
Intrinsic and Chemotherapeutic Stressors Modulate ABCC-Like Transport in Trypanosoma cruzi

 , , , , and
, , , , and 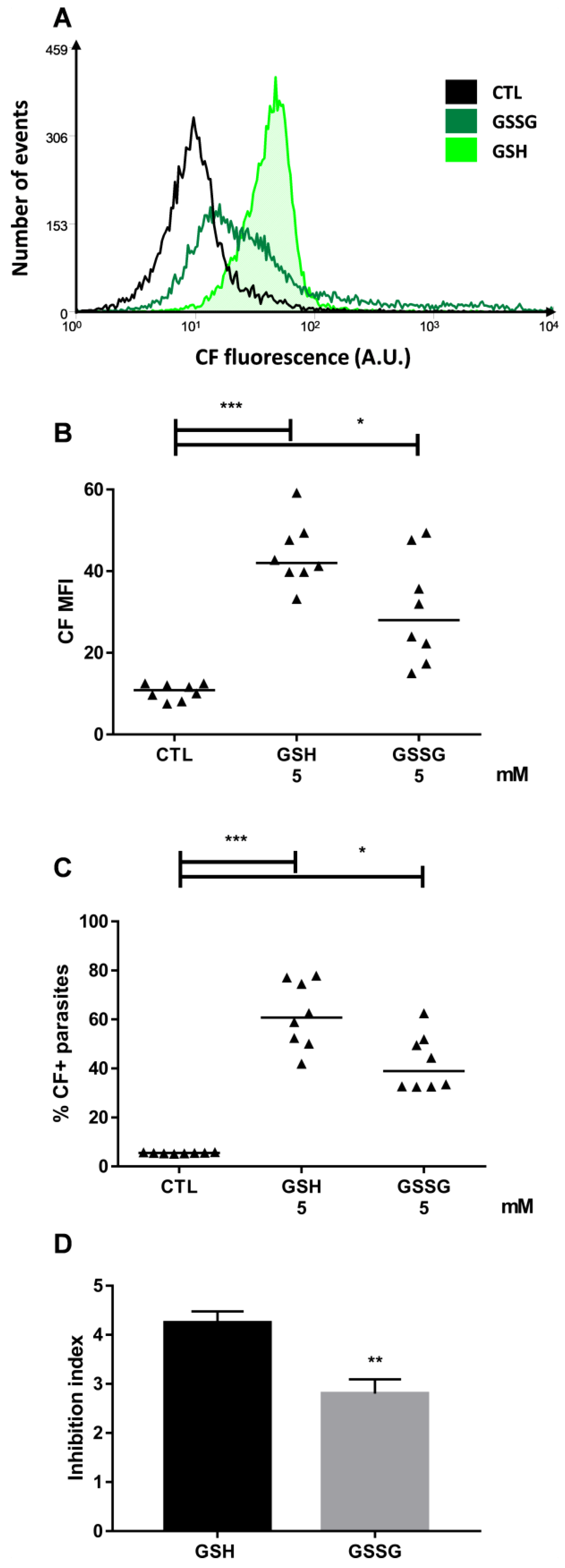{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Glutathione