
Antifungal Activities of cis-trans Citral Isomers against Trichophyton rubrum with ERG6 as a Potential Target



Abstract
:
1. Introduction
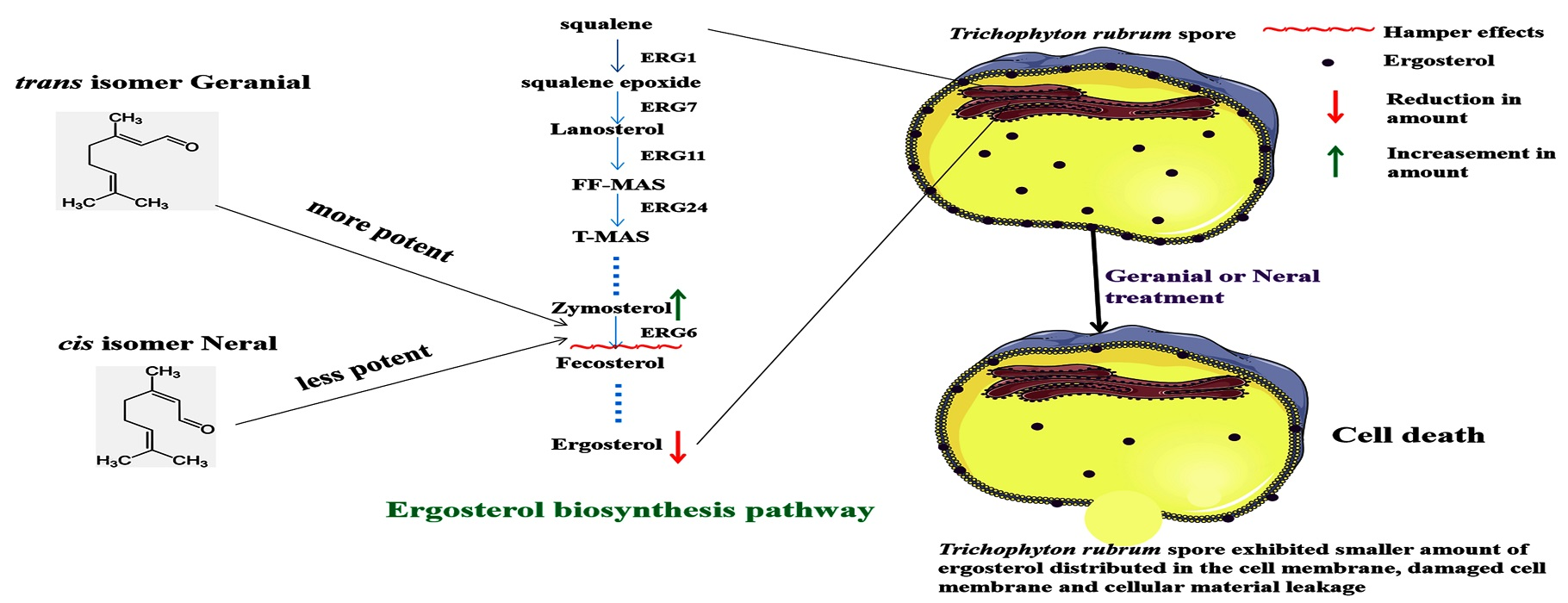
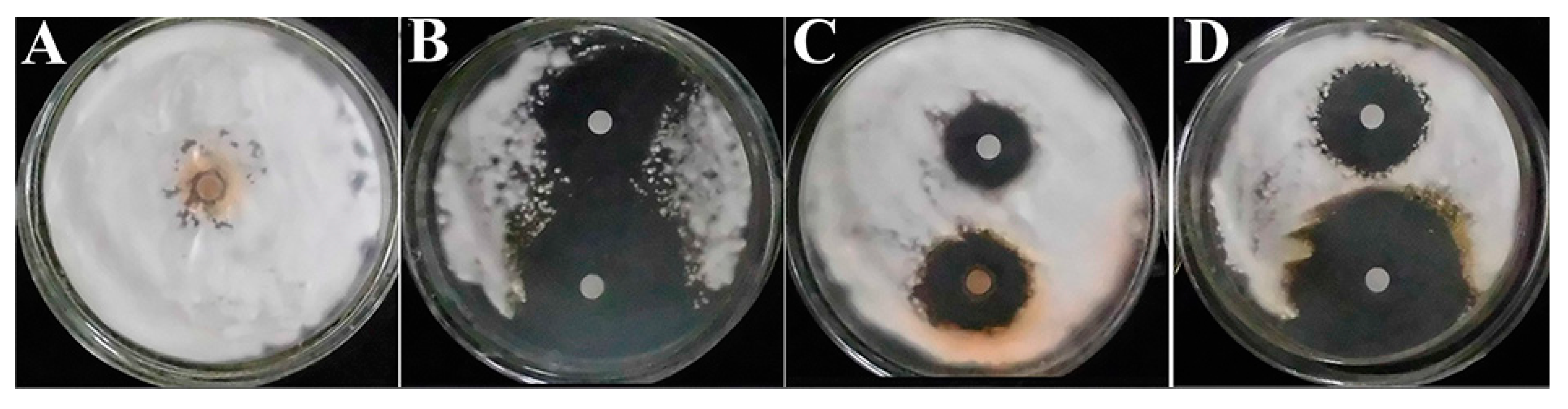
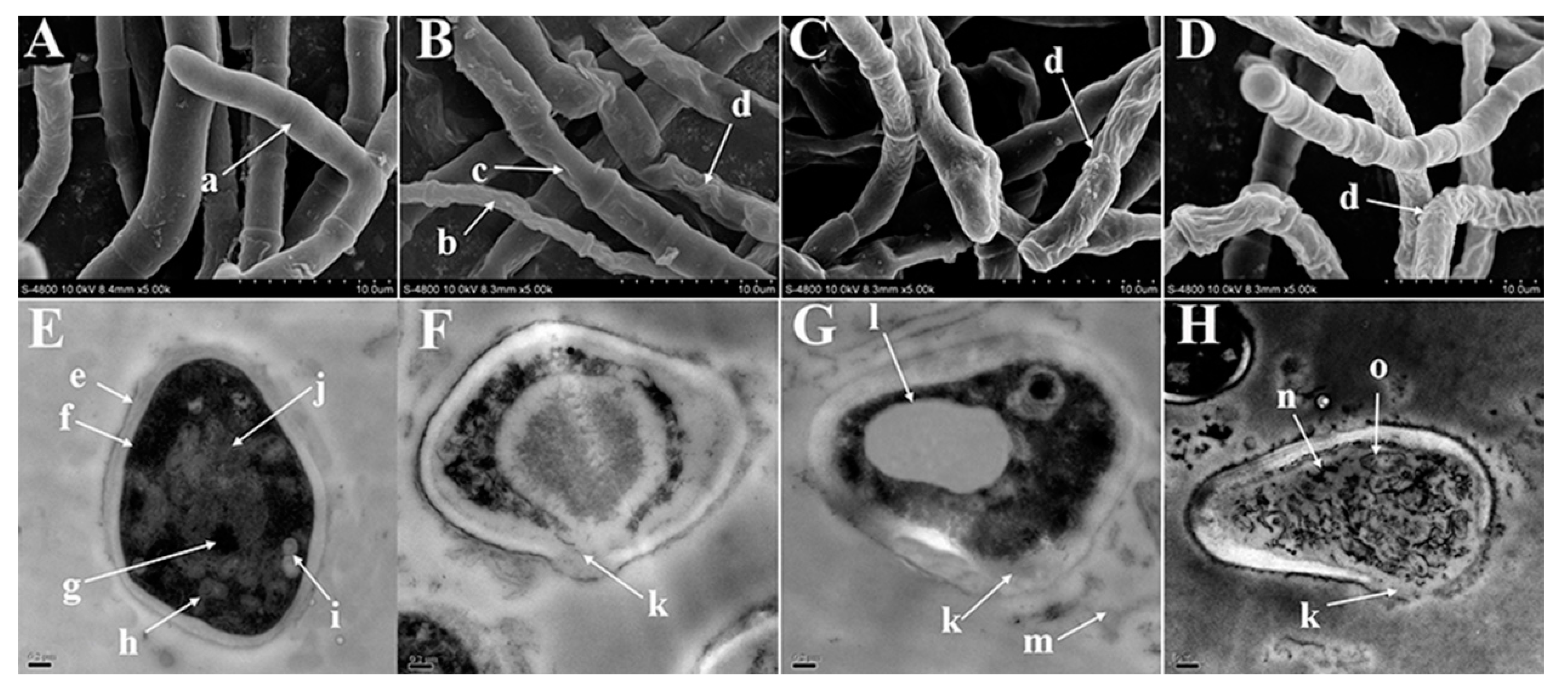
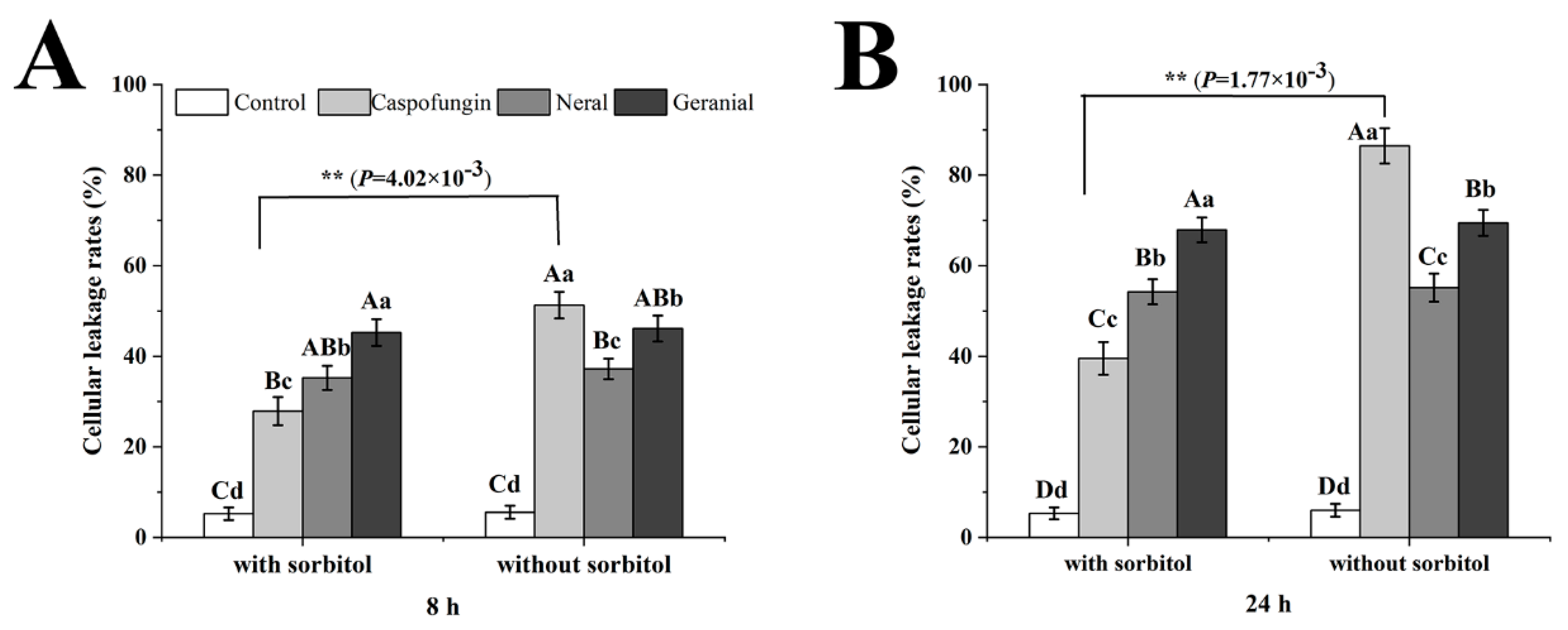
2. Results and Discussion
3. Materials and Methods
3.1. Test Strain and Chemical Compounds
3.2. Minimal Inhibitory Concentrations (MICs)/Minimal Fungicidal Concentrations (MFCs) Determination
3.3. Interaction of Neral with Geranial
3.4. Disc Diffusion Assay
3.5. SEM/TEM Observations
3.6. Cellular Leakage Measurement
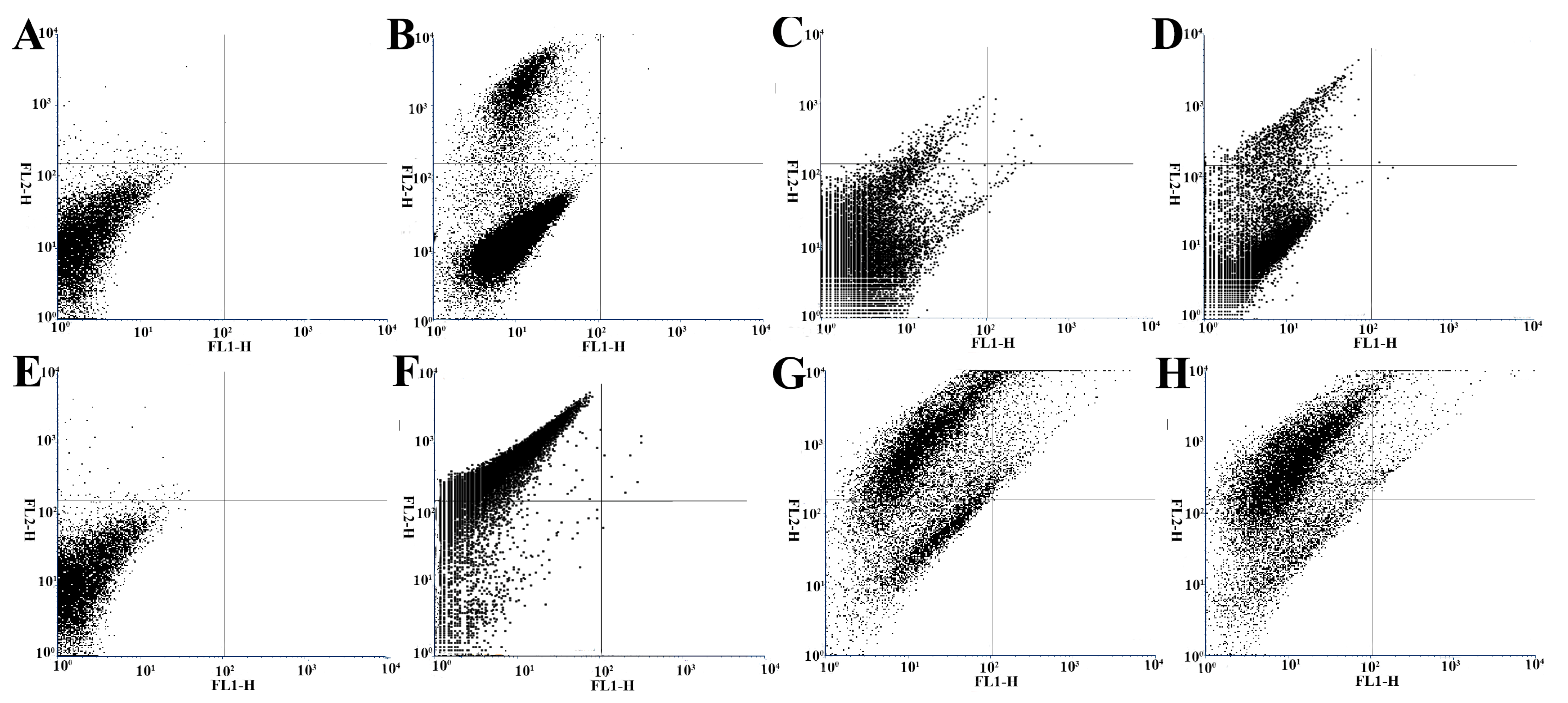
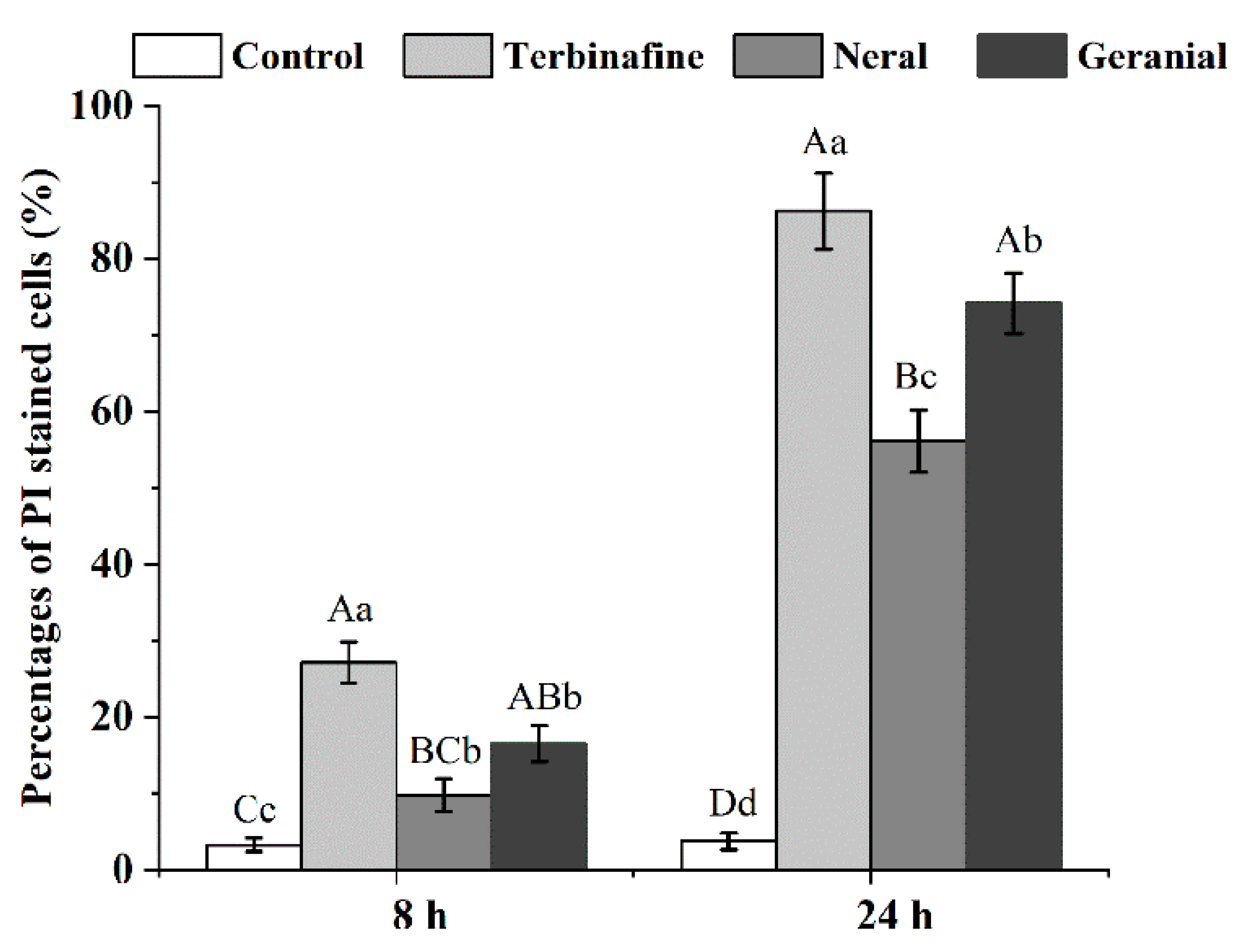
3.7. Flow Cytometry
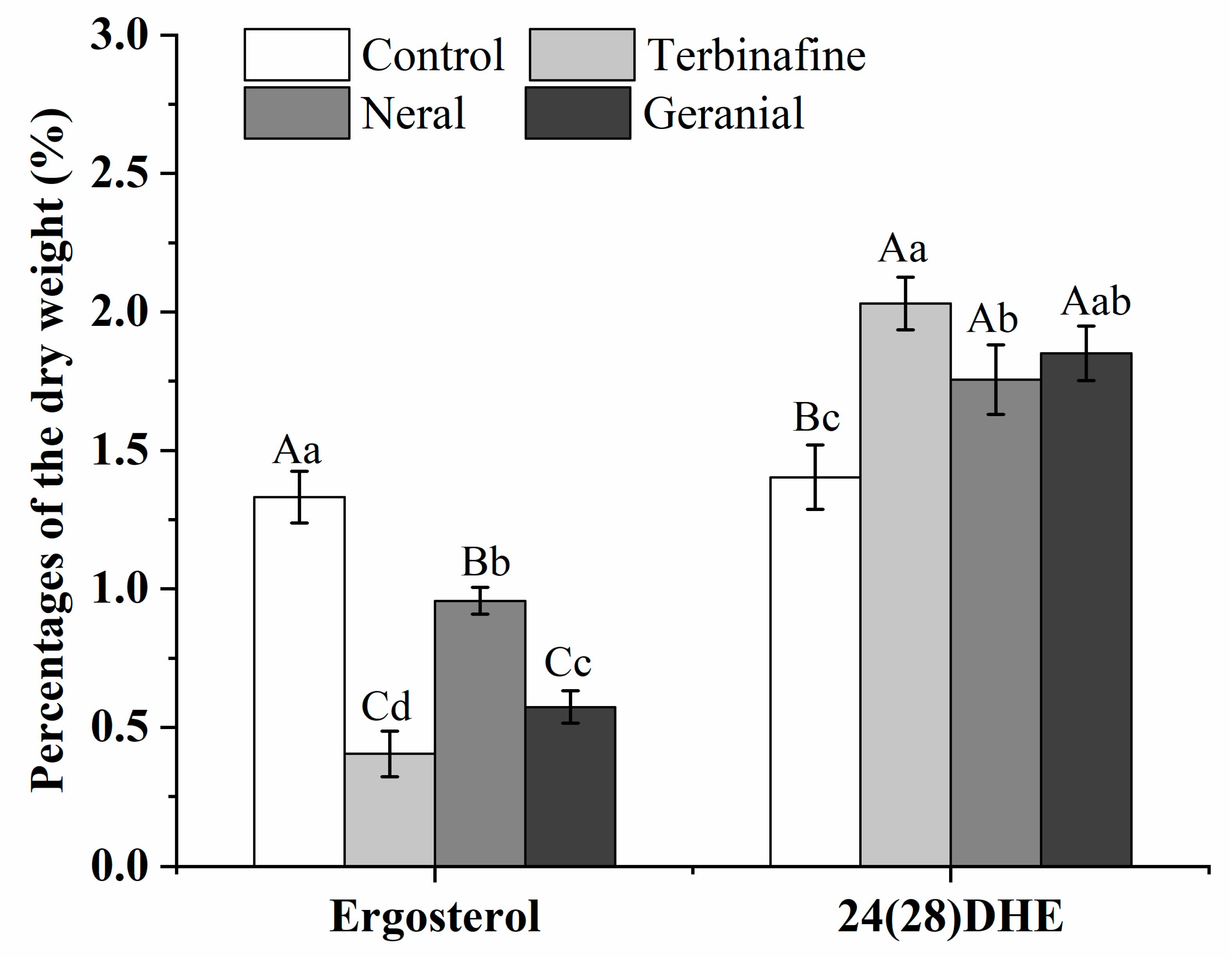
3.8. Sterol Quantitation Test
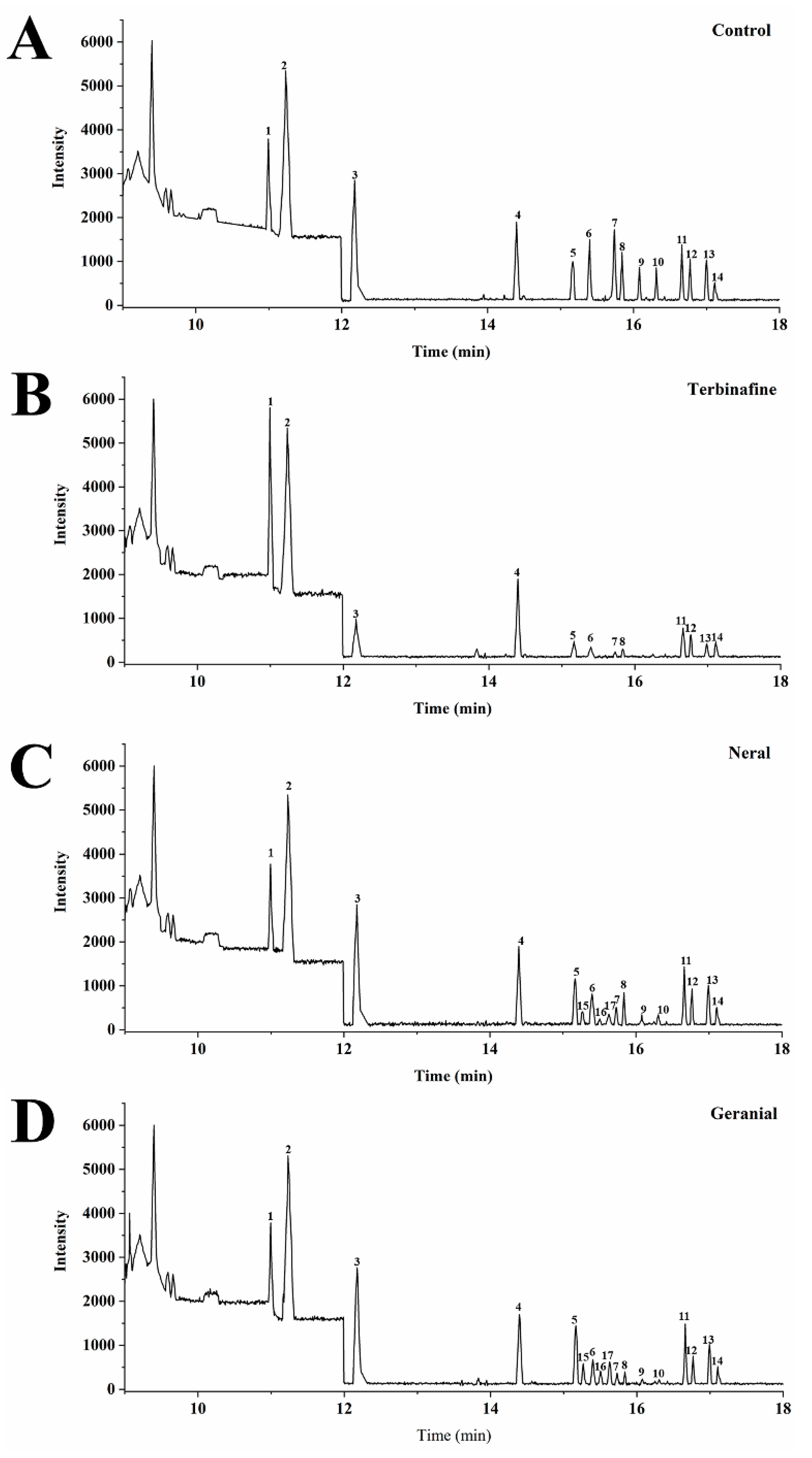
3.9. Sterols Pattern Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gräser, Y.; Monod, M.; Bouchara, J.-P.; Dukik, K.; Nenoff, P.; Kargl, A.; Kupsch, C.; Zhan, P.; Packeu, A.; Chaturvedi, V.; et al. New insights in dermatophyte research. Med. Mycol. 2018, 56, S2–S9. [Google Scholar] [CrossRef] [Green Version]
