
IgE-Induced Mast Cell Activation Is Suppressed by Dihydromyricetin through the Inhibition of NF-κB Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of DHM on the Proliferation of KU812 Cells
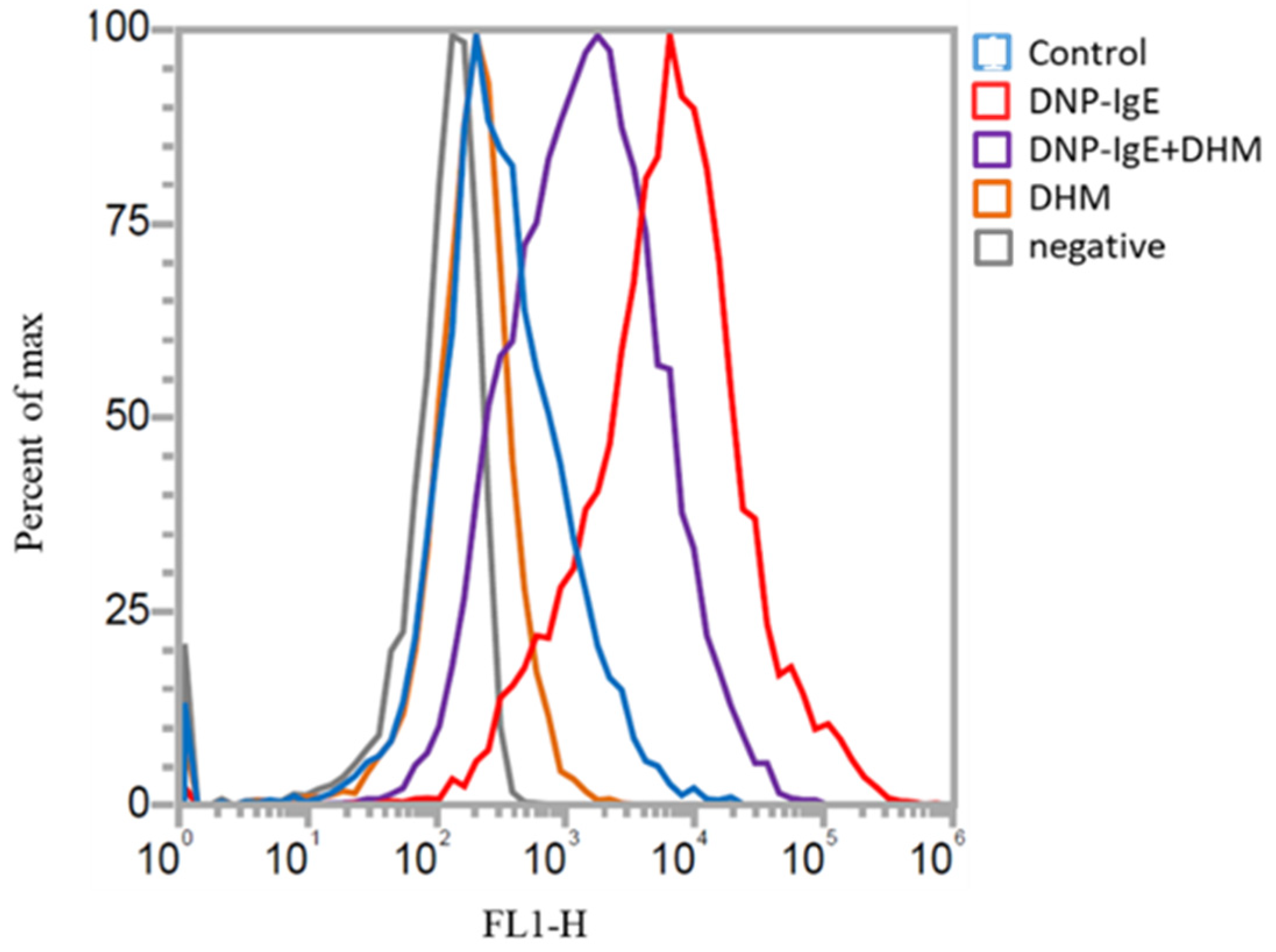
2.2. Inhibitory Effects of DHM on the Generation of Oxidative Stress
2.3. DHM Decreases DNP-IgE-Upregulated Pro-Inflammatory Cytokines and Inhibits Ige-Dependent Activation of the MAPK and NF-κB Signaling Pathways in KU812 Cells
2.4. DHM Promotes Recovery from IgE-Induced Degranulation
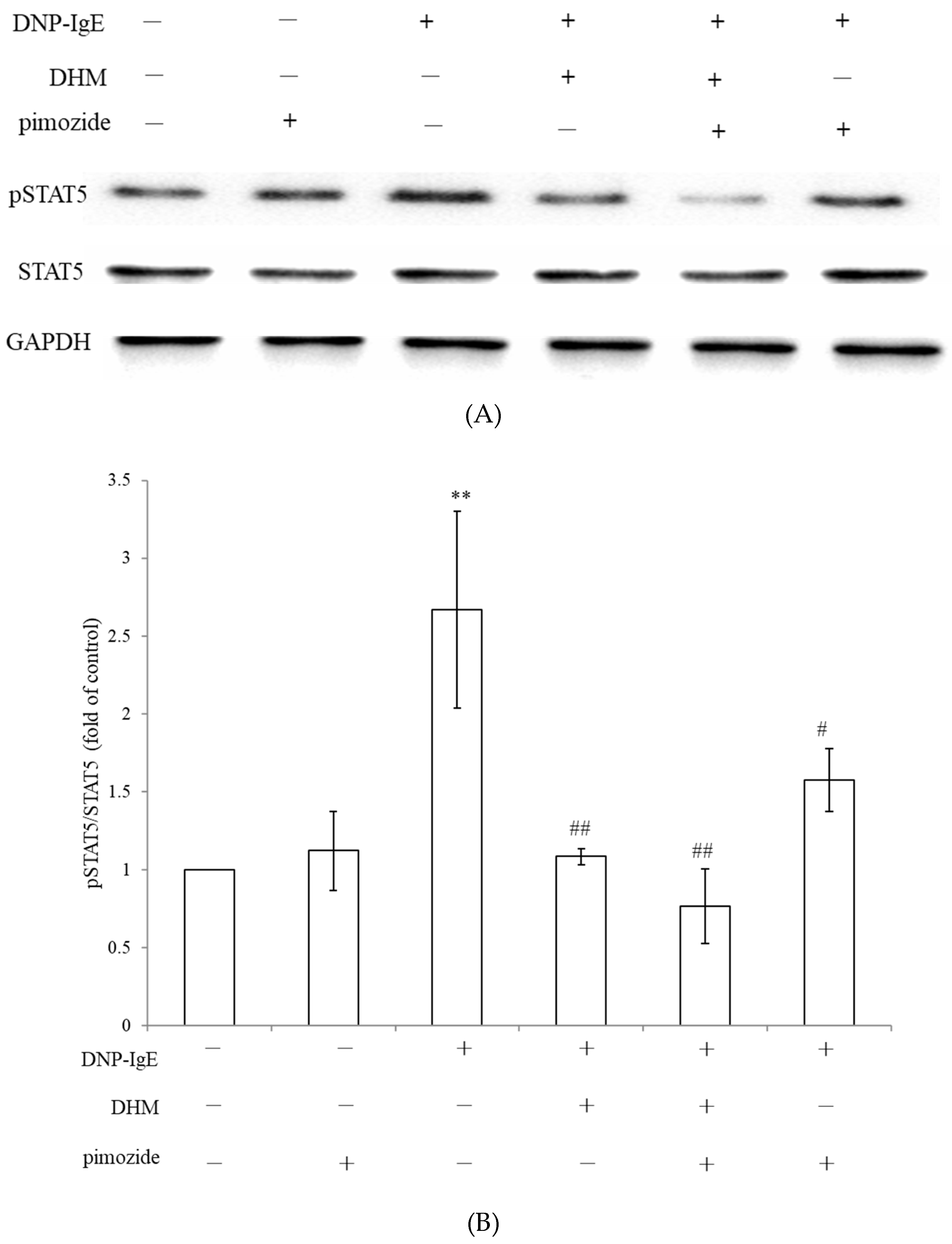
2.5. DHM Inhibited IgE-Mediated Mast Cell Activation through Suppression of STAT5 Signaling Pathway
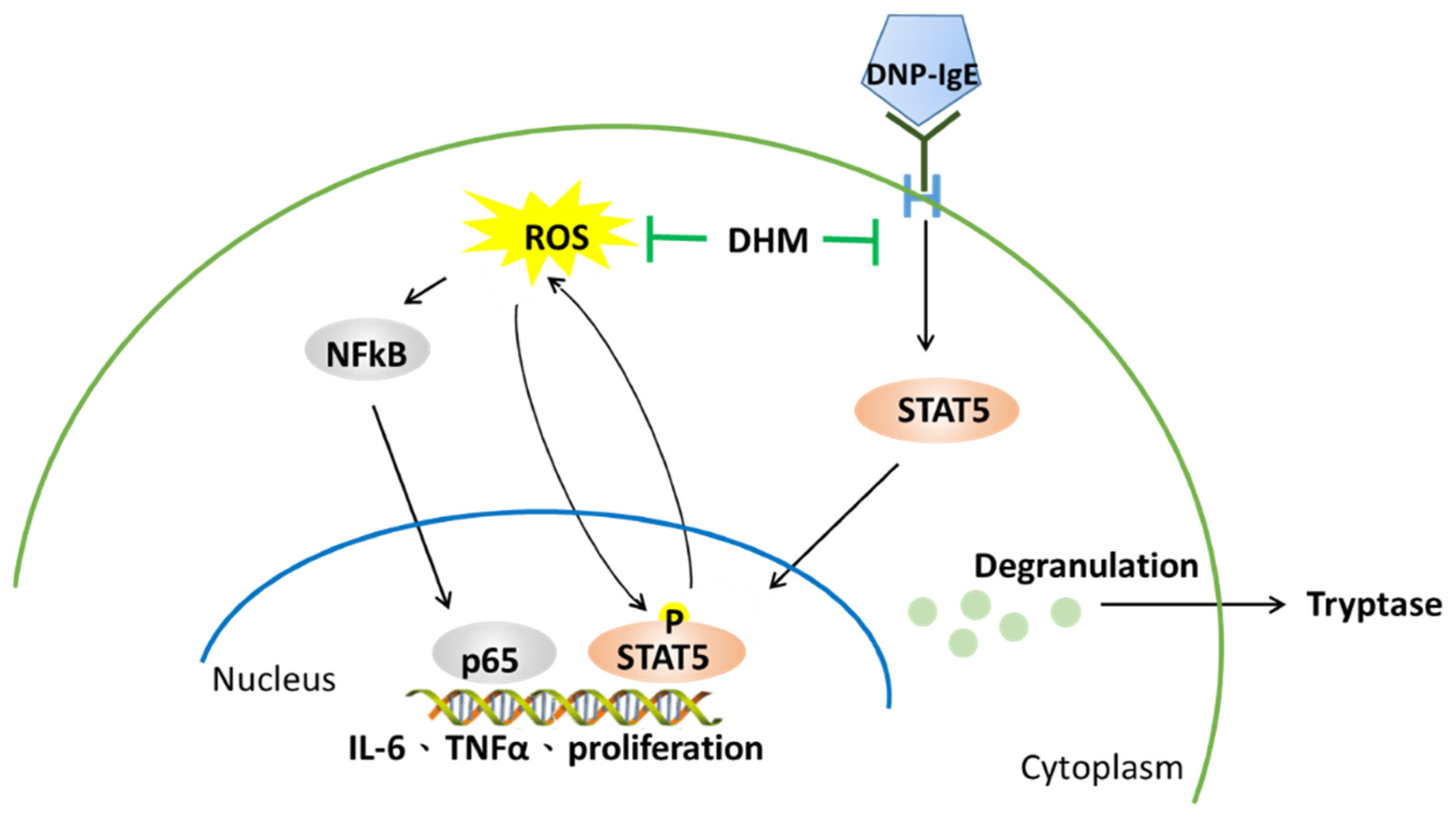
3. Discussion
4. Materials and Methods
4.1. Cell Proliferation Assay
4.2. Cytokines Measurement
4.3. Reactive Oxygen Species (ROS) Measurement Using a Flow Cytometer
4.4. Western Blot Analysis
4.5. Mast Cell Degranulation Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Zhang, J.; Chen, Y.; Luo, H.; Sun, L.; Xu, M.; Yu, J.; Zhou, Q.; Meng, G.; Yang, S. Recent Update on the Pharmacological Effects and Mechanisms of Dihydromyricetin. Front Pharm. 2018, 9, 1204. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhao, X.; Wan, J.; Ran, L.; Qin, Y.; Wang, X.; Gao, Y.; Shu, F.; Zhang, Y.; Liu, P.; et al. Dihydromyricetin improves glucose and lipid metabolism and exerts anti-inflammatory effects in nonalcoholic fatty liver disease: A randomized controlled trial. Pharmacol. Res. 2015, 99, 74–81. [Google Scholar] [CrossRef]
