
Chemical Transformation of Humic Acid Molecules under the Influence of Mineral, Fungal and Bacterial Fertilization in the Context of the Agricultural Use of Degraded Soils


Abstract
:1. Introduction
2. Results and Discussion
2.1. Structural Properties of HAs before the Addition of Fertilizers and Microorganisms
2.2. Changes in Elemental Composition and Atomic Ratios of HAs from Soils Treated with Mineral and Microbiological Additives
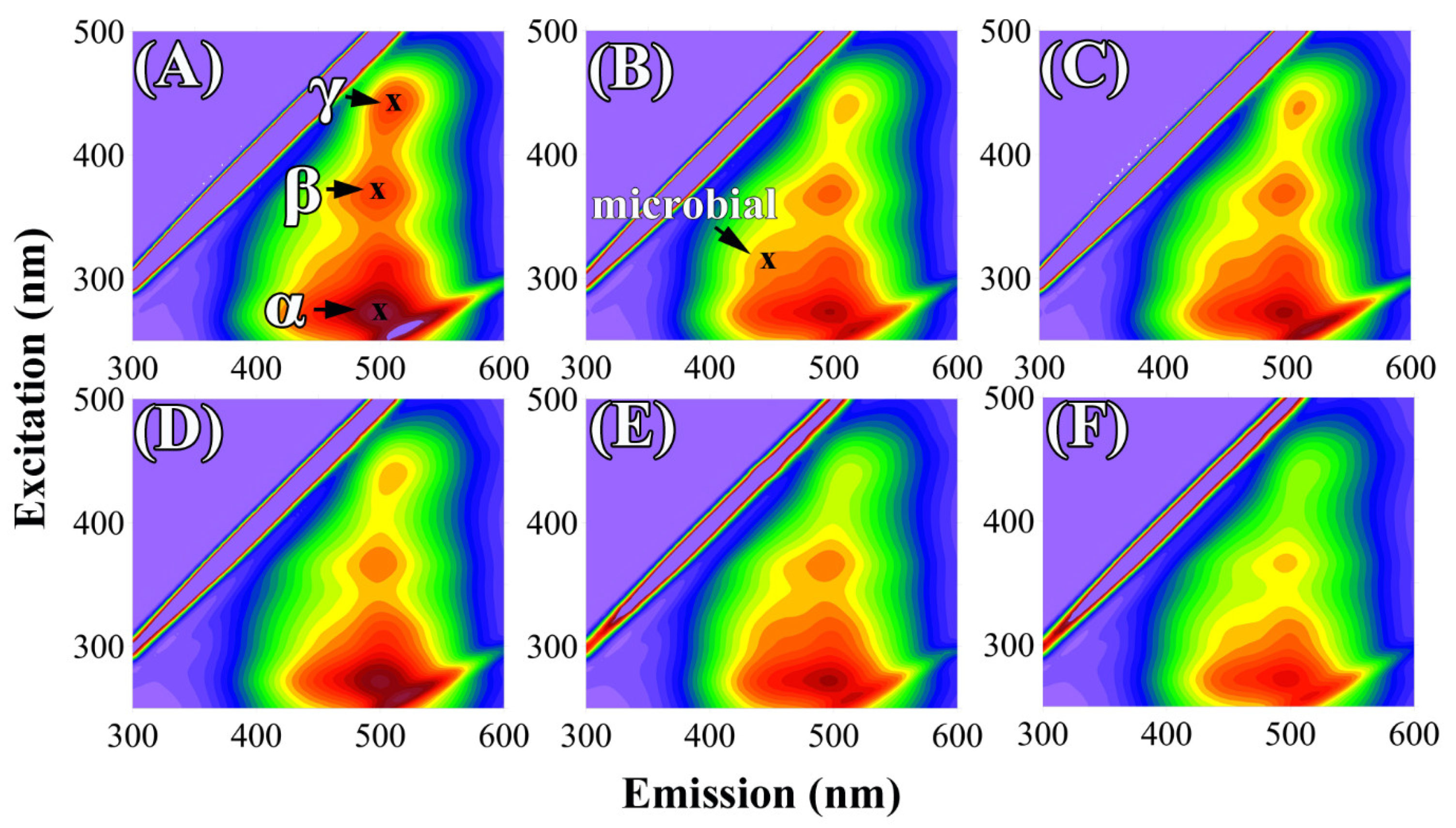
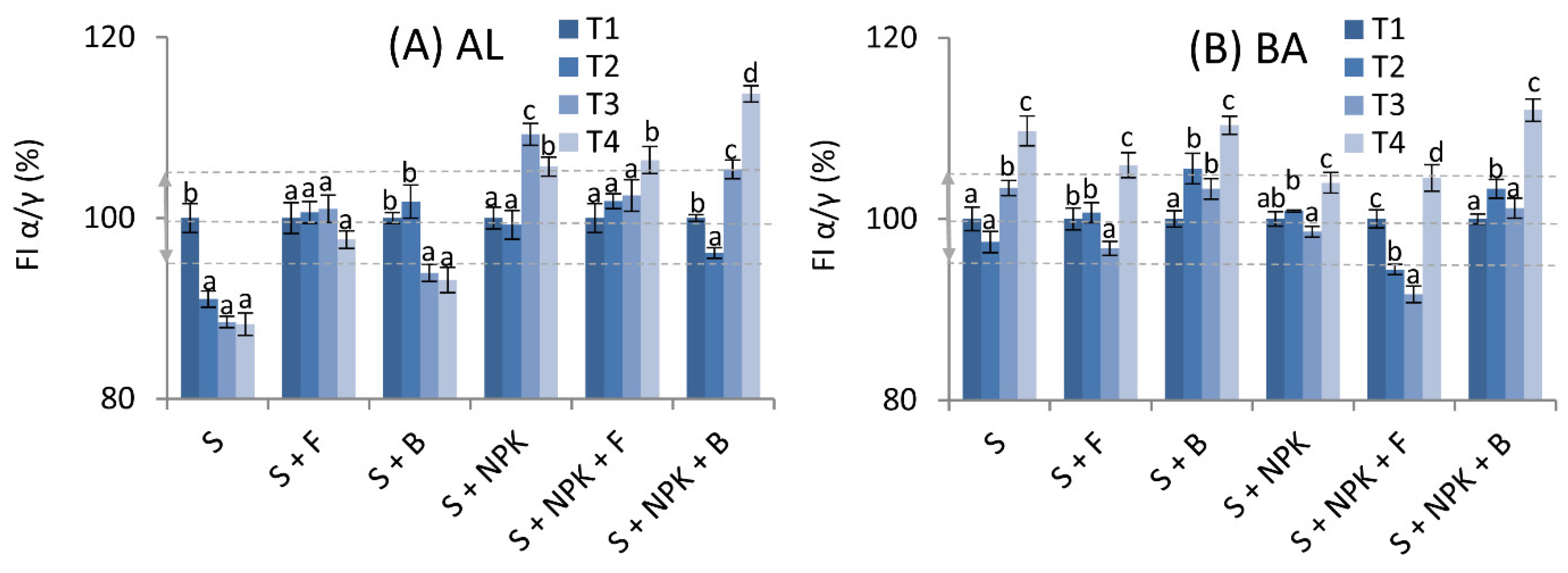
2.3. Changes in Fluorescence, and Structural and Humification Parameters of HAs from Soils Treated with Mineral Fertilizers, and Fungal and Bacterial Strains
2.4. Changes in Degree of Humification of HAs Derived from Soils with Mineral Fertilization, and Fungal and Bacterial Strains Based on UV-Vis Spectroscopy
2.5. Changes in the Content of HAs in AL and BA Soils
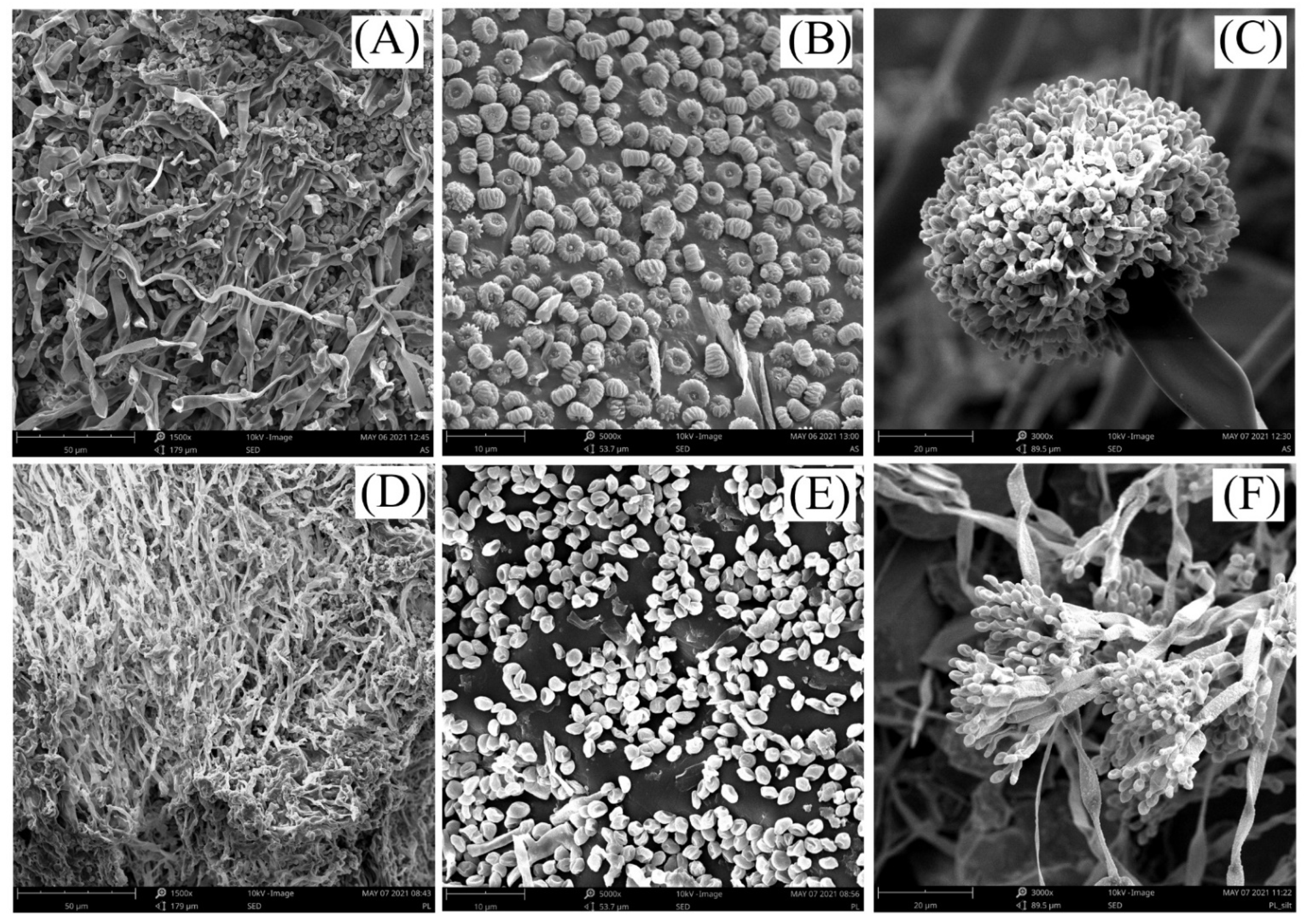
2.6. Agronomic Implications and Further Research
3. Materials and Methods
3.1. Location and Physicochemical Properties of Soils Used in Field Experiments
3.2. Selection of Mineral Fertilizers, and Fungal and Bacterial Strains
3.3. Experiment Design for Application of Mineral Fertilizers, Bacterial and Fungal Strains, and Soil Sampling
3.4. Analysis of Structural Changes and Humification of Humic Acids during Treatment with Microorganisms and Fertilizers
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- International Humic Substances Society. Available online: https://humic-substances.org/ (accessed on 13 August 2021).
- Piccolo, A.; Spaccini, R.; Nieder, R.; Richter, J. Sequestration of a biologically labile organic carbon in soils by humified organic matter. Clim. Chang. 2004, 67, 329–343. [Google Scholar] [CrossRef]
- Song, X.; Liu, S.; Liu, Q.; Zhang, W.; Hu, C. Carbon sequestration in soil humic substances under long-term fertilization in a wheat-maize system from North China. J. Integr. Agric. 2014, 13, 562–569. [Google Scholar] [CrossRef]
- Boguta, P.; Sokołowska, Z. Influence of phosphate ions on buffer capacity of soil humic acids. Int. Agroph. 2012, 26, 7–14. [Google Scholar] [CrossRef]
- Boguta, P.; Sokołowska, Z. Interactions of humic acids with metals. Acta Agroph. Monogr. 2013, 2, 1–113. [Google Scholar]
- Zhang, N.; Zhang, D.; Ji, H.; Yu, X.; Zhang, Z.; Yang, S.; Zhang, C. Redox structures of humic acids derived from different sediments and their effects on microbial reduction reactions. Front. Microbiol. 2018, 9, 1225. [Google Scholar] [CrossRef]
- Kostić, I.; Anđelković, T.; Nikolić, R.; Bojić, A.; Purenović, M.; Blagojević, S.; Darko, A. Copper(II) and lead(II) complexation by humic acid and humic-like ligands. J Serb. Chem. Soc. 2011, 76, 1325–1336. [Google Scholar] [CrossRef]
