96 | F, U | >500 |
| 4′-O-Methylalpinumisoflavone (11) | C21H18O5 (350.4) | 21.50 | L, F, U | 250 (>500) |
| 5,3′,4′-Trihydroxy-6″,6″-dimethylpyrano-[2″,3″:7,6]isoflavone (12) | C20H16O6 (352.3) | 14.67 | L, U | 31.25 (250) |
| 3′-Hydroxy-4′-O-methylalpinumisoflavone (13) | C21H18O6 (366.4) | 17.44 | U | 250 (500) |
| Euchrenone b8 (14) | C25H24O6 (420.5) | - | Not detected | 62.5 (250) |
| Derrone (15) | C20H16O5 (336.3) | 16.36 | U | >500 |
| 5, 3′,4′, 2‴-Tetrahydroxy-2″, 2″-dimethylpyrano-(5″,6″:7,8)-6-(3‴-methyl-3‴-butenyl)isoflavone (16) | C25H24O7 (436.5) | - | Not detected | 62.5 (250) |
| 4’-O-Methylerythrinin C (17) | C21H20O6 (368.4) | 14.44 | L | 125 (500) |
| (±)-1″-O-Methylerythrinin F (18) | C21H20O7 (384.4) | - | Not detected | 250 (>500) |
| Furowanin A (19) | C25H26O7 (438.5) | 14.34 | L, F | 62.5 (250) |
| 4′-O-Methyl-2″-hydroxydihydroalpinumisoflavone (20) | C21H20O6 (368.4) | 15.43 | L | 125 (500) |
| Senegalensin (21) | C25H26O6 (422.5) | - | Not detected | 7.81 (31.25) |
| Furowanin B (22) | C25H26O7 (438.5) | 14.87 | L, F | 31.25 (125) |
| OTC | 0.25 (1) | |||
| AMX | 0.0078 (0.031) | |||
| Cell No. (CFU/mL) | 7.2 × 105 |
| Compounds | S. parauberis KSP44 | S. parauberis KCTC3651 | S. iniae DSJ19 | S. iniae BS9 | S. iniae KCTC3657 |
|---|---|---|---|---|---|
| µg/mL | µg/mL | µg/mL | µg/mL | µg/mL | |
| Genistein (1) | 250 a (>250 b) | >500 | >500 | >500 | >500 |
| 6,8-Diprenylgenistein (4) | 1.95 (1.95) | 3.91 (15.63) | 3.91 (7.81) | 3.91 (7.81) | 3.91 (7.81) |
| 6,8-Diprenylorobol (5) | 31.25 (125) | 31.25 (250) | 7.81 (31.25) | 7.81 (31.25) | 7.81 (31.25) |
| Isoerysenegalensein E (7) | 1.95 (1.95) | 1.95 (15.63) | 1.95 (3.91) | 1.95 (3.91) | 1.95 (3.91) |
| Wighteone (8) | 15.63 (31.25) | 15.63 (62.5) | 3.91 (15.63) | 3.91 (15.63) | 7.81 (15.63) |
| Senegalensin (21) | 15.63 (62.5) | 62.5 (250) | 7.81 (15.63) | 7.81 (15.63) | 7.81 (31.25) |
| AMX | 0.5 (4) | 0.5 (2) | 0.0156 (0.031) | 0.0078 (0.031) | 0.0078 (0.031) |
| Cell No. (CFU/mL) | 5.6 × 105 | 1.2 × 105 | 5.5 × 105 | 4.9 × 105 | 7.2 × 105 |
| Sample | S. iniae KCTC3657 | S. parauberis KCTC3651 | E. tarda KCTC12267 | A. salmonicida KCCM40239 |
|---|---|---|---|---|
| µg/mL | µg/mL | µg/mL | µg/mL | |
| MTL | 250 | 1000 | 2000 | 1000 |
| MTF | 62.5 | 250 | >2000 | 2000 |
| MTU | >2000 | >2000 | >2000 | 2000 |
| OTC | 0.25 | 0.5 | 0.5 | 0.125 |
| Cell No. (CFU/mL) | 11.6 × 105 | 11.9 × 105 | 6.5 × 105 | 7.7 × 105 |
| Response | Optimized Condition | Composite Desirability (D) | Actual Values | Predicted Values a | Predictive Capacity (%) | ||
|---|---|---|---|---|---|---|---|
| Ethanol % (X1) | Temperature (X2) | Time (X3) | |||||
| Cytotoxicity (CC50, µg/mL) | 50 | 80 | 7.5 | 0.92 | 153.18 ± 2.93 | 140.61 | 108.9% |
| Antibacterial activity (µg/mL) | 40.00 ± 0.00 b (80.00 c) | 43.40 | 108.5% | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.-W.; Jo, Y.H.; Choi, J.-S.; Lee, M.K.; Lee, K.Y.; Kang, S.Y. Antibacterial Activities of Prenylated Isoflavones from Maclura tricuspidata against Fish Pathogenic Streptococcus: Their Structure-Activity Relationships and Extraction Optimization. Molecules 2021, 26, 7451. https://doi.org/10.3390/molecules26247451
Lim J-W, Jo YH, Choi J-S, Lee MK, Lee KY, Kang SY. Antibacterial Activities of Prenylated Isoflavones from Maclura tricuspidata against Fish Pathogenic Streptococcus: Their Structure-Activity Relationships and Extraction Optimization. Molecules. 2021; 26(24):7451. https://doi.org/10.3390/molecules26247451
Chicago/Turabian StyleLim, Jae-Woong, Yang Hee Jo, Ji-Seok Choi, Mi Kyeong Lee, Ki Yong Lee, and So Young Kang. 2021. "Antibacterial Activities of Prenylated Isoflavones from Maclura tricuspidata against Fish Pathogenic Streptococcus: Their Structure-Activity Relationships and Extraction Optimization" Molecules 26, no. 24: 7451. https://doi.org/10.3390/molecules26247451