
Caragana rosea Turcz Methanol Extract Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Suppressing the TLR4/NF-κB/IRF3 Signaling Pathways

 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Results
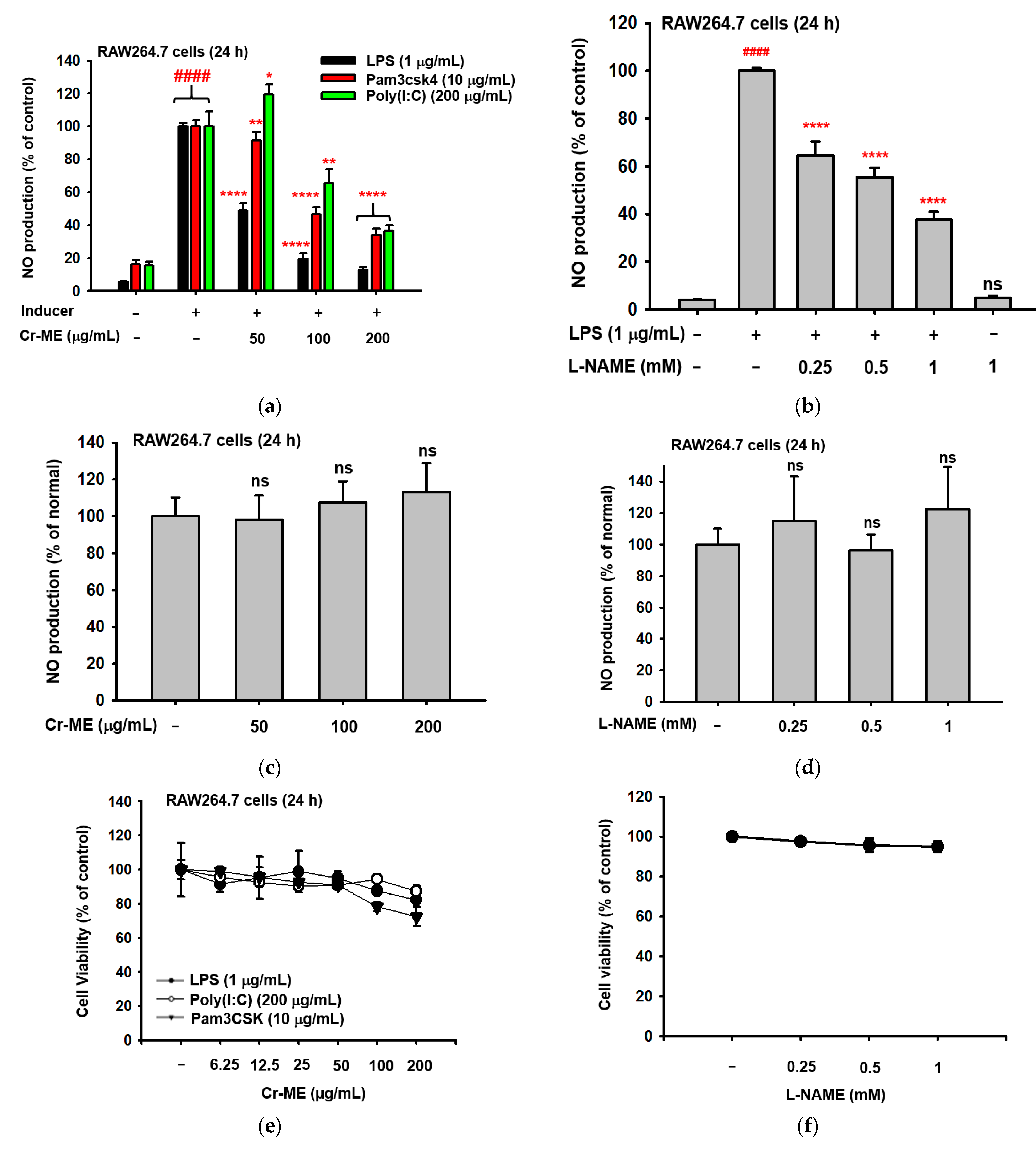
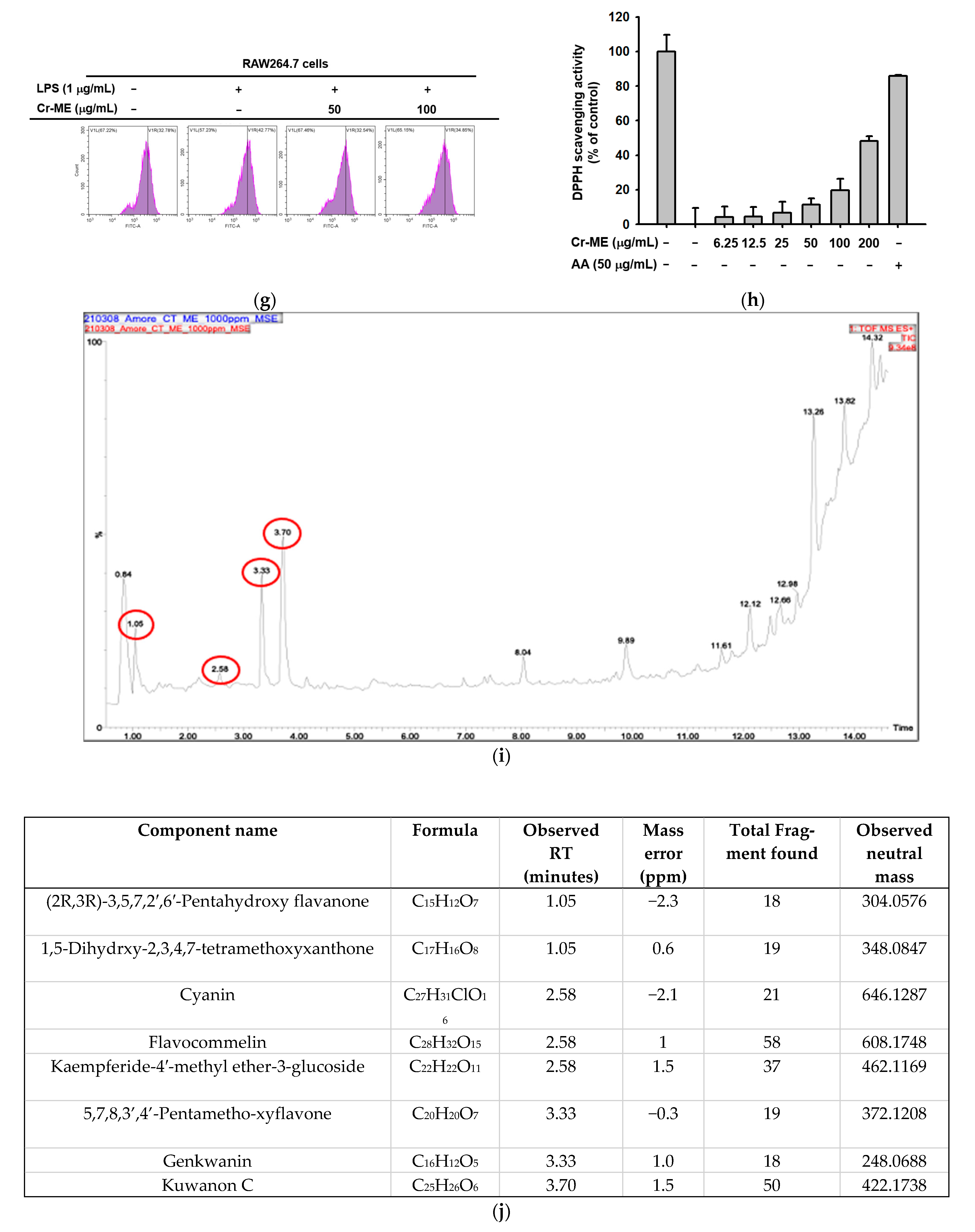
2.1. Effects of Cr-ME on Nitric Oxide (NO) Level and Cell Viability in LPS-Treated RAW264.7 Macrophages
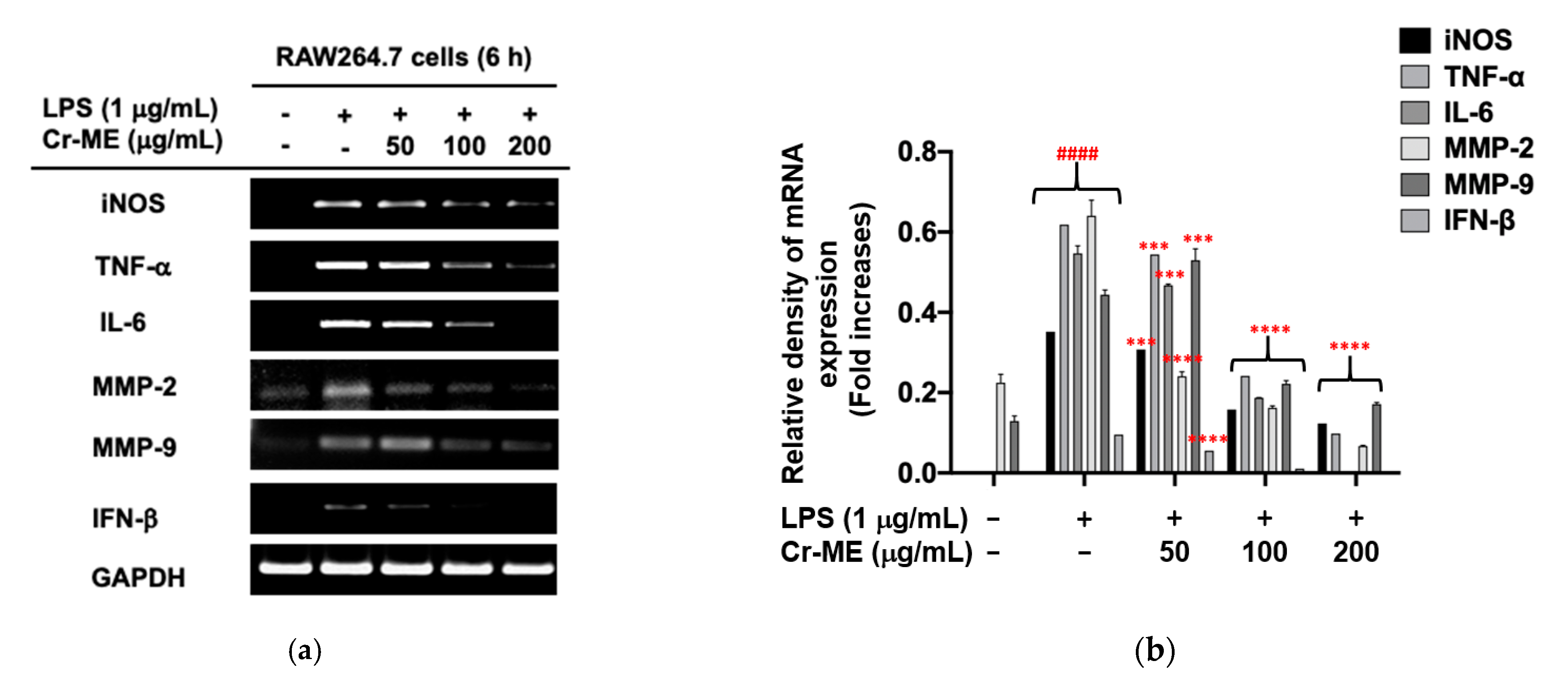
2.2. Effects of Cr-ME on Inflammatory Gene Expression at the Transcriptional Level
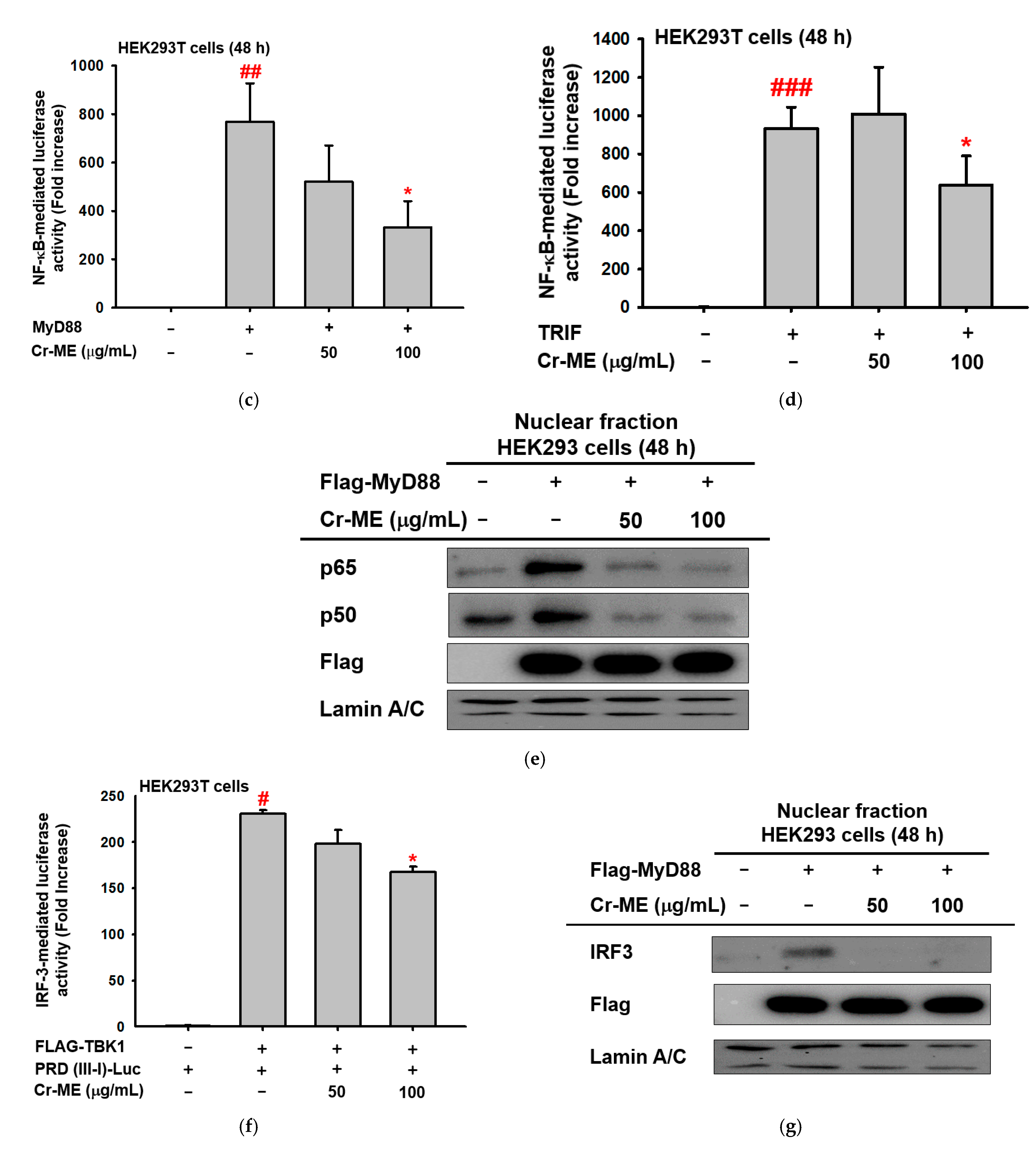
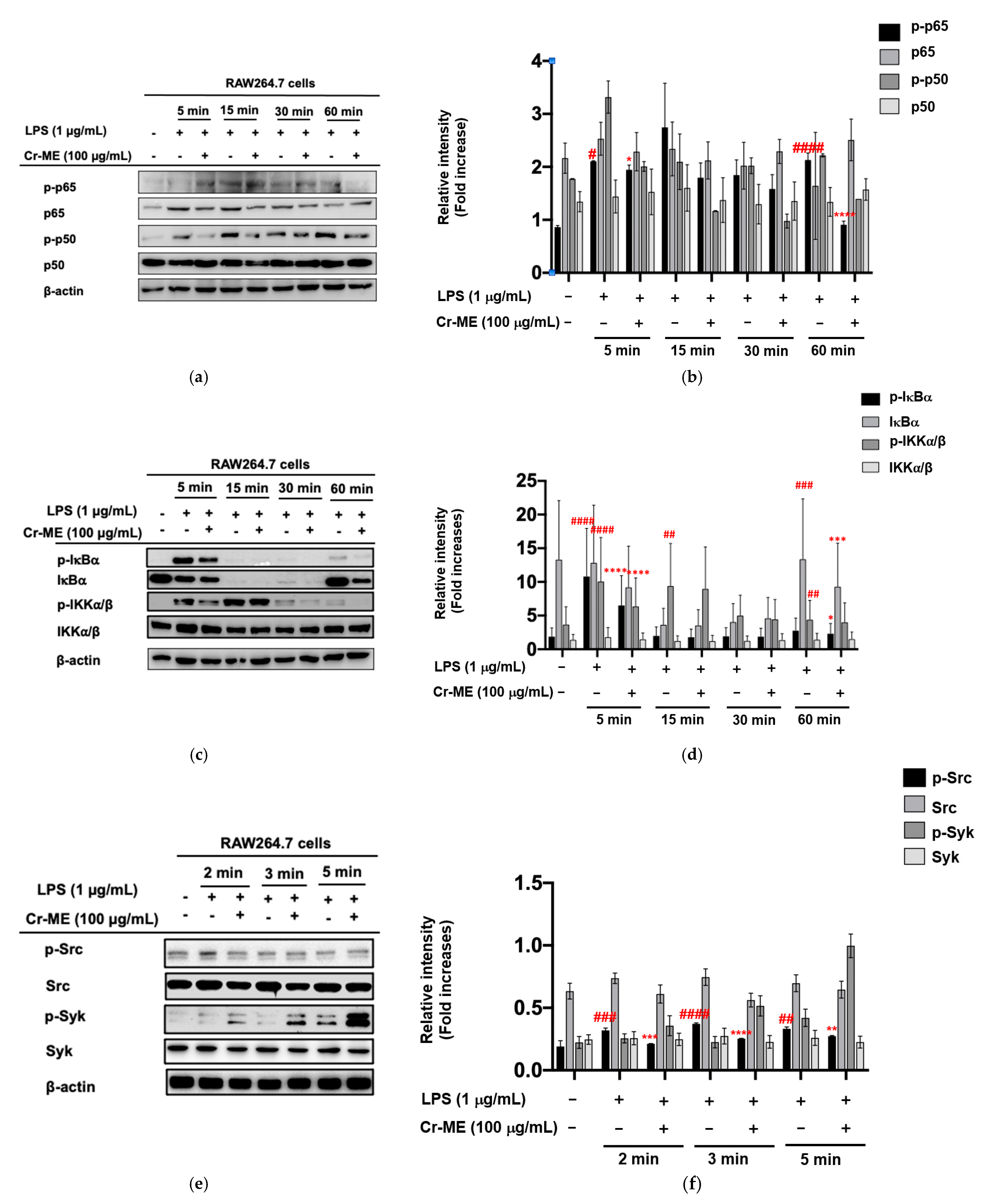
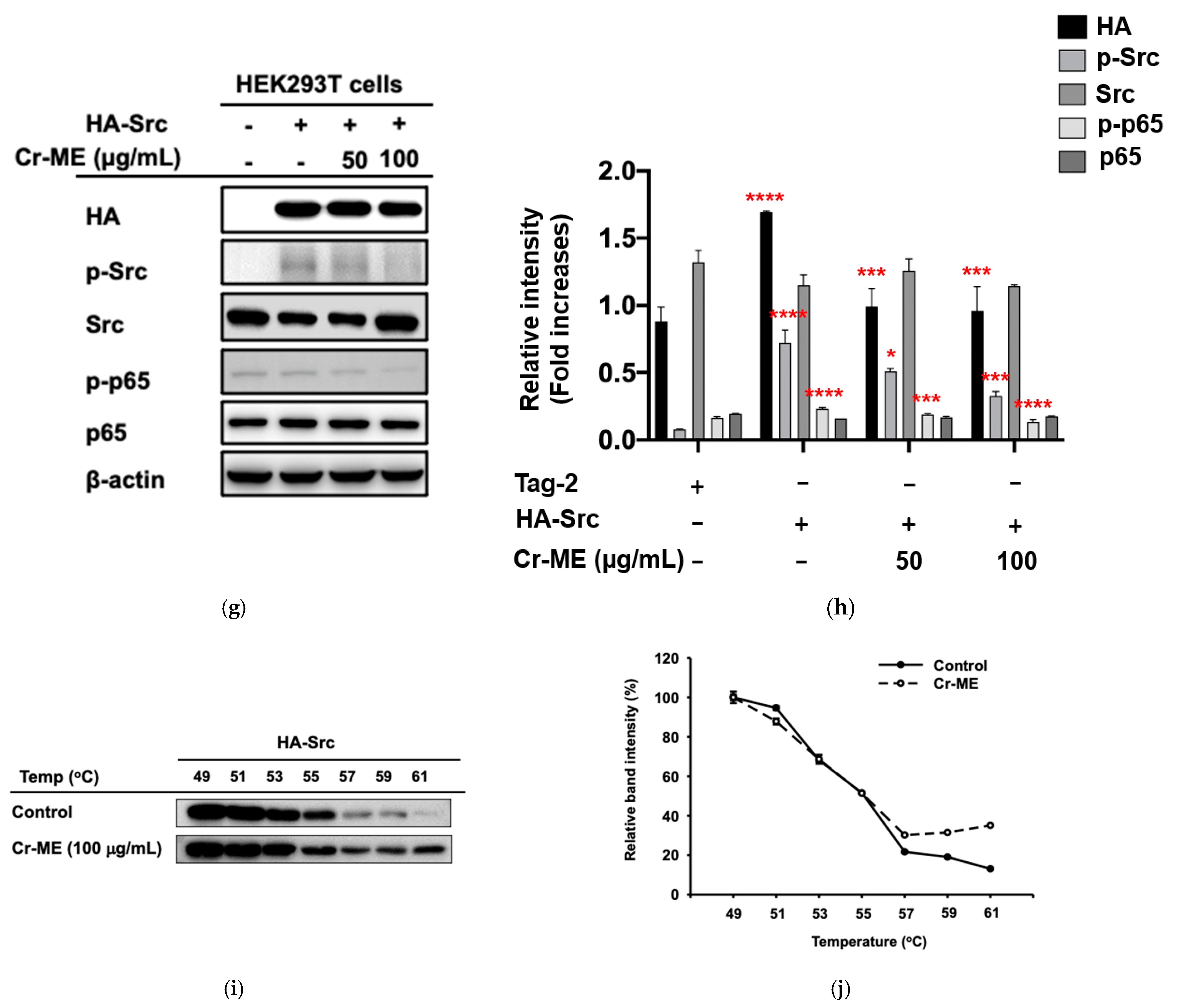
2.3. Effects of Cr-ME on LPS-Induced NF-κB Signaling and Its Upstream Enzyme Src Activity
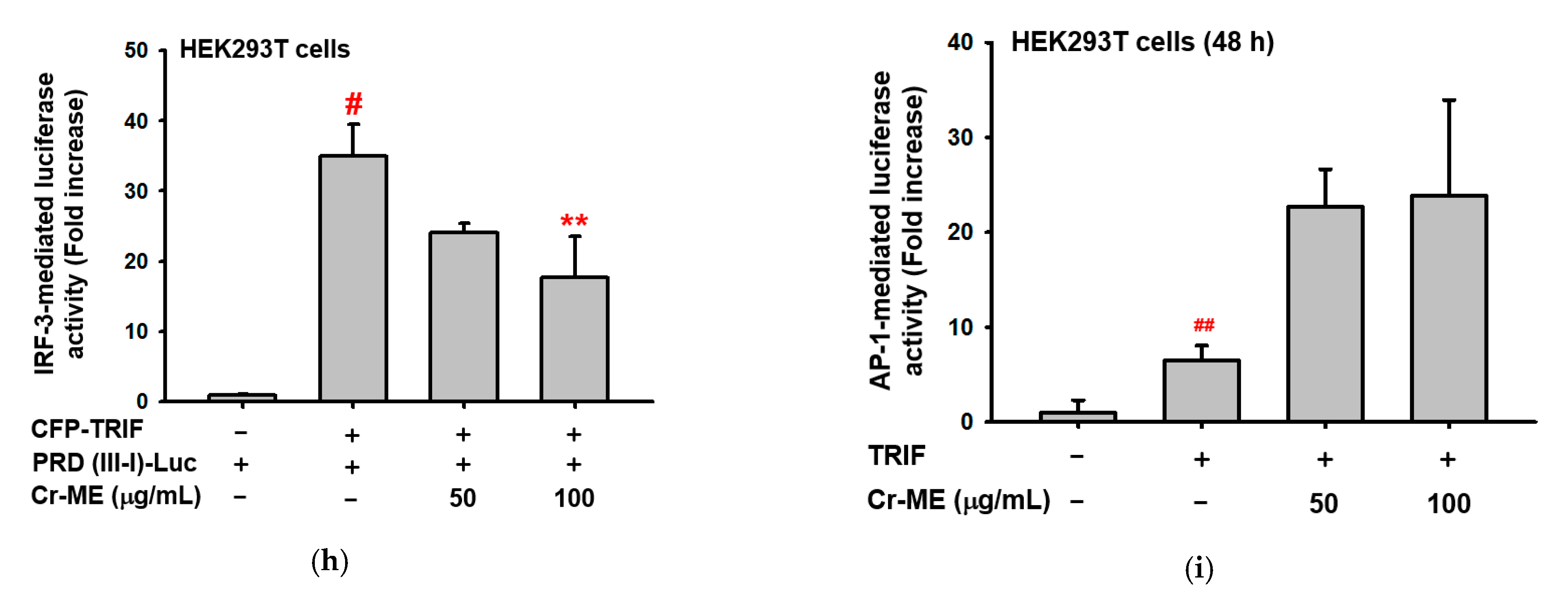
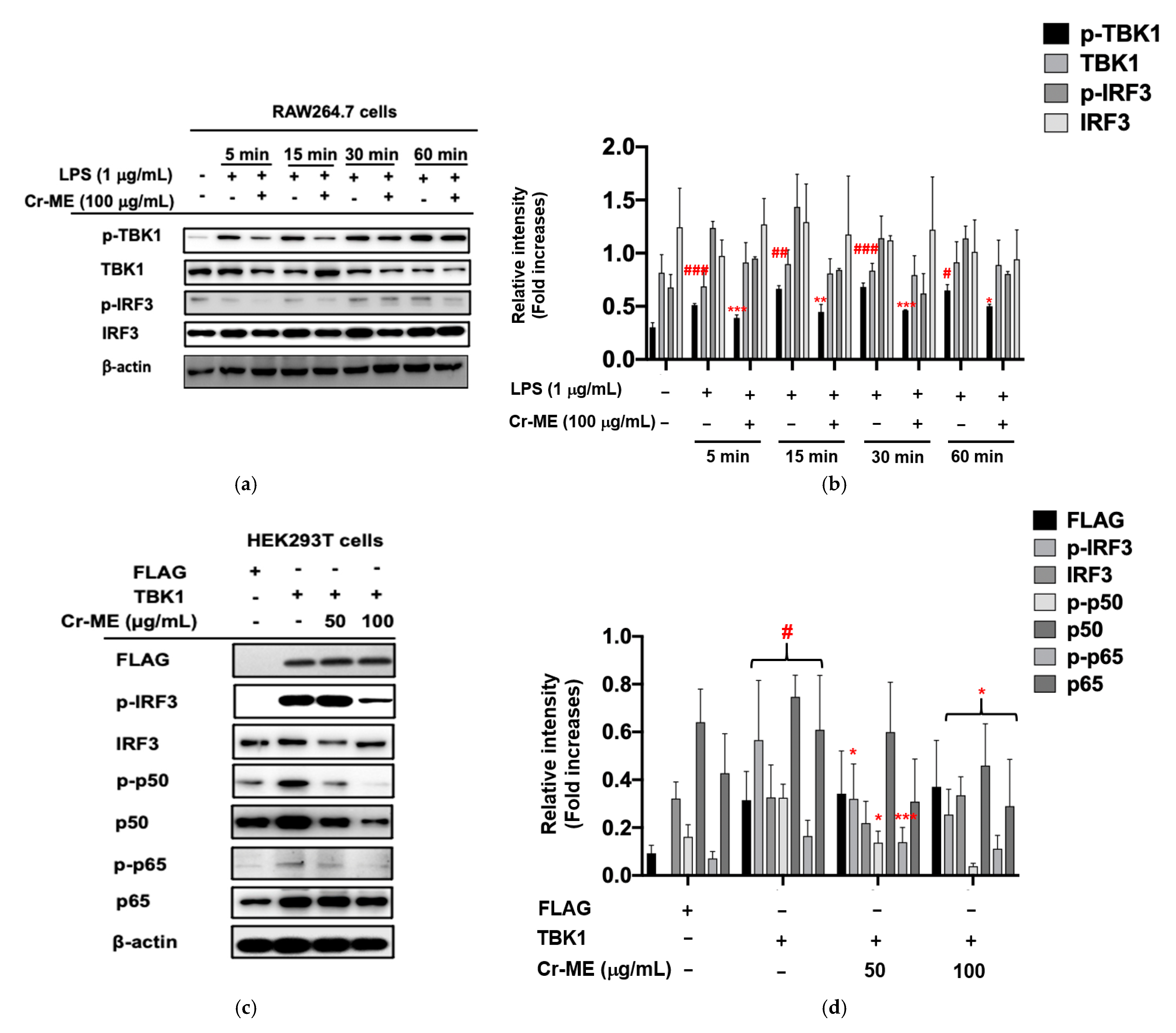
2.4. Effects of Cr-ME on LPS-Induced IRF3 Signaling and Its Upstream Enzyme TBK1 Activity
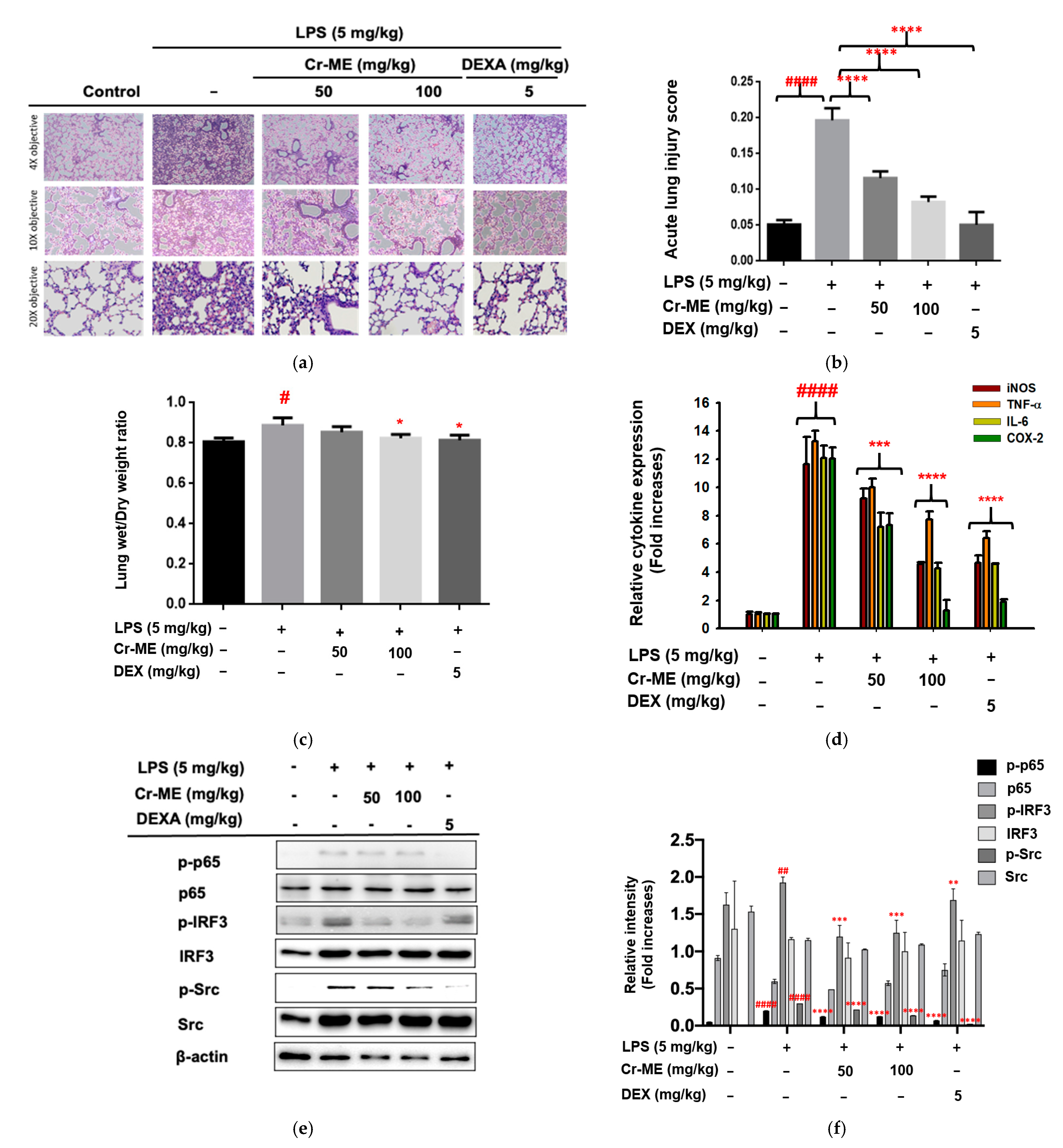
2.5. Effects of Cr-ME Treatment on LPS-Induced Acute Lung Injury (ALI)
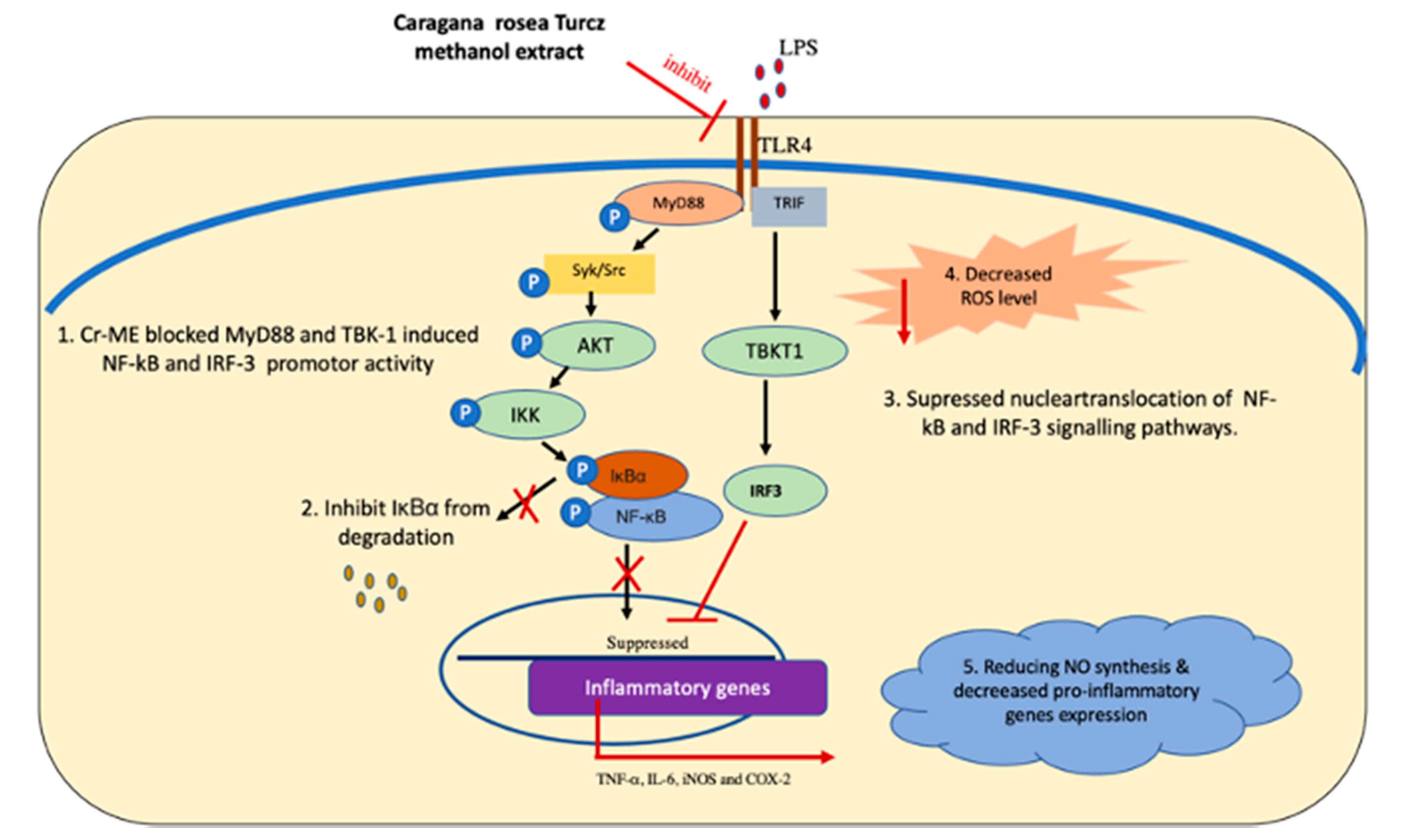
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Collection, Specimen Information, and Cr-ME Preparation
4.3. Cell Culture
4.4. Drug Treatment
4.5. Determination of NO Production by Griess Assay
4.6. Cell Viability Assay (MTT)
4.7. ROS Generation Assay
4.8. DPPH Assays
4.9. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
4.10. Gene Expression Determined by Semi-Quantitative RT-PCR and Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
4.11. Luciferase Reporter Gene Assay
4.12. Protein Lysate Isolation, Nuclear Extracts and Western Blotting Analysis
4.13. Gene Overexpression
4.14. Cellular Thermal Shift Assay (CETSA)
4.15. Lipopolysaccharide Induced Acute Lung Injury in Mice
4.16. Effects of Cr-ME on the Wet/Dry Lung Weight Ratio
4.17. Histological Analysis
4.18. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TAK1 | Transforming growth factor-β-activated kinase 1 |
| NF-κB | Nuclear factor-κB |
| AP-1 | Activator protein-1 |
| MAPKs | Mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| IRF3 | Interferon regulatory factor 3 |
| PI3K | Phosphoinositide 3-kinase |
| PGE2 | Prostaglandin E2 |
| COX-2 | Cyclooxygenase-2 |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin 6 |
| TNF-α | Tumor necrosis factor alpha |
| MKK | Mitogen-activated protein kinase |
| iNOS | Inducible nitric oxide synthase |