- Parvan, L.; Dumitru, M.; Sirbu, C.; Cioroianu, T. Fertilizer with humic substances. Rom. Agric. Res. 2013, 30, 205–212. [Google Scholar]
- Huculak-Mączka, M.; Hoffmann, J.; Hoffmann, K. Evaluation of the possibilities of using humic acids obtained from lignite in the production of commercial fertilizers. J. Soils Sediments 2018, 18, 2868–2880. [Google Scholar] [CrossRef] [Green Version]
- Cybulak, M.; Sokołowska, Z.; Boguta, P. The influence of biochar on the content of carbon and the chemical transformations of fallow and grassland humic acids. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Kononova, M.M. Soil Organic Matter: Its Nature, Its Role in Soil Formation and in Soil Fertility; Pergamon Press: Oxford, UK, 1966. [Google Scholar]
- Steelink, C. Investigating humic acids in soils. Anal. Chem. 2002, 1, 326–333. [Google Scholar]
- Sokołowska, Z.; Szajdak, L.; Boguta, P. Effect of phosphates on dissolved organic matter release from peat-muck soils. Int. Agrophys. 2011, 25, 173–180. [Google Scholar]
- De Mastro, F.; Cocozza, C.; Traversa, A.; Savy, D.; Abdelrahman, H.M.; Brunetti, G. Influence of crop rotation, tillage and fertilization on chemical and spectroscopic characteristics of humic acids. PLoS ONE 2019, 14, e0219099. [Google Scholar] [CrossRef]
- Shaffer, L.; Wandruszka, R. Temperature induced aggregation and clouding in humic acid solutions. Adv. Environ. Chem. 2015, 2015, 543614. [Google Scholar] [CrossRef] [Green Version]
- Koukol, O.; Gryndler, M.; Novák, F.; Vosátka, M. Effect of chalara longipes on decomposition of humic acids from picea abies needle litter. Folia Microbiol. 2004, 49, 574–578. [Google Scholar] [CrossRef]
- Řezáčová, V.; Gryndler, M. Fluorescence spectroscopy: A tool to characterize humic substances in soil colonized by microorganisms? Folia Microbiol. 2006, 51, 215–221. [Google Scholar] [CrossRef]
- Khundzhua, D.A.; Patsaeva, S.V.; Terekhova, V.A.; Yuzhakov, V.I. Spectral characterization of fungal metabolites in aqueous medium with humus substances. J. Spectrosc. 2013, 538608, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Steffen, K.T.; Hatakka, A.; Hofrichter, M. Degradation of humic acids by the litter-decomposing basidiomycete Collybia dryophila. Appl. Environ. Microbiol. 2002, 68, 3442–3448. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, O.V.; Kulikova, N.A.; Alekseeva, T.N.; Stepanova, E.V.; Davidchik, V.N.; Beliaeva, E.I.; Tsvetkova, E.A. A comparative characterization of fungal melanin and the humin-like substances synthesized by Cerrena maxima 0275. Prikl. Biokhim. Mikrobiol. 2007, 43, 69–76. [Google Scholar] [CrossRef]
- Ghosh, N.; Ganguly, S.; Banik, A.K. Comparison of microbial humic acid produced by Aspergillus Niger x1 and soil humic acid. J. Pharma Bio Sci. 2011, 2, 478–483. [Google Scholar]