- Scher, R.K.; Nakamura, N.; Tavakkol, A. Luliconazole: A review of a new antifungal agent for the topical treatment of onychomycosis. Mycoses 2014, 57, 389–393. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Dogra, S.; Shaw, D. Antifungal drug susceptibility testing of dermatophytes: Laboratory findings to clinical implications. Indian Dermatol. Online J. 2019, 10, 225–233. [Google Scholar] [CrossRef]
- Lopes, G.; Pinto, E.; Salgueiro, L. Natural Products: An Alternative to Conventional Therapy for Dermatophytosis? Mycopathologia 2017, 182, 143–167. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Lim, S. Antifungal effects of herbal essential oils alone and in combination with ketoconazole against Trichophyton spp. J. Appl. Microbiol. 2004, 97, 1289–1296. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Ocimum sanctum essential oil and its active principles exert their antifungal activity by disrupting ergosterol biosynthesis and membrane integrity. Res. Microbiol. 2010, 161, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Gwak, K.; Yang, I.; Kim, K.; Jeung, E.; Chang, J.; Choi, I. Effect of citral, eugenol, nerolidol and α-terpineol on the ultrastructural changes of Trichophyton mentagrophytes. Fitoterapia 2009, 80, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Uchida, K.; Abe, S. Vapor activity of 72 essential oils against a Trichophyton mentagrophytes. J. Infect. Chemother. 2006, 12, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.C.A.; Bezerra, A.P.D.B.; de Sousa, J.P.; Guerra, F.Q.S.; Lima, E.D.O. Evaluation of Antifungal Activity and Mechanism of Action of Citral againstCandida albicans. Evid. Based Complement. Altern. Med. 2014, 2014, 378280. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.D.B.D.; Guterres, S.; Weisheimer, V.; Schapoval, E. Antifungal activity of the lemongrass oil and citral against Candida spp. Braz. J. Infect. Dis. 2008, 12, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 2014, 153, 265–271. [Google Scholar] [CrossRef]