Promotes CF Accumulation
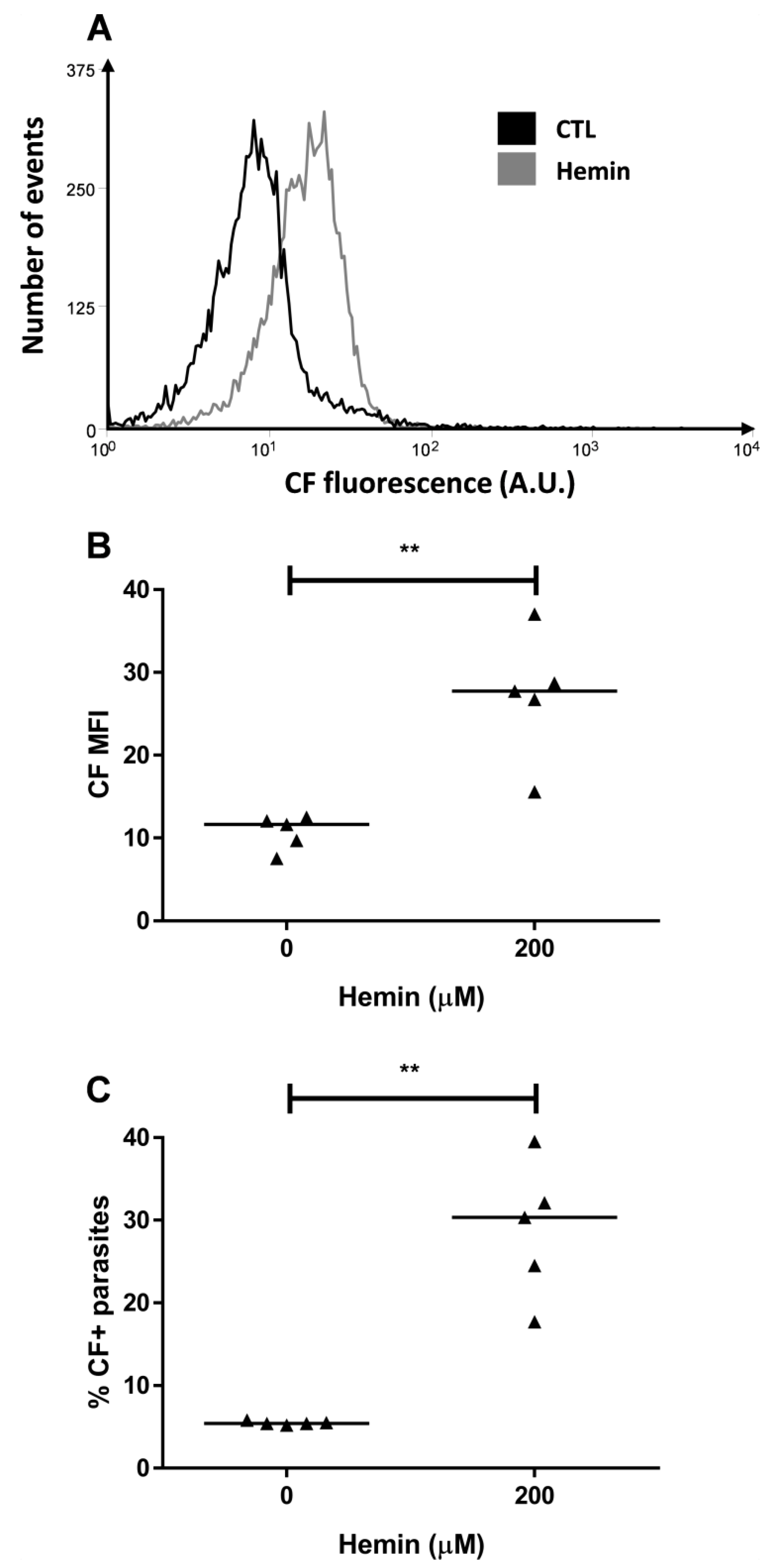
2.2. Hemin Preincubation Promotes