
Optimization of Tabersonine Methoxylation to Increase Vindoline Precursor Synthesis in Yeast Cell Factories

 , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
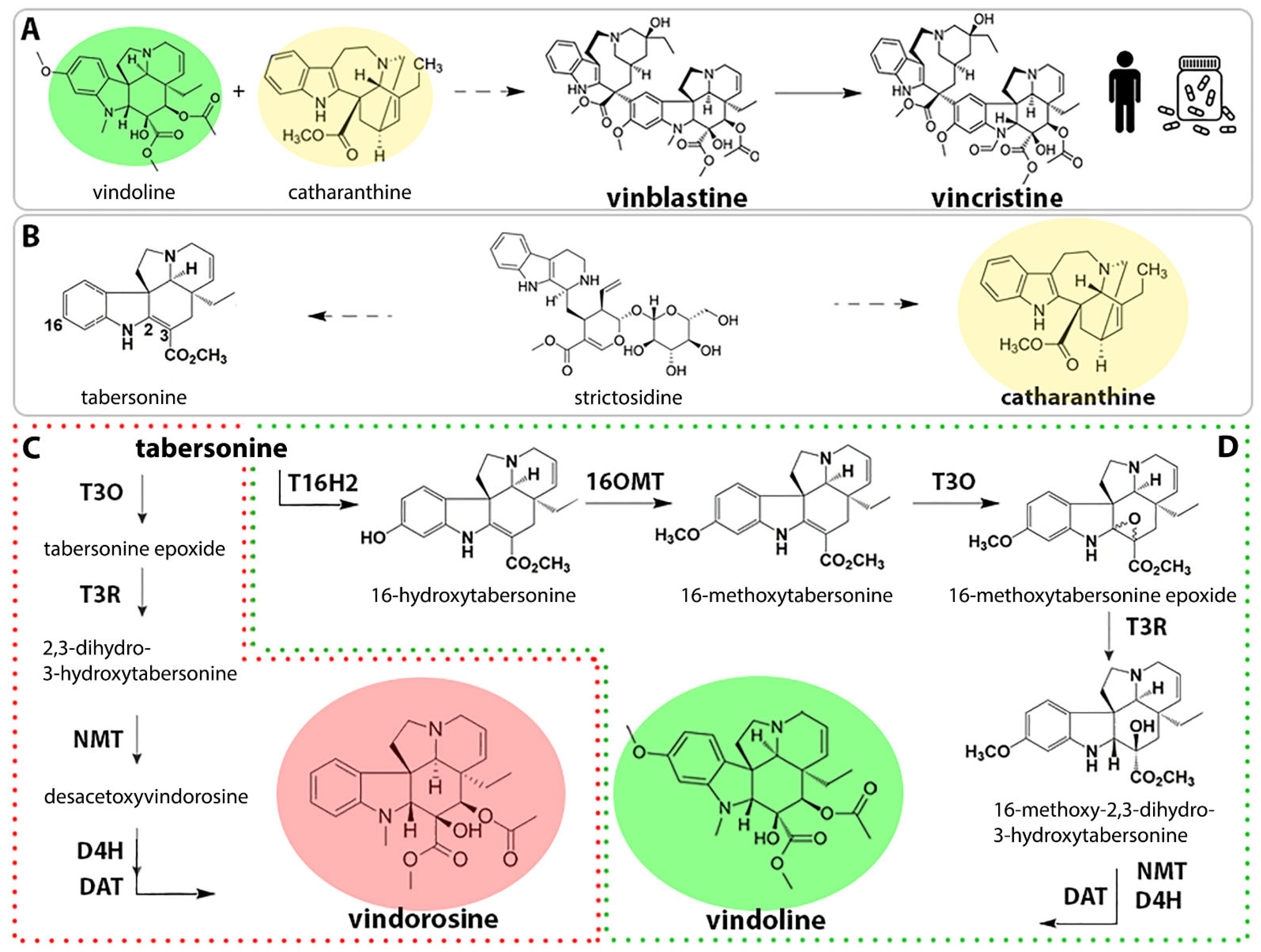
2. Results
2.1. Identification of Early Pathway Bottlenecks
2.2. Evaluation of S. cerevisiae Cell Permeability to 16-Hydroxytabersonine
2.3. Optimization of 16OMT Activity in Yeast
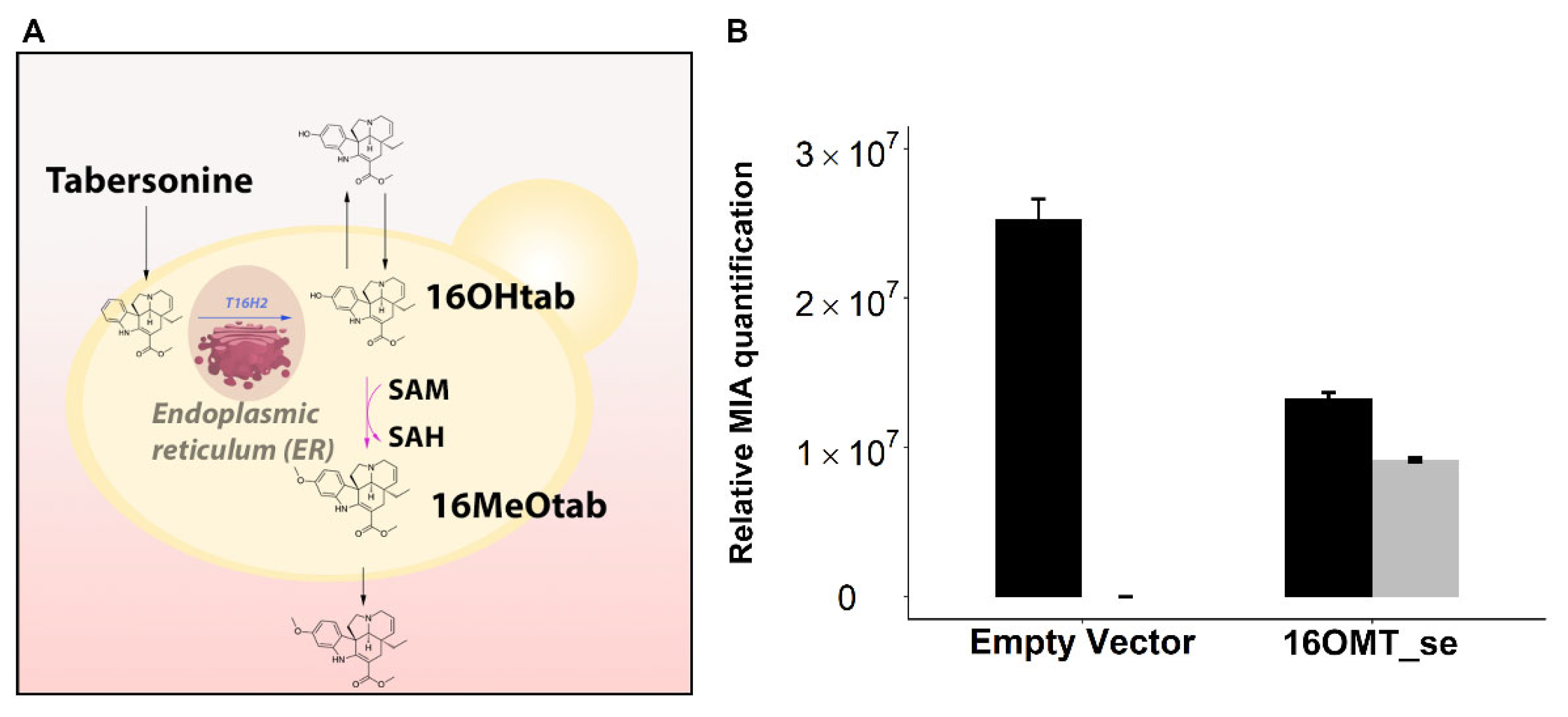
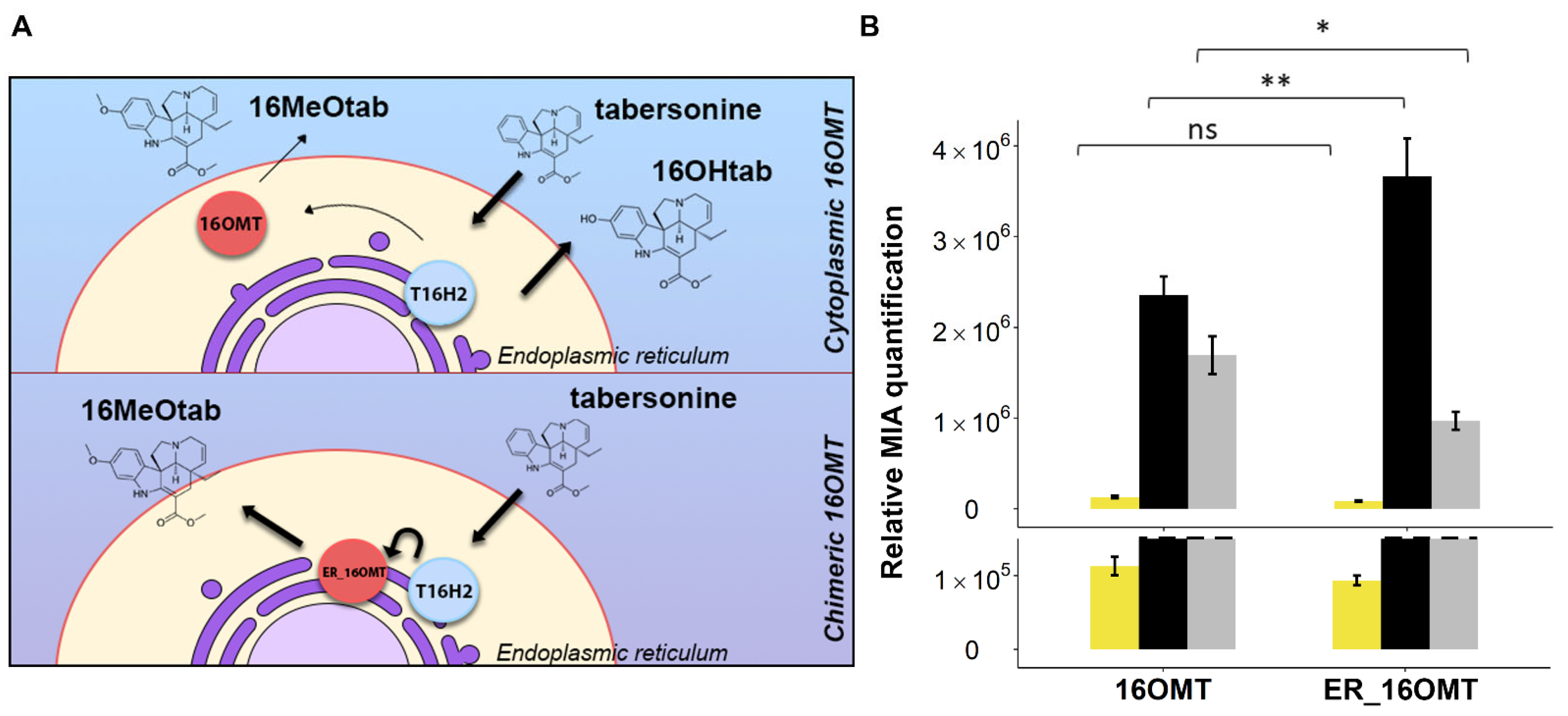
2.3.1. Targeting 16OMT to the Endoplasmic Reticulum to Favor Metabolic Channeling
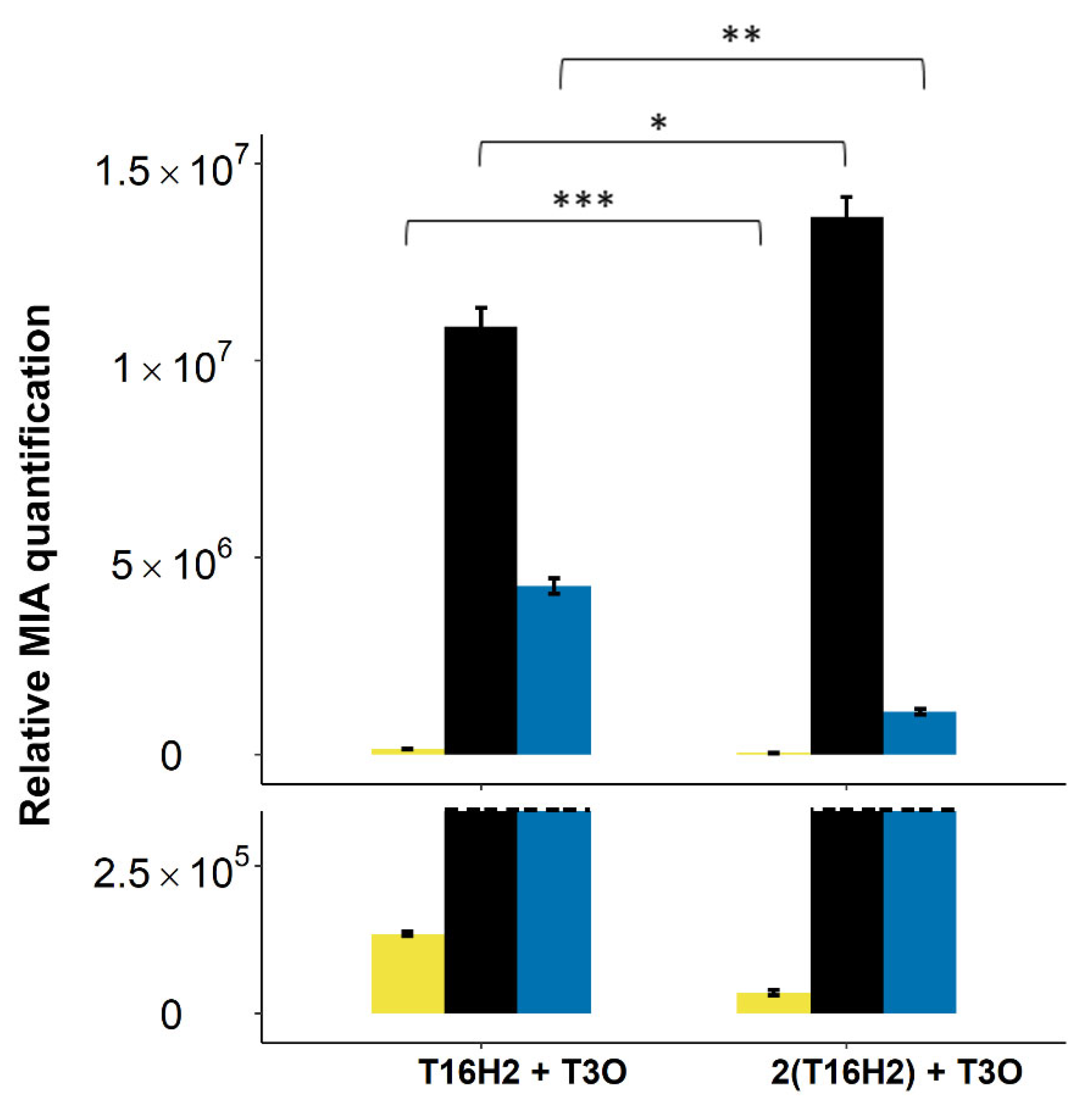
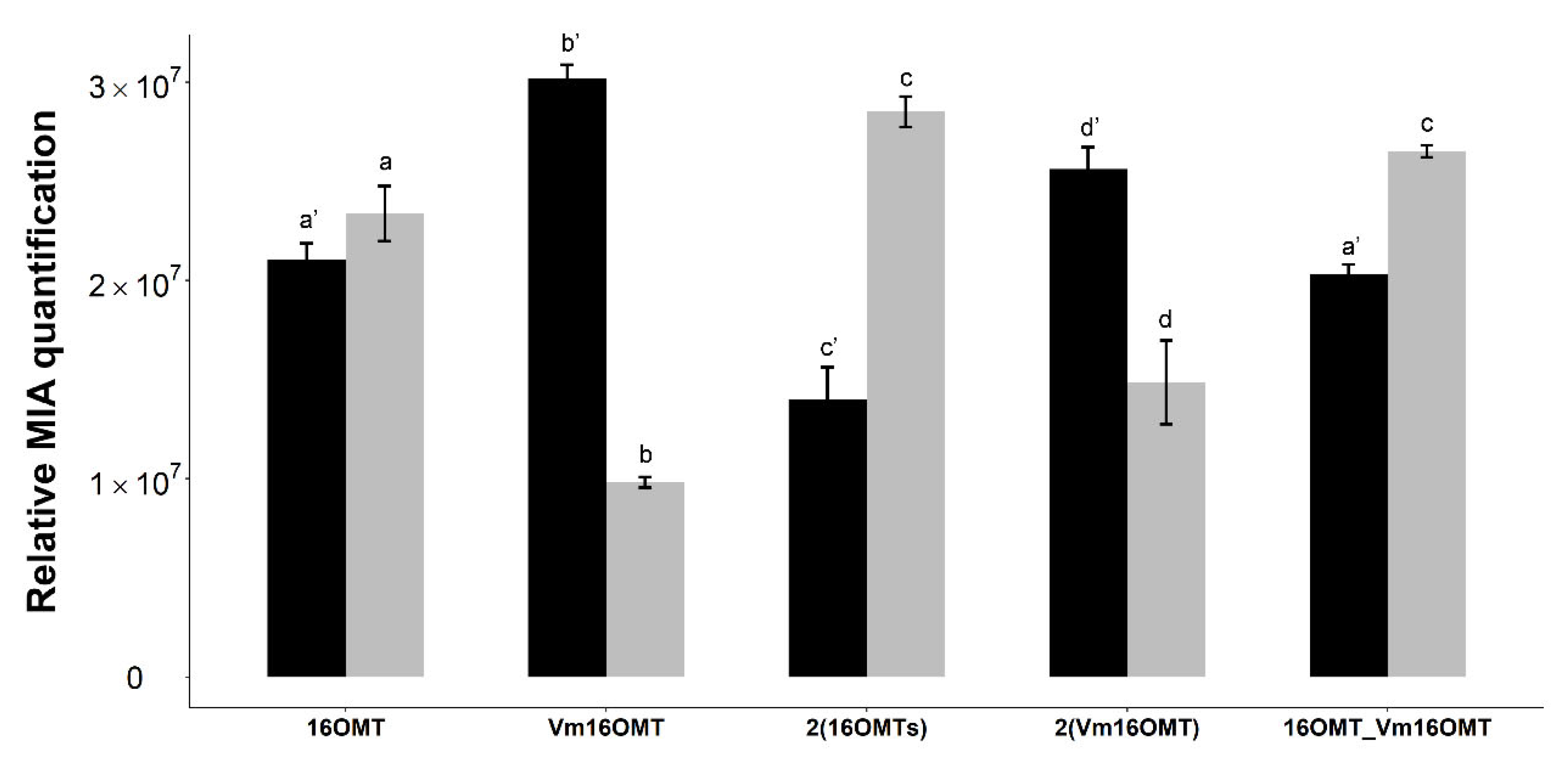
2.3.2. Testing a Distinct 16OMT Isoform and Increasing OMT Gene Copy Number to Limit 16-Hydroxytabersonine Accumulation
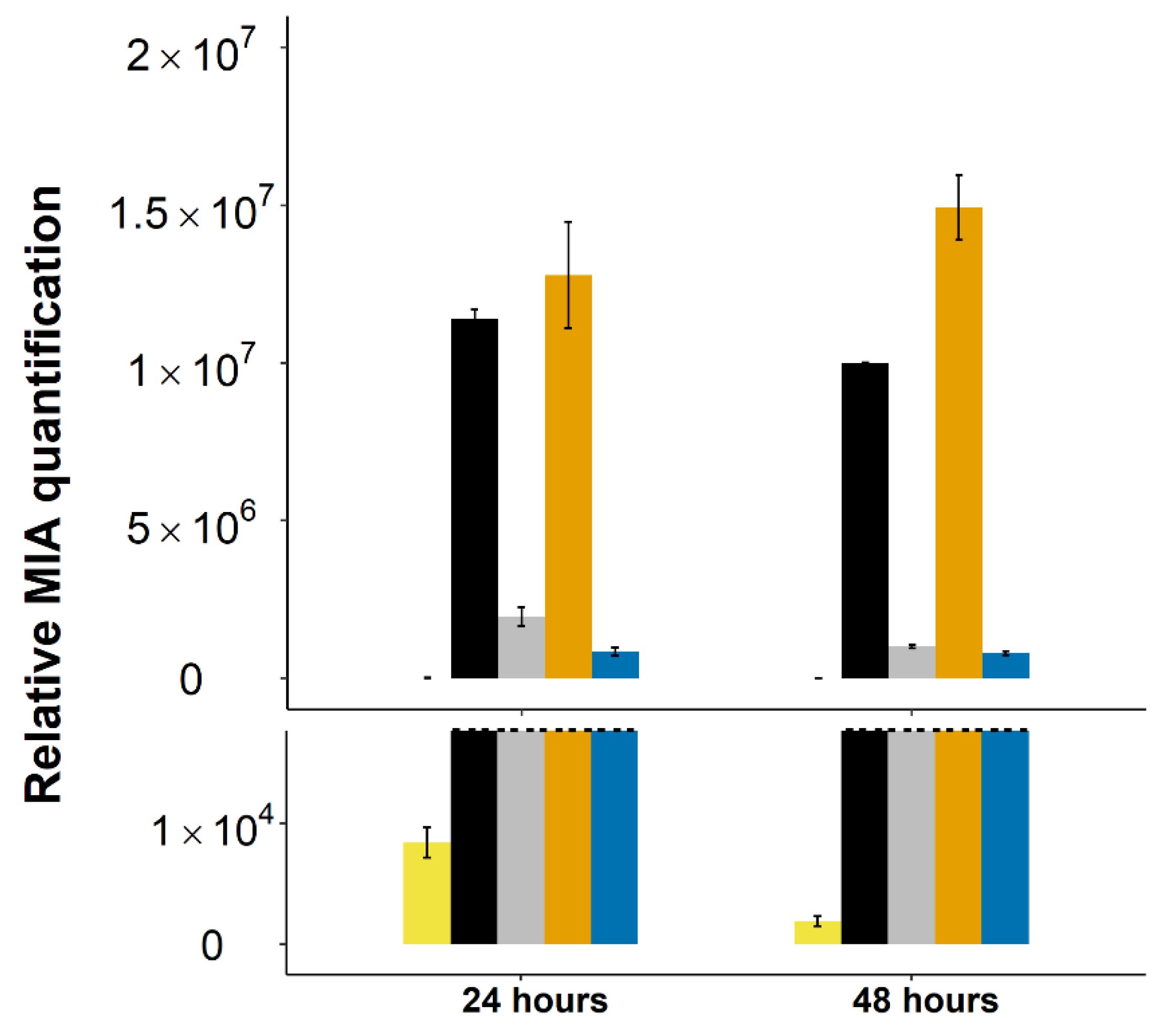
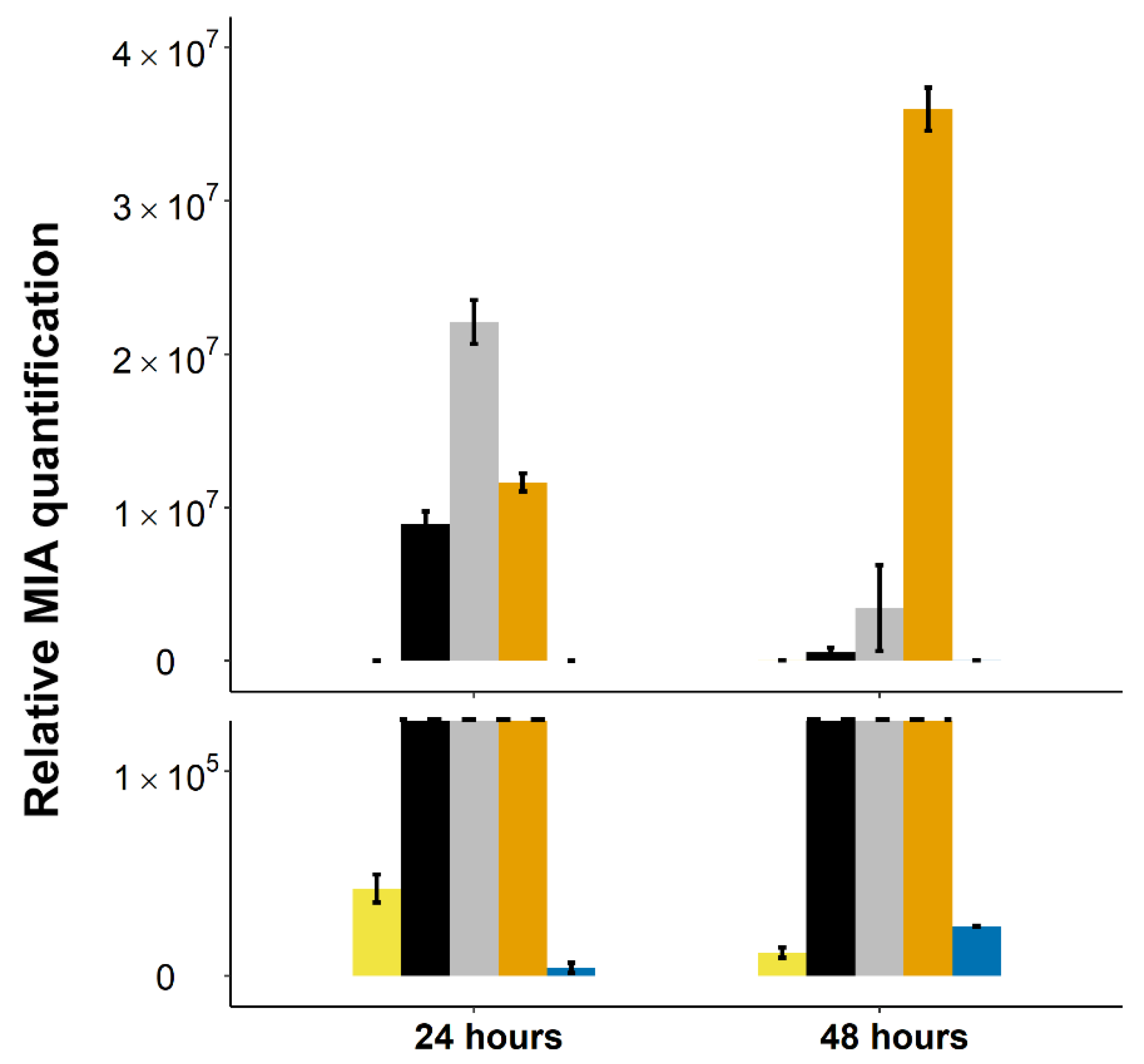
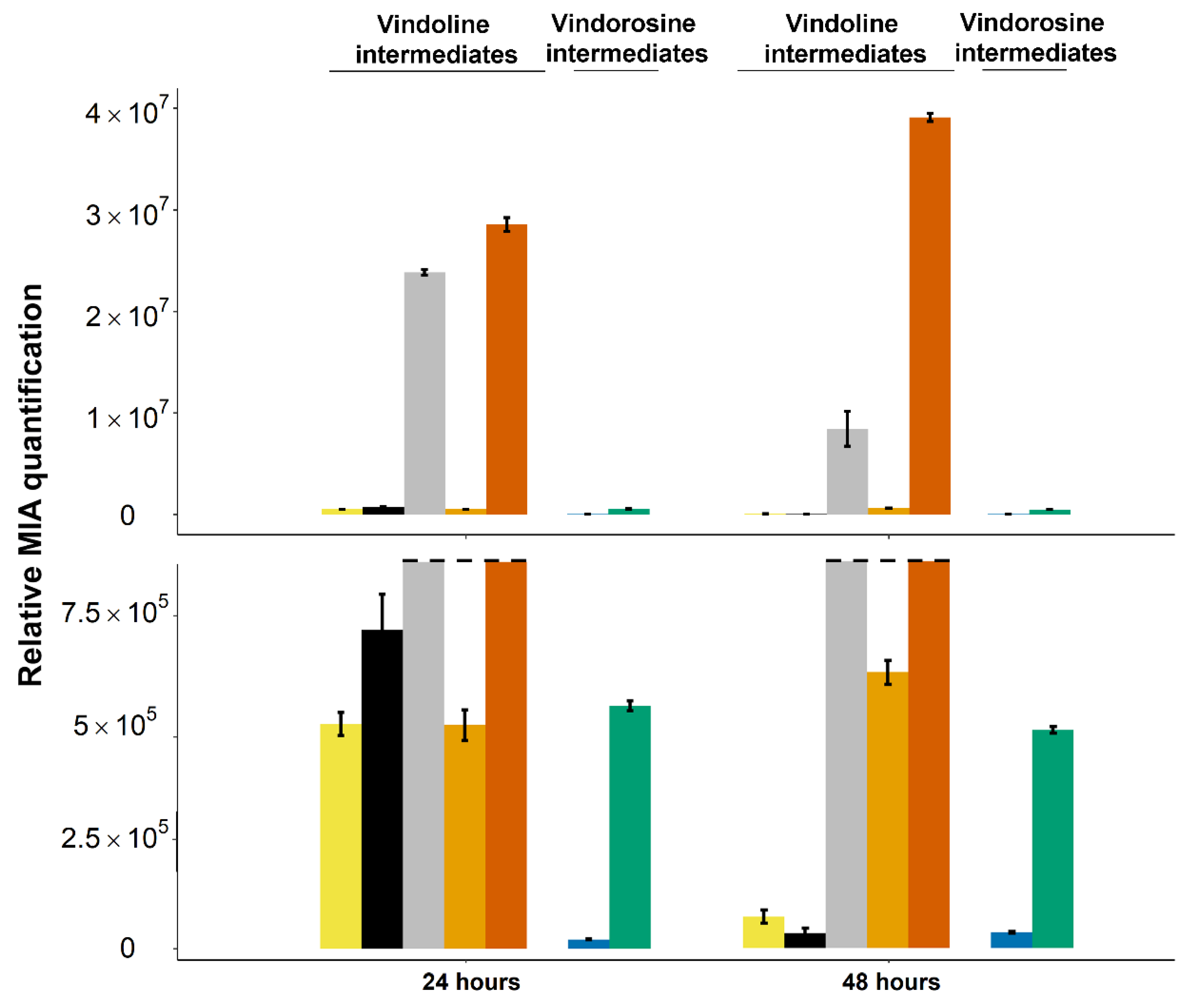
2.4. Stable Integration of the Four First Genes of the Vindoline Pathway Paves the Way for High Vindoline Production
3. Materials and Methods
3.1. Plasmid Construction
3.2. Yeast Strains
3.3. Yeast Transformations and Culture
3.4. Construction of the Chimeric 16OMT
3.5. Yeast Metabolite Analysis
3.6. Subcellular Localization in C. roseus Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Takshak, S.; Agrawal, S. Defense potential of secondary metabolites in medicinal plants under UV-B stress. J. Photochem. Photobiol. B Biol. 2019, 193, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Favarin, D.C.; De Oliveira, J.R.; De Oliveira, C.J.F.; Rogerio, A.D.P. Potential Effects of Medicinal Plants and Secondary Metabolites on Acute Lung Injury. BioMed Res. Int. 2013, 2013, 576479. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Modes of Action of Herbal Medicines and Plant Secondary Metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Bernath, J. The Genus Papaver; Harwood Academic Publishers: Amsterdam, The Netherlands; CRC Press: Boca Raton, FL, USA, 2000; pp. 169–170. ISBN 9789057022715. [Google Scholar]
