
Very Stable Two Mega Dalton High-Molecular-Mass Multiprotein Complex from Sea Cucumber Eupentacta fraudatrix

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation and Analysis of Sea Cucumbers Protein Complex
2.2. Light Scattering Assay
2.3. SDS-PAGE Analysis of the Complex Proteins
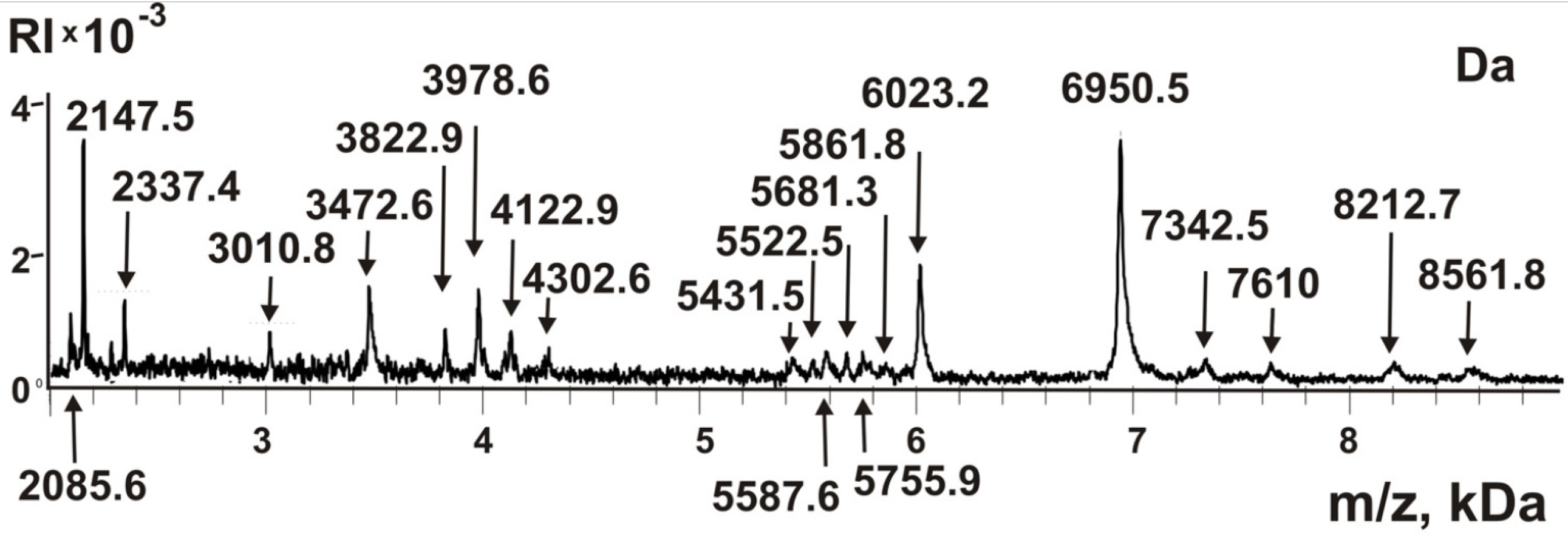
2.4. Peptides of the Complex
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Sea cucumber Extracts Preparation
4.3. Purification of Stable Complexes by Gel Filtration
4.4. SDS-PAGE Assay
4.5. Analysis of Peptides and Small Proteins
4.6. MALDI Mass Spectrometry Analysis of Proteins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Hyman, L.H. The invertebrates: Echinodermata. In The Coelome Bilateria; McGraw-Hill Book Co.: New York, NY, USA, 1955; p. 763. [Google Scholar]
- Dolmatov, I.Y.; Mashanov, V.S. Regeneration in Holothurians; Dalnauka: Vladivostok, Russia, 2007; pp. 1–212. [Google Scholar]
- Dolmatov, I.Y. Regeneration of the digestive system in holothurians. Zhurnal Obshch. Biol. 2009, 70, 319–330. [Google Scholar]
- Dolmatov, I.Y. Regeneration of the aquapharyngeal complex in the holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). In Keys for Regeneration; Taban, C.H., Boilly, B., Eds.; Karger: Basel, Germany, 1992; Volume 23, pp. 40–50. [Google Scholar]
- Levin, V.S. Dalnevostochnyi Trepang. Biologiya, Dobycha, Vosproizvodstvo (The Far East. Trepang. Biology, Catching, Reproduction); STMicroelectronics: Petersburg, Russia, 2000; pp. 1–201. [Google Scholar]
- Alberts, B. The cell as a collection of protein machines: Preparing the next generation of molecular biologists. Cell 1998, 92, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Eubel, H.; Braun, H.P.; Millar, A.H. Blue-native PAGE in plants: A tool in analysis of protein-protein interactions. Plant. Methods 2005, 1, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Wang, L.; Liang, G. Identification and analysis of multi-protein complexes in placenta. PLoS ONE 2013, 138, 62988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soboleva, S.E.; Dmitrenok, P.S.; Verkhovod, T.D.; Buneva, V.N.; Sedykh, S.E.; Nevinsky, G.A. Very stable high molecular mass multiprotein complex with DNase and amylase activities in human milk. J. Mol. Recognit. 