Thiol Depletion and CF Accumulation
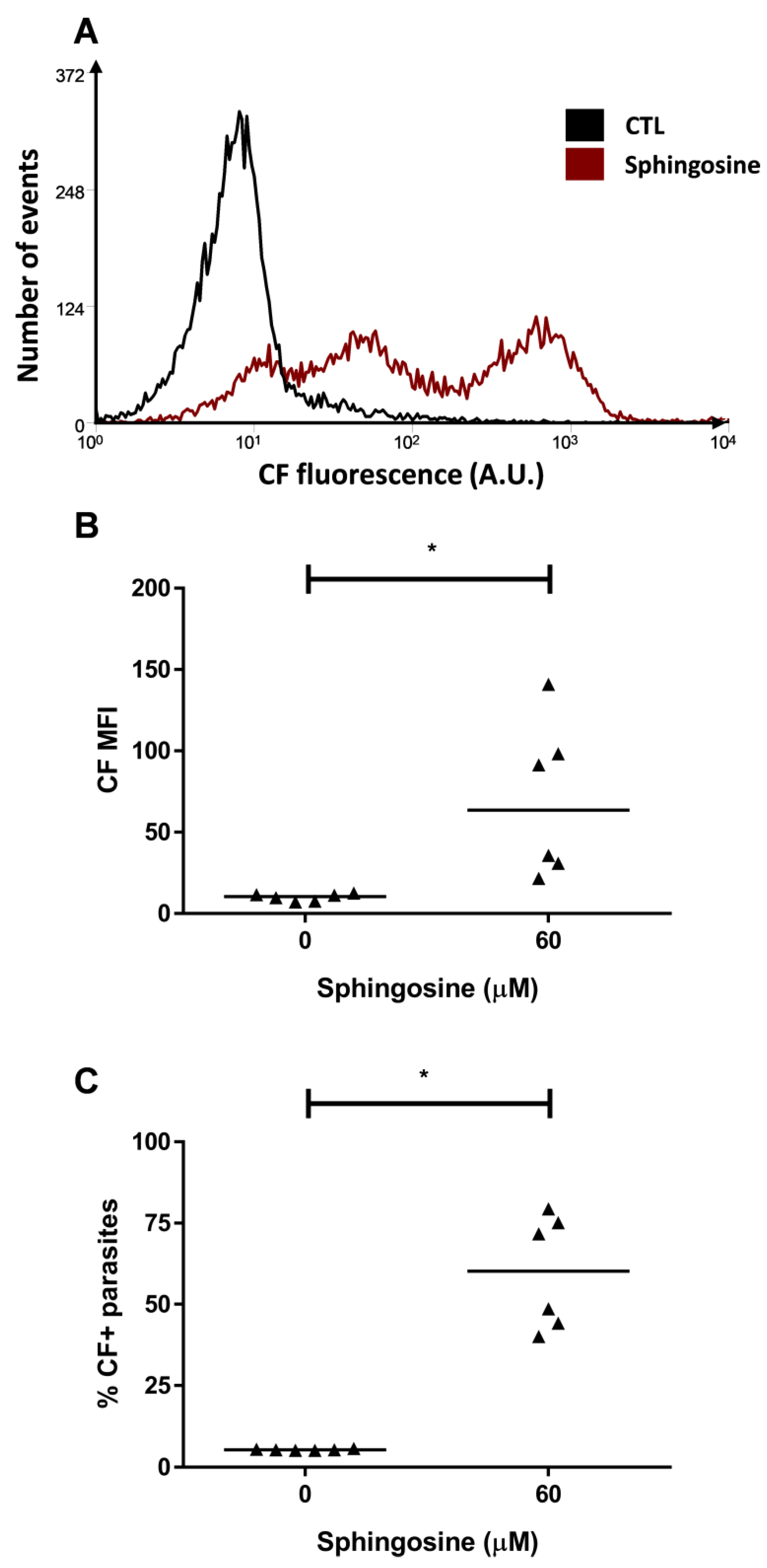
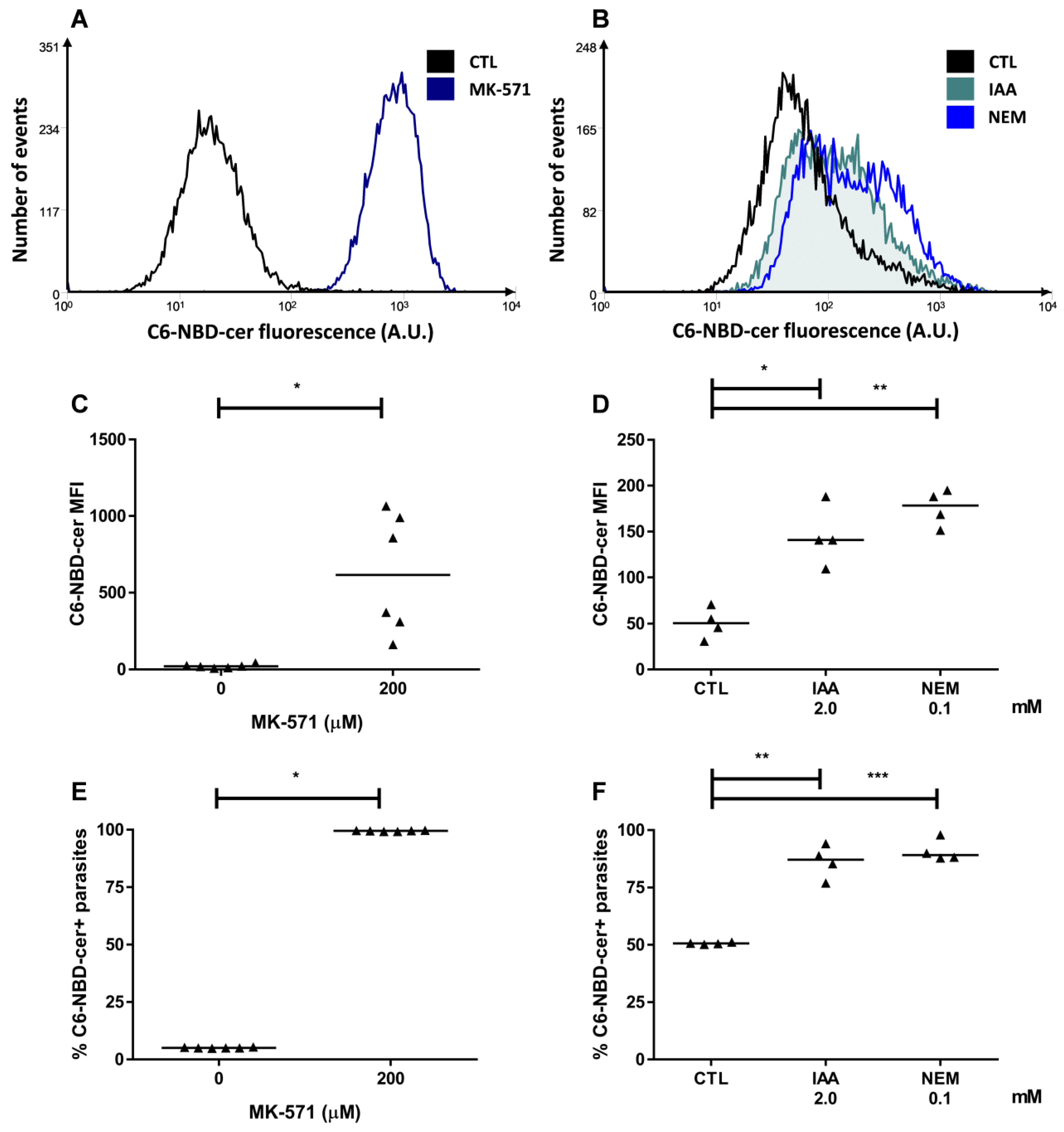
