
Proteomic Signature of Extracellular Vesicles for Lung Cancer Recognition

 ,
,  ,
,
Abstract
:
1. Introduction
2. Results
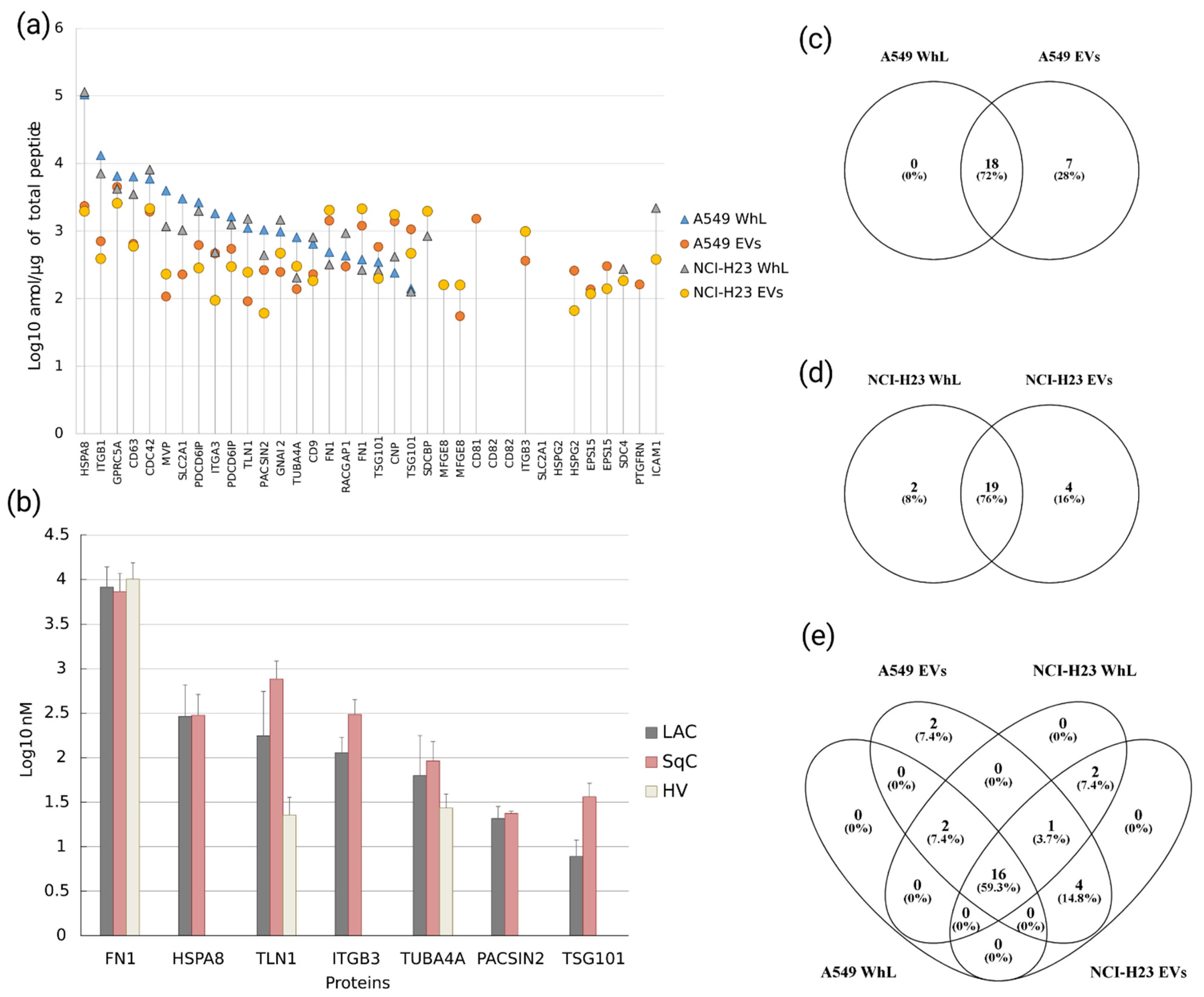
2.1. Verification of Protein Levels of 28 EV-Associated Markers in A549 and NCI-H23 Cell Lines by SRM Analysis
2.2. SRM/SIS Analysis of EV-Associated Proteins in the Blood Plasma of LC Patients and Healthy Volunteers Reveals the Proteomic Signature of EVs
2.3. Proteomic Signature of EVs Distinguishes Lung Adenocarcinoma and Squamous Cell Carcinoma Histological Types, and Its Components Are Involved in Focal Adhesion
3. Discussion
4. Materials and Methods
4.1. Cultivation of A549 and NCI-H23 Cell Lines and EV Isolation
4.2. Clinical Sample Description
4.3. Sample Preparation for Mass Spectrometry Analysis
4.4. Synthesis of SISs
4.5. Quantitative Analysis of EV-Associated Proteins by Targeted Mass Spectrometry
4.6. Statistical and Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.-W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta Rev. Cancer 2015, 1856, 189–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiles, C. Lung Cancer Screening with Low-Dose Computed Tomography. Radiol. Clin. N. Am. 2014, 52, 27–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudkerk, M.; Liu, S.; Heuvelmans, M.A.; Walter, J.E.; Field, J.K. Lung cancer LDCT screening and mortality reduction—Evidence, pitfalls and future perspectives. Nat. Rev. Clin. Oncol. 2021, 18, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Oudkerk, M.; Devaraj, A.; Vliegenthart, R.; Henzler, T.; Prosch, H.; Heussel, C.P.; Bastarrika, G.; Sverzellati, N.; Mascalchi, M.; Delorme, S.; et al. European position statement on lung cancer screening. Lancet Oncol. 2017, 18, e754–e766. [Google Scholar] [CrossRef]
- Ignatiadis, M.; Sledge, G.W.; Jeffrey, S.S. Liquid biopsy enters the clinic—Implementation issues and future challenges. Nat. Rev. Clin. Oncol. 2021, 18, 297–312. [Google Scholar] [CrossRef]
- Bandu, R.; Oh, J.W.; Kim, K.P. Mass spectrometry-based proteome profiling of extracellular vesicles and their roles in cancer biology. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, D.W.; Cho, J.-Y. Exploring the key communicator role of exosomes in cancer microenvironment through proteomics. Proteome Sci. 2019, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Balbinotti, H.; Cadore, N.A.; Dutra, C.S.; Da Silva, E.D.; Ferreira, H.B.; Zaha, A.; Monteiro, K.M. Protein Profiling of Extracellular Vesicles Associated with Cisplatin Resistance in Lung Cancer. Anticancer Res. 2020, 40, 5509–5516. [Google Scholar] [CrossRef]
- Wang, N.; Song, X.; Liu, L.; Niu, L.; Wang, X.; Song, X.; Xie, L. Circulating exosomes contain protein biomarkers of metastatic non-small-cell lung cancer. Cancer Sci. 2018, 109, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Kuang, M.; Tao, X.; Peng, Y.; Zhang, W.; Pan, Y.; Cheng, L.; Yuan, C.; Zhao, Y.; Mao, H.; Zhuge, L.; et al. Proteomic analysis of plasma exosomes to differentiate malignant from benign pulmonary nodules. Clin. Proteom. 2019, 16, 1–11. [Google Scholar] [CrossRef]