References
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nat. Med. 2007, 13, 552–559. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef]
- Hennessy, E.J.; Parker, A.E.; O’Neill, L.A. Targeting Toll-like receptors: Emerging therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [Green Version]
- Iwanaszko, M.; Kimmel, M. NF-kappaB and IRF pathways: Cross-regulation on target genes promoter level. BMC Genom. 2015, 16, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Taniguchi, T. IRFs: Master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef]
- Zhao, T.; Yang, L.; Sun, Q.; Arguello, M.; Ballard, D.W.; Hiscott, J.; Lin, R. The NEMO adaptor bridges the nuclear factor-kappaB and interferon regulatory factor signaling pathways. Nat. Immunol. 2007, 8, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Son, Y.J.; Cho, J.Y. Thymoquinone suppresses IRF-3-mediated expression of Type IiInterferons via suppression of TBK1. Int. J. Mol. Sci. 2018, 19, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balachandran, S.; Beg, A.A. Defining emerging roles for NF-kappaB in antivirus responses: Revisiting the interferon-beta enhanceosome paradigm. PLoS Pathog. 2011, 7, e1002165. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; He, X.; Yi, Z.; Xiang, W.; Ding, Y. Chelidonine suppresses LPS-Induced production of inflammatory mediators through the inhibitory of the TLR4/NF-kappaB signaling pathway in RAW264.7 macrophages. Biomed. Pharmacother. 2018, 107, 1151–1159. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, Z.; Huang, D.; Duan, J.; Huang, C.; Sullivan, S.; Vali, K.; Yin, Y.; Zhang, M.; Wegrzyn, J.; et al. JAK/STAT3 regulated global gene expression dynamics during late-stage reprogramming process. BMC Genom. 2018, 19, 183. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Yang, D.; Liu, X.; Wang, N.; Li, B.; Cao, H.; Lu, Y.; Wei, G.; Zhou, H.; Zheng, J. Identification of a new anti-LPS agent, geniposide, from Gardenia jasminoides Ellis, and its ability of direct binding and neutralization of lipopolysaccharide in vitro and in vivo. Int. Immunopharmacol. 