2.3. ABCC-Like Activity Mediates Ceramide Efflux
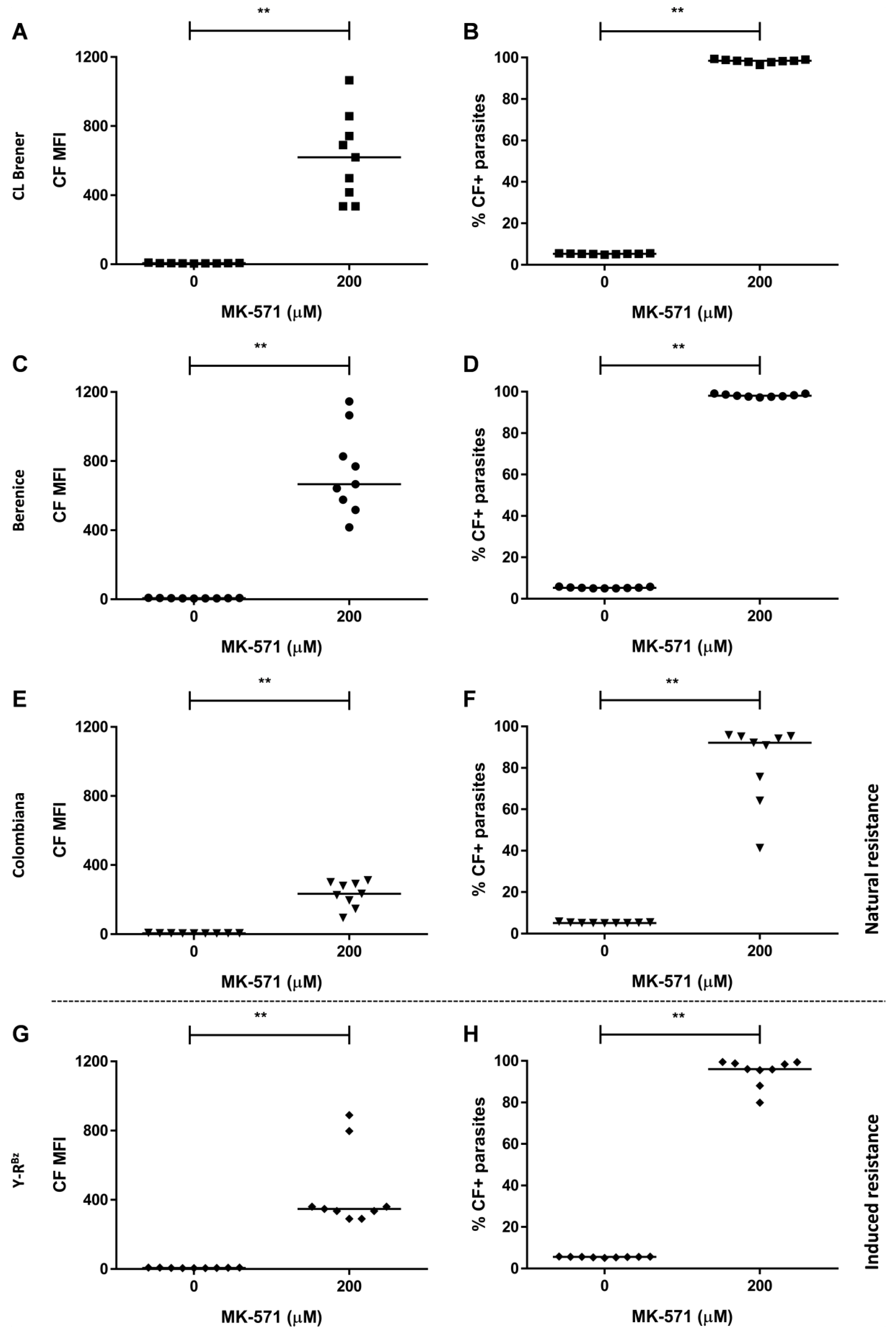