- Kontchou, C.Y.; Blondeau, R. Effect of heterotrophic bacteria on different humic substances in mixed batch cultures. Can. J. Soil Sci. 1990, 70, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Rocker, D.; Kisand, V.; Scholz-Böttcher, B.; Kneib, T.; Lemke, A.; Rullkötter, J.; Simon, M. Differential decomposition of humic acids by marine and estuarine bacterial communities at varying salinities. Biogeochemistry 2012, 111, 331–346. [Google Scholar] [CrossRef]
- Valero, N.O.; Gómez, L.C.; Melgarejo, L.M. Supramolecular characterization of humic acids obtained through the bacterial transformation of a low rank coal. J. Braz. Chem. Soc. 2018, 29, 1842–1853. [Google Scholar] [CrossRef]
- Sposito, G. The Chemistry of Soils; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Piccolo, A.; Spaccini, R.; Drosos, M.; Vinci, G.; Cozzolino, V. The molecular composition of humus carbon: Recalcitrance and reactivity in soils. In The Future of Soil Carbon; Garcia, C., Nannipieri, P., Hernandez, T., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 87–124. [Google Scholar]
- Drosos, M.; Leenheer, J.A.; Avgeropoulos, A.; Deligiannakis, Y. H-binding of size- and polarity-fractionated soil and lignite humic acids after removal of metal and ash components. Environ. Sci. Pollut. R 2014, 21, 3963–3971. [Google Scholar] [CrossRef]
- Galantini, J.; Rosell, R. Long-term fertilization effects on soil organic matter quality and dynamics under different production systems in semiarid soils. Soil Till. Res. 2006, 87, 72–79. [Google Scholar] [CrossRef]
- See, J.; Bronk, D. Changes in C:N ratios and chemical structures of estuarine humic substances during aging. Mar. Chem. 2005, 97, 334–346. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; An, T.; Wei, D.; Chi, F.; Zhou, B. Effects of long-term fertilization on soil humic acid composition and structure in Black Soil. PLoS ONE 2017, 12, e0186918. [Google Scholar] [CrossRef] [Green Version]
- Metz, B.; Davidson, O.R.; Bosch, P.R.; Dave, R.; Meyer, L.A. (Eds.) Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Schoebitz, M.; López, M.D.; Serrí, H.; Martínez, O.; Zagal, E. Combined application of microbial consortium and humic substances to improve the growth performance of blueberry seedlings. J. Soil Sci. Plant Nutr. 2016, 16, 1010–1023. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Sas-Paszt, L.; Frąc, M. The status of soil microbiome as affected by the application of phosphorus biofertilizer: Fertilizer enriched with beneficial bacterial strains. Int. J. Mol. Sci. 2020, 21, 8003. [Google Scholar] [CrossRef]
- Pertile, G.; Lamorski, K.; Bieganowski, A.; Boguta, P.; Brzezińska, M.; Polakowski, C.; Skic, K.; Sokołowska, Z.; Baranowski, P.; Gackiewicz, B.; et al. Immediate effects of the application of various fungal strains with urea fertiliser on microbiome structure and functions and their relationships with the physicochemical parameters of two different soil types. Appl. Soil Ecol. 2021, 163, 103972. [Google Scholar] [CrossRef]