2010, 10, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Shalmani, A.A.; Ghahremani, M.H.; Jeivad, F.; Shadboorestan, A.; Hassanzadeh, G.; Beh-Pajooh, A.; Ganbari-Erdi, M.; Kasirzadeh, S.; Mojtahedzadeh, M.; Sabzevari, O. Monomethyl fumarate alleviates sepsis-induced hepatic dysfunction by regulating TLR-4/NF-kappaB signalling pathway. Life Sci. 2018, 215, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Sung, N.Y.; Kim, M.Y.; Cho, J.Y. Scutellarein reduces inflammatory responses by inhibiting Src kinase activity. Korean J. Physiol. Pharmacol. 2015, 19, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossen, M.J.; Yang, W.S.; Kim, D.; Aravinthan, A.; Kim, J.H.; Cho, J.Y. Thymoquinone: An IRAK1 inhibitor with in vivo and in vitro anti-inflammatory activities. Sci. Rep. 2017, 7, 42995. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.X.; Zhou, J.T.; Li, Y.Z.; Hu, C.Q. Anti-HIV bioactive stilbene dimers of Caragana rosea. Planta Med. 2005, 71, 569–571. [Google Scholar] [CrossRef]
- Guoxun, Y.; Changqi, H. A novel stilbenolignan from Caragana rosea Turcz. Chin. J. Org. Chem. 2003, 23, 873–876. [Google Scholar]
- Yang, G.X.; Qi, J.B.; Cheng, K.J.; Hu, C.Q. Anti-HIV chemical constituents of aerial parts of Caragana rosea. Yao Xue Xue Bao Acta Pharm. Sin. 2007, 42, 179–182. [Google Scholar]
- Blackford, J.A., Jr.; Antonini, J.M.; Castranova, V.; Dey, R.D. Intratracheal instillation of silica up-regulates inducible nitric oxide synthase gene expression and increases nitric oxide production in alveolar macrophages and neutrophils. Am. J. Respir. Cell Mol. Biol. 1994, 11, 426–431. [Google Scholar] [CrossRef]
- Tian, W.L.; Jiang, Z.X.; Wang, F.; Guo, R.; Tang, P.; Huang, Y.M.; Sun, L. IRF3 is involved in human acute myeloid leukemia through regulating the expression of miR-155. Biochem. Biophys. Res. Commun. 2016, 478, 1130–1135. [Google Scholar] [CrossRef]
- Kumari, M.; Wang, X.; Lantier, L.; Lyubetskaya, A.; Eguchi, J.; Kang, S.; Tenen, D.; Roh, H.C.; Kong, X.; Kazak, L.; et al. IRF3 promotes adipose inflammation and insulin resistance and represses browning. J. Clin. Investig. 2016, 126, 2839–2854. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhu, L.H.; Guo, J.; Yuan, T.; Wang, L.Q.; Li, H.; Chen, L.X. Farnesyl phenolic enantiomers as natural MTH1 inhibitors from Ganoderma sinense. Oncotarget 2017, 8, 95865–95879. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Chen, L.; Li, H. The dietary compound luteolin inhibits pancreatic cancer growth by targeting BCL-2. Food Funct. 2018, 9, 3018–3027. [Google Scholar] [CrossRef]