- Paddon, C.J.; Keasling, J. Semi-synthetic artemisinin: A model for the use of synthetic biology in pharmaceutical development. Nat. Rev. Genet. 2014, 12, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.-W.; Yan, Z.; Amir, R.; Hong, S.; Jin, Y.-W.; Lee, E.-K.; Loake, G. Plant natural products: History, limitations and the potential of cambial meristematic cells. Biotechnol. Genet. Eng. Rev. 2012, 28, 47–60. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, B.; Besseau, S.; Clastre, M.; Courdavault, V.; Courtois, M.; Crèche, J.; Ducos, E.; De Bernonville, T.D.; Dutilleul, C.; Glevarec, G.; et al. Deciphering the Evolution, Cell Biology and Regulation of Monoterpene Indole Alkaloids. In Advances in Botanical Research; Elsevier BV: Amsterdam, The Netherlands, 2013; Volume 68, pp. 73–109. [Google Scholar]
- Srivastava, V.; Negi, A.S.; Kumar, J.; Gupta, M.; Khanuja, S.P. Plant-based anticancer molecules: A chemical and biological profile of some important leads. Bioorg. Med. Chem. 2005, 13, 5892–5908. [Google Scholar] [CrossRef]
- Alam, M.M.; Naeem, M.; Khan, M.M.A.; Uddin, M. Vincristine and vinblastine anticancer catharanthus alkaloids: Pharmacological applications and strategies for yield improvement. In Catharanthus Roseus; Springer: Berlin/Heidelberg, Germany, 2017; pp. 277–307. [Google Scholar]
- Noble, R.L. The discovery of the vinca alkaloids—chemotherapeutic agents against cancer. Biochem. Cell Biol. 1990, 68, 1344–1351. [Google Scholar] [CrossRef]
- Thomas, D.A.; Sarris, A.H.; Cortes, J.; Faderl, S.; O’Brien, S.; Giles, F.J.; Garcia-Manero, G.; Rodríguez, M.A.; Cabanillas, F.; Kantarjian, H. Phase II study of sphingosomal vincristine in patients with recurrent or refractory adult acute lymphocytic leukemia. Cancer 2005, 106, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; De Luca, V. ATP-binding cassette transporter controls leaf surface secretion of anticancer drug components in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2013, 110, 15830–15835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, V.; Laflamme, P. The expanding universe of alkaloid biosynthesis. Curr. Opin. Plant Biol. 2001, 4, 225–233. [Google Scholar] [CrossRef]