- Zheng, S.; Jing, G.; Wang, X.; Ouyang, Q.; Jia, L.; Tao, N. Citral exerts its antifungal activity against Penicillium digitatum by affecting the mitochondrial morphology and function. Food Chem. 2015, 178, 76–81. [Google Scholar] [CrossRef]
- Li, R.-Y.; Wu, X.-M.; Yin, X.-H.; Liang, J.-N.; Li, M. The Natural Product Citral Can Cause Significant Damage to the Hyphal Cell Walls of Magnaporthe grisea. Molecules 2014, 19, 10279–10290. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Lin, L.; Jiang, L.; Huang, Y. Effect of cis-trans Isomer of Citral on Microstructure and Membrane Function of As-pergillus flavus. Chin. J. Biochem. Mol. Biol. 2007, 23, 770–778. (In Chinese). [Google Scholar] [CrossRef]
- López-Oviedo, E.; Aller, A.I.; Martín, C.; Castro, C.; Ramirez, M.; Pemán, J.M.; Cantón, E.; Almeida-González, C.V.; Martín-Mazuelos, E. Evaluation of Disk Diffusion Method for Determining Posaconazole Susceptibility of Filamentous Fungi: Comparison with CLSI Broth Microdilution Method. Antimicrob. Agents Chemother. 