- Molina, D.M.; Jafari, R.; Ignatushchenko, M.; Seki, T.; Larsson, E.A.; Dan, C.; Sreekumar, L.; Cao, Y.H.; Nordlund, P. Monitoring drug target engagement in cells and tissues using the cellular thermal shift assay. Science 2013, 341, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Jang, J.; Park, J.G.; Kim, K.H.; Yoon, K.; Yoo, B.C.; Cho, J.Y. Protein arginine methyltransferase 1 (PRMT1) selective inhibitor, TC-E 5003, has anti-inflammatory properties in TLR4 signaling. Int. J. Mol. Sci. 2020, 21, 3058. [Google Scholar] [CrossRef]
- Ahuja, A.; Jeong, D.; Kim, M.Y.; Cho, J.Y. Trichosanthes tricuspidata Lour. methanol extract exhibits anti-inflammatory activity by targeting Syk, Src, and IRAK1 kinase activity. Evid. Based. Complement. Alternat. Med. 2019, 2019, 6879346. [Google Scholar] [CrossRef]
- Ahuja, A.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Ethnopharmacological properties of Artemisia asiatica: A comprehensive review. J. Ethnopharmacol. 2018, 220, 117–128. [Google Scholar] [CrossRef]
- Kim, H.; Shin, K.K.; Kim, H.G.; Jo, M.; Kim, J.K.; Lee, J.S.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, K.H.; et al. Src/NF-kappaB-targeted anti-inflammatory effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. ethanol extract. Biomolecules 2020, 10, 648. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.H.; Ohgami, K.; Shiratori, K.; Suzuki, Y.; Koyama, Y.; Yoshida, K.; Ilieva, I.; Tanaka, T.; Onoe, K.; Ohno, S. Effects of blue honeysuckle (Lonicera caerulea L.) extract on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2006, 82, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837, 837a–837d. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Hu, Z.; Fu, Q.; Song, X.; Cui, Q.; Jia, R.; Zou, Y.; He, C.; Li, L.; Yin, Z. Resveratrol mitigates lipopolysaccharide-mediated acute inflammation in rats by inhibiting the TLR4/NF-kappaBp65/MAPKs signaling cascade. Sci. Rep. 2017, 7, 45006. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Q.; Fu, Q.; Song, X.; Jia, R.; Yang, Y.; Zou, Y.; Li, L.; He, C.; Liang, X.; et al. Antiviral properties of resveratrol against pseudorabies virus are associated with the inhibition of IkappaB kinase activation. Sci. Rep. 2017, 7, 8782. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Xu, Q.S.; Du, Y.G.; Xu, J. Chitosan oligosaccharides block LPS-induced O-GlcNAcylation of NF-kappaB and endothelial inflammatory response. Carbohydr. Polym. 