- Qu, Y.; Easson, M.; Froese, J.; Simionescu, R.; Hudlicky, T.; De Luca, V. Completion of the seven-step pathway from tabersonine to the anticancer drug precursor vindoline and its assembly in yeast. Proc. Natl. Acad. Sci. USA 2015, 112, 6224–6229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueritte, F.; Bac, N.V.; Langlois, Y.; Potier, P. Biosynthesis of antitumour alkaloids from Catharanthus roseus. Conversion of 20′-deoxyleurosidine into vinblastine. J. Chem. Soc. Chem. Commun. 1980, 10, 452–453. [Google Scholar] [CrossRef]
- O’Connor, S.E. Strategies for Engineering Plant Natural Products. Biomembr. Part K Membr. Biog. Assem. Target. (Prokaryotes Mitochondria Chloroplasts) 2012, 515, 189–206. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, W.; Rong, Q.; Jin, G.; Chu, H.; Liu, W.; Yang, W.; Zhu, Z.; Li, G.; Zhu, G.; et al. Modular Pathway Engineering of Diterpenoid Synthases and the Mevalonic Acid Pathway for Miltiradiene Production. J. Am. Chem. Soc. 2012, 134, 3234–3241. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Srivastava, P. Variation in vinblastine production by Catharanthus roseus, during in vivo and in vitro differentiation. Phytochemistry 1997, 46, 135–137. [Google Scholar] [CrossRef]
- Courdavault, V.; O’Connor, S.E.; Oudin, A.; Besseau, S.; Papon, N. Towards the Microbial Production of Plant-Derived Anticancer Drugs. Trends Cancer 2020, 6, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Kulagina, N.; Papon, N.; Hasunuma, T.; Courdavault, V. Innovative Tools and Strategies for Optimizing Yeast Cell Factories. Trends Biotechnol. 2021, 39, 488–504. [Google Scholar] [CrossRef]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nat. Cell Biol. 2013, 496, 528–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczebara, F.M.; Chandelier, C.; Villeret, C.; Masurel, A.; Bourot, S.; Duport, C.; Blanchard, S.; Groisillier, A.; Testet, E.; Costaglioli, P.; et al. Total biosynthesis of hydrocortisone from a simple carbon source in yeast. Nat. Biotechnol. 2003, 21, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Duport, C.; Spagnoli, R.; Degryse, E.; Pompon, D. Self-sufficient biosynthesis of pregnenolone and progesterone in engineered yeast. Nat. Biotechnol. 