- Kopylov, A.; Ponomarenko, E.A.; Ilgisonis, E.V.; Pyatnitskiy, M.A.; Lisitsa, A.V.; Poverennaya, E.V.; Kiseleva, O.I.; Farafonova, T.E.; Tikhonova, O.V.; Zavialova, M.G.; et al. 200+ Protein Concentrations in Healthy Human Blood Plasma: Targeted Quantitative SRM SIS Screening of Chromosomes 18, 13, Y, and the Mitochondrial Chromosome Encoded Proteome. J. Proteome Res. 2018, 18, 120–129. [Google Scholar] [CrossRef]
- Novikova, S.E.; Vakhrushev, I.V.; Tsvetkova, A.V.; Shushkova, N.A.; Farafonova, T.E.; Yarygin, K.N.; Zgoda, V.G. Proteomics of Transcription Factors: Identification of a Pool of HL-60 Cell Line-Specific Regulatory Proteins. Biochem. Suppl. Ser. B Biomed. Chem. 2019, 13, 329–340. [Google Scholar] [CrossRef]
- Novikova, S.E.; Farafonova, T.E.; Tikhonova, O.V.; Shushkova, N.A.; Pyatnitsky, M.A.; Zgoda, V.G.; Ponomarenko, E.A.; Lisitsa, A.V.; Grigoryev, A.I.; Tutelyan, V.A.; et al. Mass-Spectrometric MRM Analysis of FDA-Approved Proteins in Plasma of Healthy Volunteers. Biochem. Suppl. Ser. B Biomed. Chem. 2021, 15, 40–61. [Google Scholar] [CrossRef]
- Fukuda, T.; Nomura, M.; Kato, Y.; Tojo, H.; Fujii, K.; Nagao, T.; Bando, Y.; Fehniger, T.E.; Marko-Varga, G.; Nakamura, H.; et al. A selected reaction monitoring mass spectrometric assessment of biomarker candidates diagnosing large-cell neuroendocrine lung carcinoma by the scaling method using endogenous references. PLoS ONE 2017, 12, e0176219. [Google Scholar] [CrossRef]
- Nishimura, T.; Nomura, M.; Tojo, H.; Hamasaki, H.; Fukuda, T.; Fujii, K.; Mikami, S.; Bando, Y.; Kato, H. Proteomic analysis of laser-microdissected paraffin-embedded tissues: (2) MRM assay for stage-related proteins upon non-metastatic lung adenocarcinoma. J. Proteom. 2010, 73, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Schmidlin, T.; Garrigues, L.; Lane, C.S.; Mulder, T.C.; Van Doorn, S.; Post, H.; de Graaf, E.L.; Lemeer, S.; Heck, A.; Altelaar, A.F.M. Assessment of SRM, MRM3, and DIA for the targeted analysis of phosphorylation dynamics in non-small cell lung cancer. Proteomics 2016, 16, 2193–2205. [Google Scholar] [CrossRef] [PubMed]
- Novikova, S.; Shushkova, N.; Farafonova, T.; Tikhonova, O.; Kamyshinsky, R.; Zgoda, V. Proteomic Approach for Searching for Universal, Tissue-Specific, and Line-Specific Markers of Extracellular Vesicles in Lung and Colorectal Adenocarcinoma Cell Lines. Int. J. Mol. Sci. 2020, 21, 6601. [Google Scholar] [CrossRef] [PubMed]
- Shushkova, N.A.; Vavilov, N.E.; Novikova, S.; Farafonova, T.E.; Tikhonova, O.V.; Liao, P.-C.; Zgoda, V.G. Quantitative Proteomics of Human Blood Exosomes. Biochem. Suppl. Ser. B Biomed. Chem. 2019, 13, 132–139. [Google Scholar] [CrossRef]