2014, 99, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W. Negative regulation of TBK1-mediated antiviral immunity. FEBS Lett. 2013, 587, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.; Kang, Y.G.; Kim, Y.J.; Park, W.S.; Jeong, D.; Lee, J.; Kim, D.; Cho, J.Y. Regulation of 8-hydroxydaidzein in IRF3-mediated gene expression in LPS-stimulated murine macrophages. Biomolecules 2020, 10, 238. [Google Scholar] [CrossRef] [Green Version]
- Byeon, S.E.; Yi, Y.S.; Oh, J.; Yoo, B.C.; Hong, S.; Cho, J.Y. The role of Src kinase in macrophage-mediated inflammatory responses. Mediators Inflamm. 2012, 2012, 512926. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Lee, Y.G.; Chain, B.M.; Cho, J.Y. Distinct role of spleen tyrosine kinase in the early phosphorylation of inhibitor of kappaB alpha via activation of the phosphoinositide-3-kinase and Akt pathways. Int. J. Biochem. Cell Biol. 2009, 41, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Rathee, P. Mechanism of action of flavonoids as anti-inflammatory agents: A review. Inflamm. Allergy Drug Targets Actions 2009, 8, 229–235. [Google Scholar] [CrossRef]
- Tang, Y.; Xie, D.; Gong, W.; Wu, H.; Qiang, Y. Pentahydroxy flavonoid isolated from Madhuca indica ameliorated adjuvant-induced arthritis via modulation of inflammatory pathways. Sci. Rep. 2021, 11, 17971. [Google Scholar] [CrossRef]
- Feng, Z.; Lu, X.; Gan, L.; Zhang, Q.; Lin, L. Xanthones, a promising anti-inflammatory scaffold: Structure, activity, and drug likeness analysis. Molecules 2020, 25, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chericoni, S.; Testai, L.; Calderone, V.; Flamini, G.; Nieri, P.; Morelli, I.; Martinotti, E. The xanthones gentiacaulein and gentiakochianin are responsible for the vasodilator action of the roots of Gentiana kochiana. Planta Med. 2003, 69, 770–772. [Google Scholar] [PubMed]
- Lee, D.Y.; Yun, S.M.; Song, M.Y.; Jung, K.; Kim, E.H. Cyanidin chloride induces apoptosis by inhibiting NF-κB signaling through activation of Nrf2 in colorectal cancer cells. Antioxidants 2020, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misawa, K.; Gunji, Y.; Sato, S. Concise synthesis of flavocommelin, 7-O-methylapigenin 6-C-, 4’-O-bis-β-D-glucoside, a component of the blue supramolecular pigment from Commelina communis. Carbohydr. Res. 2013, 374, 8–13. [Google Scholar] [CrossRef]