1998, 16, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Clastre, M.; Courdavault, V.; O’Connor, S.E. De novo production of the plant-derived alkaloid strictosidine in yeast. Proc. Natl. Acad. Sci. USA 2015, 112, 3205–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carqueijeiro, I.; De Bernonville, T.D.; LaNoue, A.; Dang, T.; Teijaro, C.; Paetz, C.; Billet, K.; Mosquera, A.; Oudin, A.; Besseau, S.; et al. A BAHD acyltransferase catalyzing 19- O -acetylation of tabersonine derivatives in roots of Catharanthus roseus enables combinatorial synthesis of monoterpene indole alkaloids. Plant J. 2018, 94, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carqueijeiro, I.T.; Brown, S.; Chung, K.; Dang, T.-T.T.; Walia, M.; Besseau, S.; De Bernonville, T.D.; Oudin, A.; LaNoue, A.; Billet, K.; et al. Two Tabersonine 6,7-Epoxidases Initiate Lochnericine-Derived Alkaloid Biosynthesis in Catharanthus roseus. Plant Physiol. 2018, 177, 1473–1486. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, K.M.; Smolke, C.D. Production of benzylisoquinoline alkaloids in Saccharomyces cerevisiae. Nat. Chem. Biol. 2008, 4, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Smolke, C.D. Engineering biosynthesis of the anticancer alkaloid noscapine in yeast. Nat. Commun. 2016, 7, 12137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, P.; Smolke, C.D. Biosynthesis of medicinal tropane alkaloids in yeast. Nat. Cell Biol. 2020, 585, 1–6. [Google Scholar] [CrossRef]
- Courdavault, V.; Cassereau, J.; Papon, N. Engineered Microbes for Producing Anticholinergics. ChemBioChem 2021, 22, 1368–1370. [Google Scholar] [CrossRef]
- Pyne, M.E.; Kevvai, K.; Grewal, P.S.; Narcross, L.; Choi, B.; Bourgeois, L.; Dueber, J.E.; Martin, V.J.J. A yeast platform for high-level synthesis of tetrahydroisoquinoline alkaloids. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kulagina, N.; Papon, N.; Courdavault, V. Microbial Cell Factories for Tetrahydroisoquinoline Alkaloid Production. ChemBioChem 2021, 22, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Courdavault, V.; Papon, N.; Clastre, M.; Giglioli-Guivarc’H, N.; St-Pierre, B.; Burlat, V. A look inside an alkaloid multisite plant: The Catharanthus logistics. Curr. Opin. Plant Biol. 2014, 19, 43–50. [Google Scholar] [CrossRef] [PubMed]