- Chen, T.; Zhu, S.; Lu, Y.; Cao, H.; Zhao, Y.; Jiang, G.; Zhu, L.; Lu, T. Probing the interaction of anti-cancer agent dihydromyricetin with human serum albumin: A typical method study. Anti-Cancer Agents Med. Chem. 2012, 12, 919–928. [Google Scholar] [CrossRef]
- Huang, H.-C.; Liao, C.-C.; Peng, C.-C.; Lim, J.-M.; Siao, J.-H.; Wei, C.-M.; Chen, C.-C.; Wu, C.-S.; Chang, T.-M. Dihydromyricetin from Ampelopsis grossedentata inhibits melanogenesis through down-regulation of MAPK, PKA and PKC signaling pathways. Chem. Biol. Interact. 2016, 258, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Hirano, T.; Higa, S.; Arimitsu, J.; Maruta, M.; Kuwahara, Y.; Ohkawara, T.; Hagihara, K.; Yamadori, T.; Shima, Y.; et al. Flavonoids and Related Compounds as Anti-Allergic Substances. Allergol. Int. 2007, 56, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Gong, J.; Liu, F.; Mohammed, U. Naturally occurring polyphenolic antioxidants modulate IgE-mediated mast cell activation. Immunology 2000, 100, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempuraj, D.; Selvakumar, G.P.; Ahmed, M.E.; Raikwar, S.P.; Thangavel, R.; Khan, A.; Zaheer, S.A.; Iyer, S.S.; Burton, C.; James, D.; et al. COVID-19, Mast Cells, Cytokine Storm, Psychological Stress, and Neuroinflammation. Neuroscientist 2020, 26, 402–414. [Google Scholar] [CrossRef]
- Barnstein, B.O.; Li, G.; Wang, Z.; Kennedy, S.; Chalfant, C.; Nakajima, H.; Bunting, K.D.; Ryan, J.J. Stat5 Expression Is Required for IgE-Mediated Mast Cell Function. J. Immunol. 2006, 177, 3421–3426. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, C.P.; McCoy, M.E.; Piekorz, R.; Sexl, V.; Roh, K.