- Zhang, G.B.; Bing, F.H.; Liu, J.; Li, Z.; Liao, Y.F.; Li, J.; Dong, C.Y. Effect of total alkaloids from Commelina communis L. on lung damage by influenza virus infection. Microbiol. Immunol. 2010, 54, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol—A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. J. Funct. Foods 2017, 30, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Gosslau, A.; Chen, K.Y.; Ho, C.-T.; Li, S. Anti-inflammatory effects of characterized orange peel extracts enriched with bioactive polymethoxyflavones. Food Sci. Hum. Wellness 2014, 3, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, F.; Fang, L.; Cai, R.; Zong, C.; Qi, Y. Genkwanin inhibits proinflammatory mediators mainly through the regulation of miR-101/MKP-1/MAPK pathway in LPS-activated macrophages. PLoS ONE 2014, 9, e96741. [Google Scholar] [CrossRef]
- Li, Y.; Hong, J.; Li, H.; Qi, X.; Guo, Y.; Han, M.; Wang, X. Genkwanin nanosuspensions: A novel and potential antitumor drug in breast carcinoma therapy. Drug Deliv. 2017, 24, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.; Yoon, C.-S.; Kim, K.-W.; Lee, H.; Kim, N.; Woo, E.-R.; Kim, Y.-C.; Kang, D.G.; Lee, H.S.; Oh, H.; et al. Neuroprotective and anti-inflammatory effects of kuwanon C from αCudrania tricuspidata are mediated by heme oxygenase-1 in HT22 hippocampal cells, RAW264.7 macrophage, and BV2 microglia. Int. J. Mol. Sci. 2020, 21, 4839. [Google Scholar] [CrossRef] [PubMed]
- Duxbury, M.S.; Ito, H.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. siRNA directed against c-Src enhances pancreatic adenocarcinoma cell gemcitabine chemosensitivity. J. Am. Coll. Surg. 2004, 198, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-kappaB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.; Kim, M.Y.; Cho, J.Y. Anti-inflammatory activities of Canarium subulatum Guillaumin methanol extract operate by targeting Src and Syk in the NF-κB pathway. J. Ethnopharmacol. 2019, 238, 111848. [Google Scholar] [CrossRef]
- Misko, T.P.; Schilling, R.J.; Salvemini, D.; Moore, W.M.; Currie, M.G. A fluorometric assay for the measurement of nitrite in biological samples. Anal. Biochem. 1993, 214, 11–16. [Google Scholar] [CrossRef]
- Rho, T.; Jeong, H.W.; Hong, Y.D.; Yoon, K.; Cho, J.Y.; Yoon, K.D. Identification of a novel triterpene saponin from Panax ginseng seeds, pseudoginsenoside RT8, and its antiinflammatory activity. J. Ginseng Res. 2020, 44, 145–153. [Google Scholar] [CrossRef]