- De Bernonville, T.D.; Clastre, M.; Besseau, S.; Oudin, A.; Burlat, V.; Glévarec, G.; Lanoue, A.; Papon, N.; Giglioli-Guivarc’H, N.; St-Pierre, B.; et al. Phytochemical genomics of the Madagascar periwinkle: Unravelling the last twists of the alkaloid engine. Phytochemistry 2015, 113, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Guihur, A.; Poutrain, P.; Héricourt, F.; Mahroug, S.; St-Pierre, B.; Burlat, V.; Courdavault, V. Spatial organization of the vindoline biosynthetic pathway in Catharanthus roseus. J. Plant Physiol. 2011, 168, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Poisson, J.E. Process of Obtaining Tabersonine. U.S. Patent 3758478A, 16 June 1973. [Google Scholar]
- Walia, M.; Teijaro, C.N.; Gardner, A.; Tran, T.; Kang, J.; Zhao, S.; O’Connor, S.E.; Courdavault, V.; Andrade, R.B. Synthesis of (−)-Melodinine K: A Case Study of Efficiency in Natural Product Synthesis. J. Nat. Prod. 2020, 83, 2425–2433. [Google Scholar] [CrossRef]
- O’Connor, S.E.; Maresh, J.J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat. Prod. Rep. 2006, 23, 532–547. [Google Scholar] [CrossRef]
- St-Pierre, B.; De Luca, V. A Cytochrome P-450 Monooxygenase Catalyzes the First Step in the Conversion of Tabersonine to Vindoline in Catharanthus roseus. Plant Physiol. 1995, 109, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Schröder, G.; Unterbusch, E.; Kaltenbach, M.; Schmidt, J.; Strack, D.; De Luca, V.; Schröder, J. Light-induced cytochrome P450-dependent enzyme in indole alkaloid biosynthesis: Tabersonine 16-hydroxylase. FEBS Lett. 1999, 458, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Besseau, S.; Kellner, F.; Lanoue, A.; Thamm, A.M.; Salim, V.; Schneider, B.; Geu-Flores, F.; Höfer, R.; Guirimand, G.; Guihur, A.; et al. A Pair of Tabersonine 16-Hydroxylases Initiates the Synthesis of Vindoline in an Organ-Dependent Manner in Catharanthus roseus. Plant Physiol. 2013, 163, 1792–1803. [Google Scholar] [CrossRef] [Green Version]
- Levac, D.; Murata, J.; Kim, W.S.; De Luca, V. Application of carborundum abrasion for investigating the leaf epidermis: Molecular cloning of Catharanthus roseus 16-hydroxytabersonine-16-O-methyltransferase. Plant J. 2007, 53, 225–236. [Google Scholar] [CrossRef]