2006, 50, 1108–1111. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-J.; Gwak, K.-S.; Yang, I.; Choi, W.-S.; Jo, H.-J.; Chang, J.-W.; Jeung, E.-B.; Choi, I.-G. Antifungal activities of the essential oils in Syzygium aromaticum (L.) Merr. Et Perry and Leptospermum petersonii Bailey and their constituents against various dermatophytes. J. Microbiol. 2007, 45, 460–465. [Google Scholar]
- Voříšek, J.; Pokorný, V. Cytochemical detection of polysaccharides on the surface of the cell membrane complex in fungi. Arch. Microbiol. 1975, 102, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The Mechanism of Antifungal Action of Essential Oil from Dill (Anethum graveolens L.) on Aspergillus flavus. PLoS ONE 2012, 7, e30147. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.J.; Brandt, K.D.; Cugier, D.; Goldman, R. A Whole-Cell Candida albicans Assay for the Detection of Inhibitors towards Fungal Cell Wall Synthesis and Assembly. J. Antibiot. 1995, 48, 306–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina-Vaz, C.; Sansonetty, F.; Rodrigues, A.G.; Costa-Oliveira, S.; Tavares, C.; de Oliveira, J.M. Cytometric approach for a rapid evaluation of susceptibility of Candida strains to antifungals. Clin. Microbiol. Infect. 2001, 7, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.S.A.; Ahmad, I. Antifungal activity of essential oils and their synergy with fluconazole against drug-resistant strains of Aspergillus fumigatus and Trichophyton rubrum. Appl. Microbiol. Biotechnol. 2011, 90, 1083–1094. [Google Scholar] [CrossRef]
- Rodrigues, M.L. The Multifunctional Fungal Ergosterol. mBio 2018, 9, e01755-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.; Binder, U.; Bracher, F.; Giera, M. Antifungal drug testing by combining minimal inhibitory concentration testing with target identification by gas chromatography–mass spectrometry. Nat. Protoc. 2017, 12, 947–963. [Google Scholar] [CrossRef]
- Blechert, O.; Mei, H.; Zang, X.; Zheng, H.; Liang, G.; Liu, W. Auxotrophic mutations of Trichophyton rubrum created by in vitro synthesized Cas9 ribonucleoprotein. BMC Biotechnol. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Thomson, S.; Rice, C.A.; Zhang, T.; Edrada-Ebel, R.; Henriquez, F.; Roberts, C.W. Characterisation of sterol biosynthesis and validation of 14α-demethylase as a drug target in Acanthamoeba. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of Ergosterol Content: Novel Method for Determination of Fluconazole Susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [CrossRef] [Green Version]
- Lone, S.A.; Khan, S.; Ahmad, A. Inhibition of ergosterol synthesis in Candida albicans by novel eugenol tosylate congeners targeting sterol 14α-demethylase (CYP51) enzyme. Arch. Microbiol. 2020, 202, 711–726. [Google Scholar] [CrossRef]
- Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. Approved Standard, 2nd ed; 2008. CLSI Document M38-A2. Available online: https://clsi.org/media/1455/m38a2_sample.pdf (accessed on 13 July 2021).