- Ghabbour, E.A.; Davies, G.; Misiewicz, T.; Alami, R.A.; Askounis, E.M.; Cuozzo, N.P.; Filice, A.J.; Haskell, J.M.; Moy, A.K.; Roach, A.C.; et al. Chapter one—National comparison of the total and sequestered organic matter contents of conventional and organic farm soils. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 146, pp. 1–35. [Google Scholar]
- Cybulak, M.; Sokołowska, Z.; Boguta, P.; Tomczyk, A. Influence of pH and grain size on physicochemical properties of biochar and released humic substances. Fuel 2019, 240, 334–338. [Google Scholar] [CrossRef]
- Ohno, T. Fluorescence inner-filtering correction for determining the humification index of dissolved organic matter. Environ. Sci. Technol. 2002, 36, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Boguta, P.; Sokołowska, Z. Statistical relationship between selected physicochemical properties of peaty-muck soils and their fraction of humic acids. Int. Agrophys. 2014, 28, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.Q.; Hanna, J.; Johnson, W. Source indicators of humic substances: An elemental composition, solid state 13C CP/MAS NMR and Py-GC/MS study. Appl. Geochem. 2000, 15, 1019–1033. [Google Scholar] [CrossRef]
- Goveia, D.; Melo, C.A.; Oliveira, L.K.; Fraceto, L.F.; Rocha, J.C.; Dias, F.; Newton, L.; Rosa, A.H. Adsorption and release of micronutrients by humin extracted from peat samples. J. Braz. Chem. Soc. 2013, 24, 721–730. [Google Scholar] [CrossRef]
- Canellas, L.P.; Façanha, A.R. Chemical nature of soil humified fractions and their bioactivity. Pesqui. Agropecu. Bras. 2004, 39, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Ding, G.; Mao, J.; Xing, B. Characteristics of amino acids in soil humic substances. Commun. Soil Sci. Plant Anal. 2001, 32, 1991–2005. [Google Scholar] [CrossRef]
- Niemiałkowska-Butrym, I.; Talarowska, A.; Sokołowska, Z.; Boguta, P. Optical properties of humic acids in selected organic wastes. Acta Agrophys. 2012, 19, 773–785. [Google Scholar]
- Van Krevelen, D.W. Graphical-Statistical Method for the Study of Structure and Reaction Processes of Coal. Fuel 1950, 29, 269–284. [Google Scholar]
- Pegoraro, R.F.; Moreira, C.G.; Dias, D.G.; Silveira, T.C. Carbon and nitrogen stocks in the soil and humic substances ofagricultural crops in the semi-arid region. Rev. Ciênc. Agrôn. 2018, 49, 574–583. [Google Scholar]
- Tan, K.H. Humic Matter in Soil and the Environment: Principles and Controversies; Marcel Dekker: New York, NY, USA, 2003. [Google Scholar]
- Kumada, K. Elementary composition of humic acids. Soil Sci. Plant Nutr. 1955, 1, 75–76. [Google Scholar] [CrossRef]