-H.; Jacobs-Helber, S.M.; Gillespie, S.R.; Bailey, D.P.; Mirmonsef, P.; Mann, M.N.; et al. Stat5 expression is critical for mast cell development and survival. Blood 2003, 102, 1290–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullen, N.; Falanga, Y.; Morales, J.; Ryan, J. The Fyn-STAT5 Pathway: A New Frontier in IgE- and IgG-Mediated Mast Cell Signaling. Front. Immunol. 2012, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Tang, N.; Ma, J.; Wang, K.S.; Mi, C.; Lv, Y.; Piao, L.X.; Xu, G.H.; Li, X.; Lee, J.J.; Jin, X. Dihydromyricetin suppresses TNF-α-induced NF-κB activation and target gene expression. Mol. Cell. Biochem. 2016, 422, 11–20. [Google Scholar] [CrossRef]
- Crimi, E.; Chiaramondia, M.; Milanese, M.; Rossi, G.A.; Brusasco, V. Increased numbers of mast cells in bronchial mucosa after the late-phase asthmatic response to allergen. Am. Rev. Respir. Dis. 1991, 144, 1282–1286. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabata, H.; Moro, K.; Fukunaga, K.; Suzuki, Y.; Miyata, J.; Masaki, K.; Koyasu, S.; Betsuyaku, T.; Asano, K. Pimozide, A C33 CYTOKINES AND ASTHMA MEDIATORS: Pimozide, A Stat5 Inhibitor, Counteracts Corticosteroid-Resistant Airway Inflammation Induced By Il-33 And Tslp. Am. J. Respir. Crit. Care Med. 2014, 189, 1. [Google Scholar]