- Wu, Z.H.; Lin, C.; Liu, M.M.; Zhang, J.; Tao, Z.H.; Hu, X.C. Src inhibition can synergize with gemcitabine and reverse resistance in triple negative breast cancer cells via the AKT/c-Jun pathway. PLoS ONE. 2016, 11, e0169230. [Google Scholar] [CrossRef]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Kim, E.; Kang, Y.G.; Kim, J.H.; Kim, Y.J.; Lee, T.R.; Lee, J.; Kim, D.; Cho, J.Y. The Antioxidant and anti-inflammatory activities of 8-hydroxydaidzein (8-HD) in activated macrophage-like RAW264.7 cells. Int. J. Mol. Sci. 2018, 19, 1828. [Google Scholar] [CrossRef] [Green Version]
- Hossen, M.J.; Hong, Y.D.; Baek, K.S.; Yoo, S.; Hong, Y.H.; Kim, J.H.; Lee, J.O.; Kim, D.; Park, J.; Cho, J.Y. In vitro antioxidative and anti-inflammatory effects of the compound K-rich fraction BIOGF1K, prepared from Panax ginseng. J. Ginseng Res. 2017, 41, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, W.S.; Yu, T.; Yi, Y.S.; Park, J.G.; Jeong, D.; Kim, J.H.; Oh, J.S.; Yoon, K.; Kim, J.H.; et al. Novel anti-inflammatory function of NSC95397 by the suppression of multiple kinases. Biochem. Pharmacol. 2014, 88, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Yun, E.; Chang, W.; Kim, J. Ginsenoside Rg3 protects against iE-DAP-induced endothelial-to-mesenchymal transition by regulating the miR-139-5p-NF-kappaB axis. J. Ginseng Res. 2020, 44, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Ischenko, I.; Camaj, P.; Seeliger, H.; Kleespies, A.; Guba, M.; De Toni, E.N.; Schwarz, B.; Graeb, C.; Eichhorn, M.E.; Jauch, K.W.; et al. Inhibition of Src tyrosine kinase reverts chemoresistance toward 5-fluorouracil in human pancreatic carcinoma cells: An involvement of epidermal growth factor receptor signaling. Oncogene 2008, 27, 7212–7222. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Lee, J.; Lee, Y.G.; Byeon, S.E.; Kim, M.H.; Sohn, E.H.; Lee, Y.J.; Lee, S.G.; Cho, J.Y. In vitro and in vivo anti-inflammatory effects of ethanol extract from Acer tegmentosum. J. Ethnopharmacol. 2010, 128, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Rahmawati, L.; Aziz, N.; Oh, J.; Hong, Y.H.; Woo, B.Y.; Hong, Y.D.; Manilack, P.; Souladeth, P.; Jung, J.H.; Lee, W.S.; et al. Cissus subtetragona Planch. ameliorates inflammatory responses in LPS-induced macrophages, HCl/EtOH-induced gastritis, and LPS-induced lung injury via attenuation of Src and TAK1. Molecules 2021, 26, 6073. [Google Scholar] [CrossRef] [PubMed]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M.; Acute lung injury in animals study, G. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Scoring per Field | ||