- Baddi, G.A.; Hafidi, M.; Gilard, V.; Revel, J.C. Characterization of humic acids produced during composting of olive mill wastes: Elemental and spectroscopic analyses (FTIR and 13C-NMR). Agron. EDP Sci. 2003, 23, 661–666. [Google Scholar]
- Wang, S.; Wang, N.; Xu, J.; Zhang, X.; Dou, S. Contribution of microbial residues obtained from lignin and cellulose on humus formation. Sustainability 2019, 11, 4777. [Google Scholar] [CrossRef] [Green Version]
- Baranciková, G.; Senesi, N.; Brunetti, G. Chemical and spectroscopic characterization of humic acids isolated from different Slovak soil types. Geoderma 1997, 78, 251–266. [Google Scholar] [CrossRef]
- Boguta, P.; Pieczywek, P.M.; Sokołowska, Z. A comparative study of the application of fluorescence excitation-emission matrices combined with parallel factor analysis and nonnegative matrix factorization in the analysis of Zn complexation by humic acids. Sensors 2016, 16, 1760. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Yu, H.; Yang, F.; Liu, L.; Gao, H.; Cui, B. Characterizing humic substances from native halophyte soils by fluorescence spectroscopy combined with parallel factor analysis and canonical correlation analysis. Sustainability 2020, 12, 9787. [Google Scholar] [CrossRef]
- Valencia, S.; Marín, J.; Restrepo, G.; Frimmel, F. Application of excitation-emission fluorescence matrices and UV/Vis absorption to monitoring the photocatalytic degradation of commercial humic acid. Sci. Total Environ. 2013, 442, 207–214. [Google Scholar] [CrossRef]
- Wu, F.; Midorikawa, T.; Tanoue, E. Fluorescence properties of organic ligands for copper(II) in Lake Biwa and its rivers. Geochem. J. 2001, 35, 333–346. [Google Scholar] [CrossRef]
- Coble, P.G. Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy. Mar. Chem. 1996, 51, 325–346. [Google Scholar] [CrossRef]
- Gao, J.F.; Dou, S.; Wang, Z.G. Structural analysis of humic acid in soil at different corn straw returning modes through fluorescence spectroscopy and infrared spectroscopy. Int. J. Anal. Chem. 2019, 2019, 1086324. [Google Scholar] [CrossRef] [Green Version]
- Matthews, B.J.H.; Jones, A.C.; Theodorou, N.K.; Tudhope, A.W. Excitation-emission-matrix fluorescence spectroscopy applied to humic acid bands in coral reefs. Mar. Chem. 1996, 55, 317–332. [Google Scholar] [CrossRef]
- Hernandez, D.; Plaza, C.; Senesi, N.; Polo, A. Detection of Copper(II) and zinc(II) binding to humic acids from pig slurry and amended soils by fluorescence spectroscopy. Environ. Pollut. 2006, 143, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Plaza, C.; Brunetti, G.; Senesi, N.; Polo, A. Molecular and quantitative analysis of metal ion binding to humic acids from sewage sludge and sludge-amended soils by fluorescence spectroscopy. Environ. Sci. Technol. 2006, 40, 917–923. [Google Scholar] [CrossRef]