- Rondanin, R.; Simoni, D.; Maccesi, M.; Romagnoli, R.; Grimaudo, S.; Pipitone, R.M.; Meli, M.; Cascio, A.; Tolomeo, M. Effects of Pimozide Derivatives on pSTAT5 in K562 Cells. ChemMedChem 2017, 12, 1183–1190. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, T.; Yi, L.; Zhou, X.; Mi, M. Dihydromyricetin inhibits NLRP3 inflammasome-dependent pyroptosis by activating the Nrf2 signaling pathway in vascular endothelial cells. Biofactors 2018, 44, 123–136. [Google Scholar] [CrossRef]
- Zeng, Y.; Peng, Y.; Tang, K.; Wang, Y.Q.; Zhao, Z.Y.; Wei, X.Y.; Xu, X.L. Dihydromyricetin ameliorates foam cell formation via LXRα-ABCA1/ABCG1-dependent cholesterol efflux in macrophages. Biomed. Pharmacother. 2018, 101, 543–552. [Google Scholar] [CrossRef]
- Manikandan, J.; Kothandaraman, N.; Hande, M.P.; Pushparaj, P.N. Deciphering the structure and function of FcεRI/mast cell axis in the regulation of allergy and anaphylaxis: A functional genomics paradigm. Cell. Mol. Life Sci. Cmls 2012, 69, 1917–1929. [Google Scholar] [CrossRef] [PubMed]
- Siebenhaar, F.; Redegeld, F.A.; Bischoff, S.C.; Gibbs, B.F.; Maurer, M. Mast Cells as Drivers of Disease and Therapeutic Targets. Trends Immunol. 2018, 39, 151–162. [Google Scholar] [CrossRef]
- Sun, S.-C.; Maggirwar, S.B.; Harhaj, E.W.; Uhlik, M. Binding of c-Rel to STAT5 target sequences in HTLV-I-transformed T cells. Oncogene 1999, 18, 1401–1409. [Google Scholar] [CrossRef] [Green Version]
- Morales, J.K.; Falanga, Y.T.; Depcrynski, A.; Fernando, J.; Ryan, J.J. Mast cell homeostasis and the JAK–STAT pathway. Genes Immun. 2010, 11, 599–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, T.; Xiao, W.; Gao, P.; Namiranian, S.; Matsumoto, K.; Tomimori, Y.; Hong, H.; Yamashita, H.; Kimura, M.; Kashiwakura, J.