- Lee, C.-H.; Im, E.-J.; Moon, P.-G.; Baek, M.-C. Discovery of a diagnostic biomarker for colon cancer through proteomic profiling of small extracellular vesicles. BMC Cancer 2018, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Garcia, L.M.C.; Peterson, T.E.; Cepeda, M.A.; Johnson, A.J.; Parney, I.F. Isolation and Analysis of Plasma-Derived Exosomes in Patients with Glioma. Front. Oncol. 2019, 9, 651. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.N.; Yin, H. Polymer-Based Purification of Extracellular Vesicles. Methods Mol. Biol. 2017, 1660, 91–103. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef]
- Beckler, M.D.; Higginbotham, J.N.; Franklin, J.L.; Ham, A.-J.; Halvey, P.J.; Imasuen, I.E.; Whitwell, C.; Li, M.; Liebler, D.; Coffey, R.J. Proteomic Analysis of Exosomes from Mutant KRAS Colon Cancer Cells Identifies Intercellular Transfer of Mutant KRAS. Mol. Cell. Proteom. 2013, 12, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Deng, T.; Liu, R.; Bai, M.; Zhou, L.; Wang, X.; Li, S.; Wang, X.; Yang, H.; Li, J.; et al. Exosome-delivered EGFR regulates liver microenvironment to promote gastric cancer liver metastasis. Nat. Commun. 2017, 8, 15016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, J.; Caja, S.; Moraes, M.C.S.; Couto, N.; Costa-Silva, B. Exosome-Based Cell-Cell Communication in the Tumor Microenvironment. Front. Cell Dev. Biol. 2018, 6, 18. [Google Scholar] [CrossRef]
- Othman, N.; Jamal, R.; Abu, N. Cancer-Derived Exosomes as Effectors of Key Inflammation-Related Players. Front. Immunol. 2019, 10, 2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shushkova, N.A.; Novikova, S.E.; Zgoda, V.G. Exosomes of Malignant Tumors: Prospects of Omics Diagnostics. Biochem. Suppl. Ser. B Biomed. Chem. 2020, 14, 105–115. [Google Scholar] [CrossRef]
- Torrisi, M.R.; Lotti, L.V.; Belleudi, F.; Gradini, R.; Salcini, A.E.; Confalonieri, S.; Pelicci, P.G.; Di Fiore, P.P. Eps15 Is Recruited to the Plasma Membrane upon Epidermal Growth Factor Receptor Activation and Localizes to Components of the Endocytic Pathway during Receptor Internalization. Mol. Biol. Cell 1999, 10, 417–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kreuk, B.-J.; Anthony, E.C.; Geerts, D.; Hordijk, P.L. The F-BAR Protein PACSIN2 Regulates Epidermal Growth Factor Receptor Internalization. J. Biol. Chem. 2012, 287, 43438–43453. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.L.; Fransen, M.; Brees, C.; Ghys, K.; Amery, L.; Mannaerts, G.P.; Ladant, D.; Van Veldhoven, P.P. The Human Plasma Proteome: History, Character, and Diagnostic Prospects. Mol. Cell. Proteom. 2003, 2, 50–252. [Google Scholar] [CrossRef] [Green Version]
- Kopylov, A.; Ilgisonis, E.V.; Moysa, A.; Tikhonova, O.V.; Zavialova, M.; Novikova, S.; Lisitsa, A.V.; Ponomarenko, E.A.; Moshkovskii, S.A.; Markin, A.A.; et al. Targeted Quantitative Screening of Chromosome 18 Encoded Proteome in Plasma Samples of Astronaut Candidates. J. Proteome Res. 2016, 15, 4039–4046. [Google Scholar] [CrossRef]