- Marinari, S.; Dell’Abate, M.T.; Brunetti, G.; Dazzi, C. Differences of stabilized organic carbon fractions and microbiological activity along Mediterranean Vertisols and Alfisols profiles. Geoderma 2010, 156, 379–388. [Google Scholar] [CrossRef]
- Kakde, U.B.; Jamdhade, V.M. Studies on fungi responcible for biodegradation and humification of organic matter. Bionano Front. 2009, 2, 77–81. [Google Scholar]
- Guggenberger, G. Humification and mineralization in soils. In Microorganisms in Soils: Roles in Genesis and Functions; Buscot, F., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2005; pp. 85–106. [Google Scholar]
- Sollins, P.; Homann, P.; Caldwell, B.A. Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma 1996, 74, 65–105. [Google Scholar] [CrossRef]
- Karavanova, E.I. Dissolved organic matter: Fractional composition and sorbability by the soil solid phase (review of literature). Eurasian Soil Sci. 2013, 46, 833–844. [Google Scholar] [CrossRef]
- Kluczek-Turpeinen, B.; Steffen, K.T.; Tuomela, M.; Hatakka, A.; Hofrichter, M. Modification of humic acids by the compost dwelling deuteromycete Paecilomyces inflatus. Appl. Microbiol. Biotechnol. 2005, 66, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Ghosh, R.; Banerjee, D.; Nath, P.; Mazumder, U.K.; Ghosal, S. Biotransformation of 3-hydroxydibenzo-α-pyrone into 3,8 dihydroxydibenzo-α-pyrone and aminoacyl conjugates by Aspergillus niger isolated from native“shilajit”. Electron. J. Biotechnol. 2008, 11, 1–10. [Google Scholar]
- Heidarzadeh, M.H.; Amani, H.; Javadian, B. Improving municipal solid waste compost process by cycle time reduction through inoculation of Aspergillus niger. J. Environ. Health Sci. Eng. 2019, 17, 295–303. [Google Scholar] [CrossRef]
- Ghervase, L.; Ioja, C.; Carstea, E.M.; Niculita, L.; Savastru, D.; Pavelescu, G.; Vanau, G. Evaluation of lentic ecosystems from Bucharest City. Int. J. Energy Environ. Econ. 2011, 5, 183–192. [Google Scholar]
- Yadav, M.K.; Aryal, R.; Short, M.D.; Saint, C.P. Fluorescence excitation-emission spectroscopy: An analytical technique to monitor drugs of addiction in wastewater. Water 2019, 11, 377. [Google Scholar] [CrossRef] [Green Version]
- Boguta, P.; Sokołowska, Z. Zinc binding to fulvic acids: Assessing the impact of pH, metal concentrations and chemical properties of fulvic acids on the mechanism and stability of formed soluble complexes. Molecules 2020, 25, 1297. [Google Scholar] [CrossRef] [Green Version]
- Esham, E.C.; Ye, W.; Moran, M.A. Identification and characterization of humic substances-degrading bacterial isolates from an estuarine environment. FEMS Microbiol. Ecol. 2000, 34, 103–111. [Google Scholar] [CrossRef]