-I.; et al. Critical role for mast cell Stat5 activity in skin inflammation. Cell Rep. 2014, 6, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Bourgeais, J.; Gouilleux-Gruart, V.; Gouilleux, F. Oxidative metabolism in cancer: A STAT affair? Jak-Stat 2013, 2, e25764. [Google Scholar] [CrossRef] [Green Version]
- Mi, T.; Wang, Z.; Bunting, K.D. The Cooperative Relationship between STAT5 and Reactive Oxygen Species in Leukemia: Mechanism and Therapeutic Potential. Cancers 2018, 10, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Ferraresi, R.; Troiano, L.; Roat, E.; Gibellini, L.; Bertoncelli, L.; Nasi, M.; Pinti, M. Simultaneous analysis of reactive oxygen species and reduced glutathione content in living cells by polychromatic flow cytometry. Nat. Protoc. 2009, 4, 1790–1797. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.-M.; Hsiao, T.-C.; Yang, T.-Y.; Huang, H.-C. IgE-Induced Mast Cell Activation Is Suppressed by Dihydromyricetin through the Inhibition of NF-κB Signaling Pathway. Molecules 2021, 26, 3877. https://doi.org/10.3390/molecules26133877
Chang T-M, Hsiao T-C, Yang T-Y, Huang H-C. IgE-Induced Mast Cell Activation Is Suppressed by Dihydromyricetin through the Inhibition of NF-κB Signaling Pathway. Molecules. 2021; 26(13):3877. https://doi.org/10.3390/molecules26133877
Chicago/Turabian StyleChang, Tsong-Min, Tzu-Chih Hsiao, Ting-Ya Yang, and Huey-Chun Huang. 2021. "IgE-Induced Mast Cell Activation Is Suppressed by Dihydromyricetin through the Inhibition of NF-κB Signaling Pathway" Molecules 26, no. 13: 3877. https://doi.org/10.3390/molecules26133877