- Wang, J.P.; Hielscher, A. Fibronectin: How Its Aberrant Expression in Tumors May Improve Therapeutic Targeting. J. Cancer 2017, 8, 674–682. [Google Scholar] [CrossRef] [Green Version]
- Schor, S.L.; Ellis, I.R.; Jones, S.J.; Baillie, R.; Seneviratne, K.; Clausen, J.; Motegi, K.; Vojtesek, B.; Kankova, K.; Furrie, E.; et al. Migration-stimulating factor: A genetically truncated onco-fetal fibronectin isoform expressed by carcinoma and tumor-associated stromal cells. Cancer Res. 2003, 63, 8827–8836. [Google Scholar]
- Desiniotis, A.; Kyprianou, N. Significance of Talin in Cancer Progression and Metastasis. Int. Rev. Cell Mol. Biol. 2011, 289, 117–147. [Google Scholar] [CrossRef] [Green Version]
- Everley, P.A.; Krijgsveld, J.; Zetter, B.R.; Gygi, S.P. Quantitative Cancer Proteomics: Stable Isotope Labeling with Amino Acids in Cell Culture (SILAC) as a Tool for Prostate Cancer Research. Mol. Cell. Proteom. 2004, 3, 729–735. [Google Scholar] [CrossRef] [Green Version]
- Fang, K.-P.; Dai, W.; Ren, Y.-H.; Xu, Y.-C.; Zhang, S.-M.; Qian, Y.-B. Both Talin-1 and Talin-2 correlate with malignancy potential of the human hepatocellular carcinoma MHCC-97 L cell. BMC Cancer 2016, 16, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, M.-T.; Hua, C.-H.; Tsai, M.-H.; Wan, L.; Lin, Y.-J.; Chen, C.-M.; Chiu, I.-W.; Chan, C.; Tsai, F.-J.; Sheu, J.J.-C. Talin-1 overexpression defines high risk for aggressive oral squamous cell carcinoma and promotes cancer metastasis. J. Pathol. 2011, 224, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-F.; Ren, X.-Y.; Li, Y.-Q.; Mu-Sheng, Z.; Tang, X.-R.; Sun, Y.; Shao, J.-Y.; Jian-Yong, S.; Kang, T.-B.; Zeng, M.-S.; et al. High expression of Talin-1 is associated with poor prognosis in patients with nasopharyngeal carcinoma. BMC Cancer 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Chen, H.-J.; Chen, S.-H.; Xue, X.-Y.; Chen, H.; Zheng, Q.-S.; Wei, Y.; Li, X.-D.; Huang, J.-B.; Cai, H.; et al. Upregulation of Talin-1 expression associates with advanced pathological features and predicts lymph node metastases and biochemical recurrence of prostate cancer. Medicine 2016, 95, e4326. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, H.; Wang, K.; Shang, X.; Chen, X.; Zheng, L.; Yu, J. Expression of Talin1 in tissues of ovarian cancer and its role in invasion and migration of ovarian cancer cells. Int. J. Clin. Exp. Med. 2017, 10, 410–417. [Google Scholar]
- Sherman, M.Y.; Gabai, V.L. Hsp70 in cancer: Back to the future. Oncogene 2015, 34, 4153–4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Shan, N.; Zhou, W.; Zhang, S. Identification of HSPA8 as a candidate biomarker for endometrial carcinoma by using iTRAQ-based proteomic analysis. OncoTargets Ther. 2016, 9, 2169–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ge, Z. High HSPA8 expression predicts adverse outcomes of acute myeloid leukemia. BMC Cancer 2021, 21, 1–11. [Google Scholar] [CrossRef]