- Walkiewicz, A.; Brzezińska, M.; Bieganowski, A.; Sas-Paszt, L.; Frąc, M. Early response of soil microbial biomass and activity to biofertilizer application in degraded brunic arenosol and abruptic luvisol of contrasting textures. Agronomy 2020, 10, 1347. [Google Scholar] [CrossRef]
- Borowik, K.; Rusek, P.; Schab, S.; Rusek, Ł. Biofertilizer and Method of Its Production (Bionawóz i Sposób Jego Wytwarzania). U.S. Patent D431350, 10 March 2019. [Google Scholar]
- Swift, R.S. Organic matter characterization. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loepper, R.H., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; Chapter 35; pp. 1018–1020. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HAs (%) | E4/E6 | ΔlgK | E2/E6 | HIX | C (%) | H (%) | N (%) | S (%) | O (%) | O/H | O/C | C/N | H/C | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AL | 0.30 | 6.12 | 0.74 | 38.2 | 21.7 | 50.7 | 5.45 | 5.02 | 1.40 | 37.5 | 0.43 | 0.55 | 11.8 | 1.29 |
| ±0.02 | ±0.85 | ±0.03 | ±6.65 | ±1.13 | ±0.31 | ±0.07 | ±0.09 | ±0.03 | ±0.37 | ±0.01 | ±0.01 | ±0.20 | ±0.02 | |
| BA | 0.29 | 6.43 | 0.72 | 36.7 | 23.3 | 51.8 | 5.44 | 4.54 | 1.12 | 37.1 | 0.43 | 0.54 | 13.3 | 1.26 |
| ±0.02 | ±0.55 | ±0.02 | ±4.00 | ±2.12 | ±0.71 | ±0.08 | ±0.08 | ±0.04 | ±0.75 | ±0.01 | ±0.02 | ±0.30 | ±0.01 |
| Influencing Factor (Bold) | Compared to: | C/N | H/C | Humification as FIα/γ | Humification as HIX | Humification as ΔlogK | Humification as O/C | O/H |
|---|---|---|---|---|---|---|---|---|
| AL (S + F) | AL (S) | - | - | ↓ | ↓ | ↓ | - | ↓ |
| AL (S + B) | AL (S) | - | - | ↓ | ↓ | ↓ | - | - |
| AL (S + NPK + F) | AL (S + NPK) | - | - | - | ↓* | - | ↓ | ↑ |
| AL (S + NPK + B) | AL (S + NPK) | - | - | - | - | ↑ | - | - |
| AL (S + NPK) | AL (S) | ↓ | - | ↓ | ↓ | - | ↓* | ↑* |
| BA (S + F) | BA (S) | - | - | - | - | ↑* | ↑* | ↓ |
| BA (S + B) | BA (S) | - | - | - | - | - | - | - |
| BA (S + NPK + F) | BA (S + NPK) | - | - | ↑* | - | - | - | - |
| BA (S + NPK + B) | BA (S + NPK) | - | - | - | ↓ | - | - | - |
| BA (S + NPK) | BA (S) | - | - | ↑* | - | - | ↓* | ↑ |
| % HAs in AL | % HAs in BA (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 | |
| S | 0.33 c ± 0.011 | 0.35 d ± 0.017 | 0.27 a ± 0.026 | 0.30 b ± 0.013 | 0.30 b ± 0.017 | 0.29 b ± 0.024 | 0.15 a ± 0.024 | 0.31 b ± 0.025 |
| S + F | 0.30 b ± 0.007 | 0.36 c ± 0.024 | 0.26 a ± 0.019 | 0.27 a ± 0.018 | 0.33 b ± 0.008 | 0.32 b ± 0.021 | 0.24 a ± 0.035 | 0.32 b ± 0.019 |
| S + B | 0.29 c ± 0.013 | 0.32 d ± 0.009 | 0.16 a ± 0.023 | 0.27 b ± 0.013 | 0.28 b ± 0.011 | 0.29 b ± 0.025 | 0.22 a ± 0.026 | 0.29 b ± 0.013 |
| S + NPK | 0.30 b ± 0.015 | 0.31 c ± 0.014 | 0.28 a ± 0.006 | 0.38 d ± 0.009 | 0.30 c ± 0.015 | 0.28 b ± 0.023 | 0.21 a ± 0.025 | 0.29 bc ± 0.019 |
| S + NPK + F | 0.27 b ± 0.012 | 0.29 c ± 0.013 | 0.23 a ± 0.036 | 0.32 d ± 0.021 | 0.29 c ± 0.011 | 0.24 b ± 0.018 | 0.23 a ± 0.002 | 0.29 c ± 0.020 |