- De Luca, V.; Fernandez, J.A.; Campbell, D.; Kurz, W.G.W. Developmental Regulation of Enzymes of Indole Alkaloid Biosynthesis in Catharanthus roseus. Plant Physiol. 1988, 86, 447–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellner, F.; Geu-Flores, F.; Sherden, N.H.; Brown, S.; Foureau, E.; Courdavault, V.; O’Connor, S.E. Discovery of a P450-catalyzed step in vindoline biosynthesis: A link between the aspidosperma and eburnamine alkaloids. Chem. Commun. 2015, 51, 7626–7628. [Google Scholar] [CrossRef]
- Liscombe, D.K.; Usera, A.R.; O’Connor, S.E. Homolog of tocopherol C methyltransferases catalyzes N methylation in anticancer alkaloid biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 18793–18798. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Flota, F.; De Carolis, E.; Alarco, A.-M.; De Luca, V. Molecular cloning and characterization of desacetoxyvindoline-4-hydroxylase, a 2-oxoglutarate dependent-dioxygenase involved in the biosynthesis of vindoline in Catharanthus roseus (L.) G. Don. Plant Mol. Biol. 1997, 34, 935–948. [Google Scholar] [CrossRef] [PubMed]
- De Carolis, E.; De Luca, V. A novel 2-oxoglutarate-dependent dioxygenase involved in vindoline biosynthesis: Characterization, purification and kinetic properties. Plant Cell Tissue Organ Cult. (PCTOC) 1994, 38, 281–287. [Google Scholar] [CrossRef]
- Rabeharindranto, H.; Cerezo, S.C.; Lautier, T.; Garcia-Alles, L.; Treitz, C.; Tholey, A.; Truan, G. Enzyme-fusion strategies for redirecting and improving carotenoid synthesis in S. cerevisiae. Metab. Eng. Commun. 2019, 8, e00086. [Google Scholar] [CrossRef] [PubMed]
- Kulagina, N.; Besseau, S.; Papon, N.; Courdavault, V. Peroxisomes: A New Hub for Metabolic Engineering in Yeast. Front. Bioeng. Biotechnol. 2021, 9, 659431. [Google Scholar] [CrossRef]
- Zhu, J.; Schwartz, C.; Wheeldon, I. Controlled intracellular trafficking alleviates an expression bottleneck in S. cerevisiae ester biosynthesis. Metab. Eng. Commun. 2019, 8, e00085. [Google Scholar] [CrossRef] [PubMed]
- Thodey, K.; Galanie, S.; Smolke, C.D. A microbial biomanufacturing platform for natural and semisynthetic opioids. Nat. Chem. Biol. 2014, 10, 837–844. [Google Scholar] [CrossRef]