- Tang, T.; Yang, C.; Brown, H.E.; Huang, J. Circulating Heat Shock Protein 70 Is a Novel Biomarker for Early Diagnosis of Lung Cancer. Dis. Markers 2018, 2018, 1–8. [Google Scholar] [CrossRef]
- Li, L.; Cohen, S.N. tsg101: A Novel Tumor Susceptibility Gene Isolated by Controlled Homozygous Functional Knockout of Allelic Loci in Mammalian Cells. Cell 1996, 85, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.B.; Stanton, M.J.; West, W.W.; Todd, G.L.; Wagner, K.-U. Tsg101 is upregulated in a subset of invasive human breast cancers and its targeted overexpression in transgenic mice reveals weak oncogenic properties for mammary cancer initiation. Oncogene 2007, 26, 5950–5959. [Google Scholar] [CrossRef] [Green Version]
- Young, T.W.; Rosen, D.G.; Mei, F.C.; Li, N.; Liu, J.; Wang, X.-F.; Cheng, X. Up-regulation of Tumor Susceptibility Gene 101 Conveys Poor Prognosis through Suppression of p21 Expression in Ovarian Cancer. Clin. Cancer Res. 2007, 13, 3848–3854. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Tian, Z.; Cao, K.; Zhang, B.; Wen, Q.; Zhou, X.; Yang, W.; Wang, T.; Shi, H.; Wang, R. TSG101 promotes the proliferation, migration and invasion of hepatocellular carcinoma cells by regulating the PEG10. J. Cell. Mol. Med. 2019, 23, 70–82. [Google Scholar] [CrossRef]
- Hurwitz, S.N.; Rider, M.A.; Bundy, J.L.; Liu, X.; Singh, R.K.; Meckes, D.G., Jr. Proteomic profiling of NCI-60 extracellular vesicles uncovers common protein cargo and cancer type-specific biomarkers. Oncotarget 2016, 7, 86999–87015. [Google Scholar] [CrossRef]
- Goksuluk, D.; Korkmaz, S.; Zararsiz, G.; Karaagaoglu, E.A. easyROC: An Interactive Web-tool for ROC Curve Analysis Using R Language Environment. R J. 2016, 8, 213–230. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAC | SqC | HV | |
|---|---|---|---|
| Total | 23 | 11 | 23 |
| Age | 46–77 | 46–73 | 23–42 |
| Male | 15 | 9 | 10 |
| Female | 8 | 2 | 13 |
| Stage 1, 1A, and 1B | 10 | 3 | - |
| Stage 2, 2A, and 2B | 4 | 3 | - |
| Stage 3, 3A, and 3B | 6 | 4 | - |
| Stage 4 | 3 | 1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novikova, S.E.; Soloveva, N.A.; Farafonova, T.E.; Tikhonova, O.V.; Liao, P.-C.; Zgoda, V.G. Proteomic Signature of Extracellular Vesicles for Lung Cancer Recognition. Molecules 2021, 26, 6145. https://doi.org/10.3390/molecules26206145
Novikova SE, Soloveva NA, Farafonova TE, Tikhonova OV, Liao P-C, Zgoda VG. Proteomic Signature of Extracellular Vesicles for Lung Cancer Recognition. Molecules. 2021; 26(20):6145. https://doi.org/10.3390/molecules26206145
Chicago/Turabian StyleNovikova, Svetlana E., Natalia A. Soloveva, Tatiana E. Farafonova, Olga V. Tikhonova, Pao-Chi Liao, and Victor G. Zgoda. 2021. "Proteomic Signature of Extracellular Vesicles for Lung Cancer Recognition" Molecules 26, no. 20: 6145. https://doi.org/10.3390/molecules26206145