| S + NPK + B | 0.31 bc ± 0.007 | 0.30 b ± 0.019 | 0.27 a ± 0.040 | 0.32 c ± 0.013 | 0.27 b ± 0.012 | 0.27 b ± 0.016 | 0.23 a ± 0.016 | 0.32 c ± 0.010 |
| Variant | Description | Symbol | AL Soil (Doses per 1 ha) | BA Soil (Doses per 1 ha) |
|---|---|---|---|---|
| I | Soil without NPK fertilizers | S | No additives | No additives |
| II | Soil without NPK fertilizers, treated with fungal strains | S + F | 30.5 kg (17 + 7.5 + 6) fungal strains | 32 kg (17 + 8 + 7) fungal strains |
| III | Soil without NPK fertilizers, treated with bacterial strains | S + B | 14 kg bacterial strains | 14 kg bacterial strains |
| IV | Soil with NPK fertilizers | S + NPK | 300 kg Polifoska 6 (18 kg N + 60 kg P2O5 + 90 kg K2O) 135 kg potassium salt (81 kg K2O) 442 kg (295 + 147) ammonium nitrate (100 + 50 kg N) | 250 kg Polifoska 6, (15 kg N + 50 kg P2O5 + 75 kg K2O) 162 kg potassium salt (97 kg K2O), 295 + 147 kg ammonium nitrate (100 + 50 kg N) |
| V | Soil with NPK fertilizers and fungal strains | S + NPK + F | 300 kg Polifoska 6 (18 kg N + 60 kg P2O5 + 90 kg K2O) 135 kg potassium salt (81 kg K2O) 30.5 kg (17 + 7.5 + 6) fungal strains 442 kg (295 + 147) ammonium nitrate (100 + 50 kg N) | 250 kg Polifoska 6 (15 kg N + 50 kg P2O5 + 75 kg K2O) 162 kg potassium salt (97 kg K2O) 32 kg (17 + 8 + 7) fungal strains 442 kg (295 + 147) ammonium nitrate (100 + 50 kg N) |
| VI | Soil with NPK fertilizers and bacterial strains | S + NPK + B | 300 kg Polifoska 6 (18 kg N + 60 kg P2O5 + 90 kg K2O) 135 kg potassium salt (81 kg K2O) 14 kg bacterial strains 442 kg (295 + 147) ammonium nitrate (100 + 50 kg N) | 250 kg Polifoska 6 (15 kg N + 50 kg P2O5 + 75 kg K2O) 162 kg potassium salt (97 kg K2O) 14 kg bacterial strains 442 kg (295 + 147) ammonium nitrate (100 + 50 kg N) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boguta, P.; Skic, K.; Sokołowska, Z.; Frąc, M.; Sas-Paszt, L. Chemical Transformation of Humic Acid Molecules under the Influence of Mineral, Fungal and Bacterial Fertilization in the Context of the Agricultural Use of Degraded Soils. Molecules 2021, 26, 4921. https://doi.org/10.3390/molecules26164921
Boguta P, Skic K, Sokołowska Z, Frąc M, Sas-Paszt L. Chemical Transformation of Humic Acid Molecules under the Influence of Mineral, Fungal and Bacterial Fertilization in the Context of the Agricultural Use of Degraded Soils. Molecules. 2021; 26(16):4921. https://doi.org/10.3390/molecules26164921
Chicago/Turabian StyleBoguta, Patrycja, Kamil Skic, Zofia Sokołowska, Magdalena Frąc, and Lidia Sas-Paszt. 2021. "Chemical Transformation of Humic Acid Molecules under the Influence of Mineral, Fungal and Bacterial Fertilization in the Context of the Agricultural Use of Degraded Soils" Molecules 26, no. 16: 4921. https://doi.org/10.3390/molecules26164921