- Stander, E.A.; Sepúlveda, L.J.; De Bernonville, T.D.; Carqueijeiro, I.; Koudounas, K.; Cruz, P.L.; Besseau, S.; LaNoue, A.; Papon, N.; Giglioli-Guivarc’H, N.; et al. Identifying Genes Involved in alkaloid Biosynthesis in Vinca minor Through Transcriptomics and Gene Co-Expression Analysis. Biomolecules 2020, 10, 1595. [Google Scholar] [CrossRef]
- van Dijken, J.; Bauer, J.; Brambilla, L.; Duboc, P.; Francois, J.; Gancedo, C.; Giuseppin, M.; Heijnen, J.; Hoare, M.; Lange, H.; et al. An interlaboratory comparison of physiological and genetic properties of four Saccharomyces cerevisiae strains. Enzym. Microb. Technol. 2000, 26, 706–714. [Google Scholar] [CrossRef]
- Urban, P.; Mignotte, C.; Kazmaier, M.; Delorme, F.; Pompon, D. Cloning, Yeast Expression, and Characterization of the Coupling of Two Distantly Related Arabidopsis thalianaNADPH-Cytochrome P450 Reductases with P450 CYP73A5. J. Biol. Chem. 1997, 272, 19176–19186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, M.; Isobe, M.; Hatakeyama, T. Cryopreservation of competent intact yeast cells for efficient electroporation. Yeast 2000, 16, 889–896. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Parage, C.; Foureau, E.; Kellner, F.; Burlat, V.; Mahroug, S.; Lanoue, A.; de Bernonville, T.D.; Londono, M.A.; Carqueijeiro, I.; Oudin, A.; et al. Class II Cytochrome P450 Reductase Governs the Biosynthesis of Alkaloids. Plant Physiol. 2016, 172, 1563–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabin, R.C. Faced with a Drug Shortfall, Doctors Scramble to Treat. Children with Cancer. New York Times, 14 October 2019; Section A. 21. [Google Scholar]
- Courdavault, V.; O’Connor, S.E.; Jensen, M.K.; Papon, N. Metabolic engineering for plant natural products biosynthesis: New procedures, concrete achievements and remaining limits. Nat. Prod. Rep. 2021. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Plasmids |
|---|---|
| Inducible WAT11 strains | |
| T16H2+T3O | pYEDP60_T16H2, pESC-TRP_empty, pESC-HIS_empty, pESC-LEU_T3O |
| 2(T16H2)+T3O | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_empty, pESC-LEU_T3O |
| 2(T16H2)_T3O | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_empty, pESC-LEU_16OMT_T3O |