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Aljabre, S.H.M.; Randhawa, M.A.; Akhtar, N.; Alakloby, O.M.; Alqurashi, A.M.; Aldossary, A. Antidermatophyte activity of ether extract of Nigella sativa and its active principle, thymoquinone. J. Ethnopharmacol. 2005, 101, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Lunde, C.S.; Kubo, I. Effect of Polygodial on the Mitochondrial ATPase of Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2000, 44, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Inhibition Zone (mm) | |
|---|---|---|
| MIC | MFC | |
| Control | 0.0 ± 0.0 dC | 0.0 ± 0.0 cC |
| Terbinafine | 30.2 ± 1.6 aA | 52.4 ± 3.0 aA |
| Neral | 20.4 ± 2.3 cB | 39.6 ± 2.9 bB |
| Geranial | 27.8 ± 2.5 bA | 50.6 ± 3.3 aA |
| Peak No. | Sterols | Control | Terbinafine (MIC) | Neral (MIC) | Geranial (MIC) |
|---|---|---|---|---|---|
| 1 | Squalene | 1 | 2.13 | 0.97 | 0.98 |
| 2 | Cholestane (IS) | 1 | 0.97 | 0.98 | 0.98 |
| 3 | Squalene epoxide | 1 | 0.56 | 0.97 | 0.99 |
| 4 | Cholesterol (IS) | 1 | 0.98 | 0.95 | 0.99 |
| 5 | Zymosterol | 1 | 0.43 | 1.28 | 1.57 |
| 6 | Ergosterol | 1 | 0.21 | 0.49 | 0.35 |
| 7 | Dehydroergosterol | 1 | 0.07 | 0.22 | 0.14 |
| 8 | Fecosterol | 1 | 0.11 | 0.67 | 0.22 |
| 9 | 5-Dehydroergosterol | 1 | - | 0.17 | 0.15 |
| 10 | Episterol | 1 | - | 0.18 | 0.15 |
| 11 | Lanosterol | 1 | 0.61 | 1.11 | 1.21 |
| 12 | 4-Methylfecosterol | 1 | 0.53 | 0.88 | 0.65 |
| 13 | 4,4-Dimethylcholesta-8,24-dien-3β-ol (T-MAS) | 1 | 0.25 | 0.97 | 0.98 |
| 14 | 4,4-Dimethylcholesta-8,14,24-trien-3β-ol (FF-MAS) | 1 | 0.68 | 0.98 | 0.97 |
| 15 | 7-Dehydrodesmosterol | - | - | + | + |
| 16 | Cholesta-7,24-dien-3β-ol | - | - | + | + |
| 17 | Cholesta-5,7,22,24-tetraen-3β-ol | - | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Shang, Y.; Li, M.; Li, Y.; Ouyang, W. Antifungal Activities of cis-trans Citral Isomers against Trichophyton rubrum with ERG6 as a Potential Target. Molecules 2021, 26, 4263. https://doi.org/10.3390/molecules26144263
Zheng Y, Shang Y, Li M, Li Y, Ouyang W. Antifungal Activities of cis-trans Citral Isomers against Trichophyton rubrum with ERG6 as a Potential Target. Molecules. 2021; 26(14):4263. https://doi.org/10.3390/molecules26144263
Chicago/Turabian StyleZheng, Yin, Yanhong Shang, Mengyun Li, Yunzhou Li, and Wuqing Ouyang. 2021. "Antifungal Activities of cis-trans Citral Isomers against Trichophyton rubrum with ERG6 as a Potential Target" Molecules 26, no. 14: 4263. https://doi.org/10.3390/molecules26144263