2015, 28, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Burkova, E.E.; Dmitrenok, P.S.; Sedykh, S.E.; Buneva, V.N.; Soboleva, S.E.; Nevinsky, G.A. Extremely stable soluble high molecular mass multi-protein complex with DNase activity in human placental tissue. PLoS ONE 2014, 9, 111234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkova, E.E.; Dmitrenok, P.S.; Bulgakov, D.V.; Ermakov, E.A.; Buneva, V.N.; Soboleva, S.E.; Nevinsky, G.A. Identification of major proteins of a very stable high molecular mass multi-protein complex of human placental tissue possessing nine different catalytic activities. Biochem. Anal. Biochem. 2018, 7, 351. [Google Scholar] [CrossRef]
- Soboleva, S.E.; Burkova, E.E.; Dmitrenok, P.S.; Bulgakov, D.V.; Menzorova, N.I.; Buneva, V.N.; Nevinsky, G.A. Extremely stable high molecular mass soluble multiprotein complex from eggs of sea urchin Strongylocentrotus intermedius with phosphatase activity. J. Mol. Recognit. 2018, 31, 2753. [Google Scholar] [CrossRef] [PubMed]
- Welker, M. Proteomics for routine identification of microorganisms. Proteomics 2011, 11, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Pislyagin, E.A.; Manzhulo, I.V.; Gorpenchenko, T.Y.; Dmitrenok, P.S.; Avilov, S.A.; Silchenko, A.S.; Wang, Y.M.; Aminin, D.L. Cucumarioside A2-2 causes macrophage activation in mouse spleen. Mar. Drugs 2017, 15, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pislyagin, E.A.; Manzhulo, I.V.; Dmitrenok, P.S.; Aminin, D.L. Cucumarioside A2-2 causes changes in the morphology and proliferative activity in mouse spleen. Acta Histochem. 2016, 118, 387–392. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.M.; Kostrikina, I.A.; Dmitrenok, P.S.; Soboleva, S.E.; Nevinsky, G.A. Very Stable Two Mega Dalton High-Molecular-Mass Multiprotein Complex from Sea Cucumber Eupentacta fraudatrix. Molecules 2021, 26, 5703. https://doi.org/10.3390/molecules26185703
Timofeeva AM, Kostrikina IA, Dmitrenok PS, Soboleva SE, Nevinsky GA. Very Stable Two Mega Dalton High-Molecular-Mass Multiprotein Complex from Sea Cucumber Eupentacta fraudatrix. Molecules. 2021; 26(18):5703. https://doi.org/10.3390/molecules26185703
Chicago/Turabian StyleTimofeeva, Anna M., Irina A. Kostrikina, Pavel S. Dmitrenok, Svetlana E. Soboleva, and Georgy A. Nevinsky. 2021. "Very Stable Two Mega Dalton High-Molecular-Mass Multiprotein Complex from Sea Cucumber Eupentacta fraudatrix" Molecules 26, no. 18: 5703. https://doi.org/10.3390/molecules26185703