| 16OMT_se * | pESC_LEU_16OMT |
| ER_16OMT | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_empty, pESC-LEU_EROMT |
| 16OMT | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_16OMT, pESC-LEU_empty |
| 2(16OMTs) | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_16OMT, pESC-LEU_16OMT |
| Vm16OMT | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_Vm16OMT, pESC-LEU_empty |
| 2(Vm16OMT) | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_Vm16OMT, pESC-LEU_Vm16OMT |
| 16OMT_Vm16OMT | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_16OMT, pESC-LEU_Vm16OMT |
| T16H | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS empty, pESC-LEU_empty |
| 16OMT_T3O | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS empty, pESC-LEU_16OMT_T3O |
| 2(16OMTs)_T3O | pYEDP60_T16H2, pESC-TRP_T16H2, pESC-HIS_16OMT, pESC-LEU_16OMT_T3O |
| Empty_vectors | pYEDP60_empty, pESC-TRP_empty, pESC-HIS_empty, pESC-LEU_empty |
| Stable JDC058_CPR strains | |
| stable_2(16OMT)s | pHISA_pGK1-T16H2-tPRM5_pTEF1-16OMT-tCPS1, pURAK_pTDH3-T16H2-tPRM5_pTEF1T3O-tIDP1,pLEUA_pTEF1-16OMT-tCPS1_pGK1-T3R-tHIS5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemos Cruz, P.; Kulagina, N.; Guirimand, G.; De Craene, J.-O.; Besseau, S.; Lanoue, A.; Oudin, A.; Giglioli-Guivarc’h, N.; Papon, N.; Clastre, M.; et al. Optimization of Tabersonine Methoxylation to Increase Vindoline Precursor Synthesis in Yeast Cell Factories. Molecules 2021, 26, 3596. https://doi.org/10.3390/molecules26123596
Lemos Cruz P, Kulagina N, Guirimand G, De Craene J-O, Besseau S, Lanoue A, Oudin A, Giglioli-Guivarc’h N, Papon N, Clastre M, et al. Optimization of Tabersonine Methoxylation to Increase Vindoline Precursor Synthesis in Yeast Cell Factories. Molecules. 2021; 26(12):3596. https://doi.org/10.3390/molecules26123596
Chicago/Turabian StyleLemos Cruz, Pamela, Natalja Kulagina, Grégory Guirimand, Johan-Owen De Craene, Sébastien Besseau, Arnaud Lanoue, Audrey Oudin, Nathalie Giglioli-Guivarc’h, Nicolas Papon, Marc Clastre, and et al. 2021. "Optimization of Tabersonine Methoxylation to Increase Vindoline Precursor Synthesis in Yeast Cell Factories" Molecules 26, no. 12: 3596. https://doi.org/10.3390/molecules26123596