|---|---|---|---|
| 0 | 1 | 2 | |
| Neutrophils in the alveolar space | none | 1 to 5 | >5 |
| Neutrophils in the interstitial space | none | 1 to 5 | >5 |
| Number of hyalin membrane | none | 1 | >1 |
| Amount of proteinaceous debris filling in the airspace | none | 1 | >1 |
| Alveolar septal thickening | <2× | 2×–4× | >4× |
| Targets | Direction | Sequences (5′ to 3′) |
|---|---|---|
| iNOS | Forward Reverse | GTGAAGAAAACCCCTGTGCTG AGTTCCGAGCGTCAAAGACC |
| TNF-α | Forward Reverse | TGCCTATGTCTCAGCCTCTT GAGGCCATTTGGGAACTTCT |
| IL-6 | Forward Reverse | GCCTTCTTGGGACTGATGCT TGGAAATTGGGGTAGGAAGGAC |
| COX-2 | Forward Reverse | CATCCCCTTCCTGCGAAGTT CATGGGAGTTGGGCAGTCAT |
| MMP-2 | Forward Reverse | TCCCTGGAGACCTGAGAACC GGCAAGTCTTCCGAGTAGTTT |
| MMP-9 | Forward Reverse | GCCACTTGTCGGCGATAAGG CACTGTCCACCCCTCAGAGC |
| IFN-β | Forward Reverse | GGCCTTGGGCCTCAAAGGAA GCTTGGGATCCACACTCTCCA |
| GAPDH | Forward Reverse | ACCACAGTCCATGCCATCAC CCACCACCCTGTTGCTGTAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitra, A.; Ahuja, A.; Rahmawati, L.; Kim, H.G.; Woo, B.Y.; Hong, Y.D.; Hossain, M.A.; Zhang, Z.; Kim, S.-Y.; Lee, J.; et al. Caragana rosea Turcz Methanol Extract Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Suppressing the TLR4/NF-κB/IRF3 Signaling Pathways. Molecules 2021, 26, 6660. https://doi.org/10.3390/molecules26216660
Mitra A, Ahuja A, Rahmawati L, Kim HG, Woo BY, Hong YD, Hossain MA, Zhang Z, Kim S-Y, Lee J, et al. Caragana rosea Turcz Methanol Extract Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Suppressing the TLR4/NF-κB/IRF3 Signaling Pathways. Molecules. 2021; 26(21):6660. https://doi.org/10.3390/molecules26216660
Chicago/Turabian StyleMitra, Ankita, Akash Ahuja, Laily Rahmawati, Han Gyung Kim, Byoung Young Woo, Yong Deog Hong, Mohammad Amjad Hossain, Zhiyun Zhang, Soo-Yong Kim, Jongsung Lee, and et al. 2021. "Caragana rosea Turcz Methanol Extract Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Suppressing the TLR4/NF-κB/IRF3 Signaling Pathways" Molecules 26, no. 21: 6660. https://doi.org/10.3390/molecules26216660