2.4. Resistance to Benznidazole
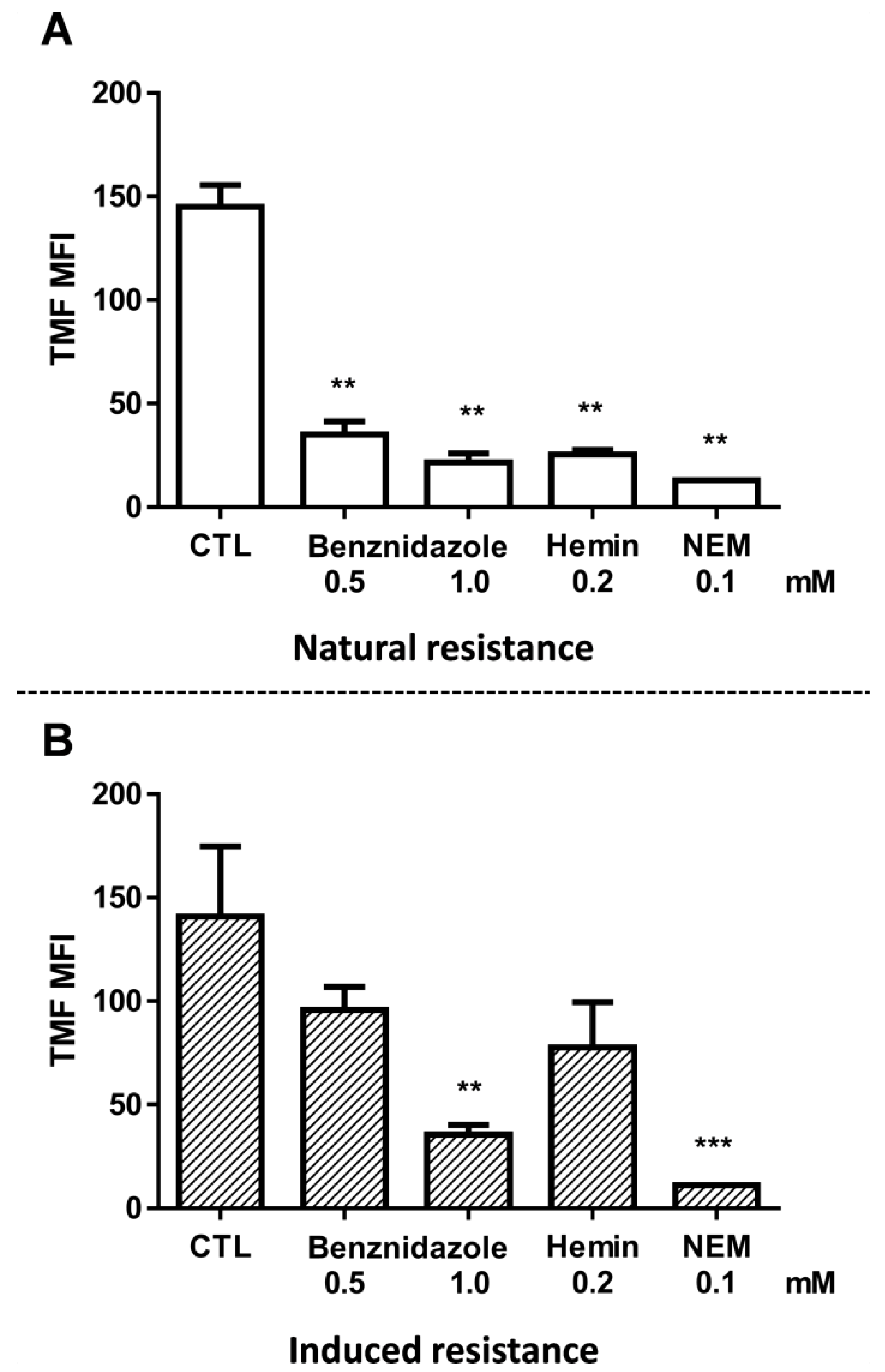
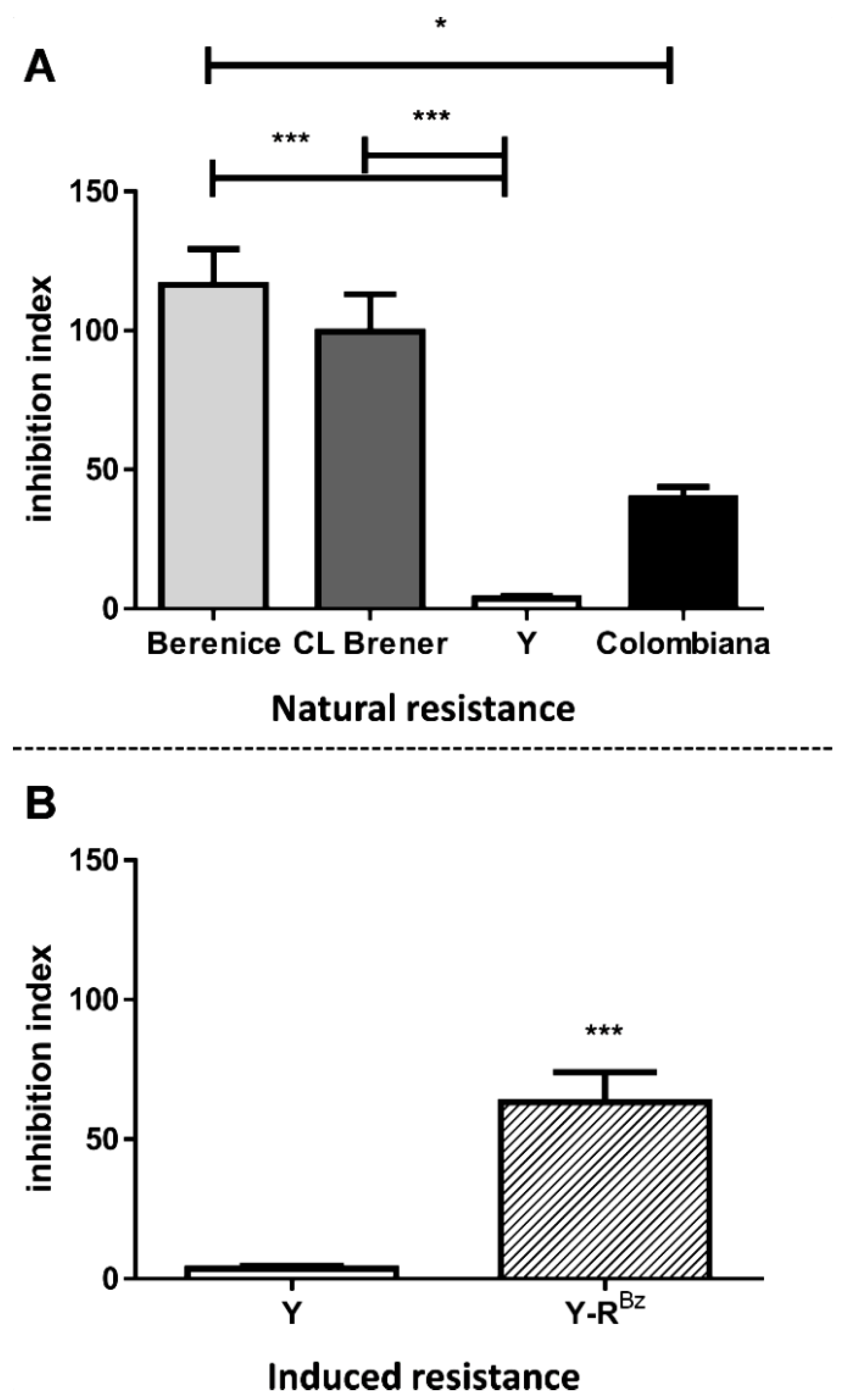
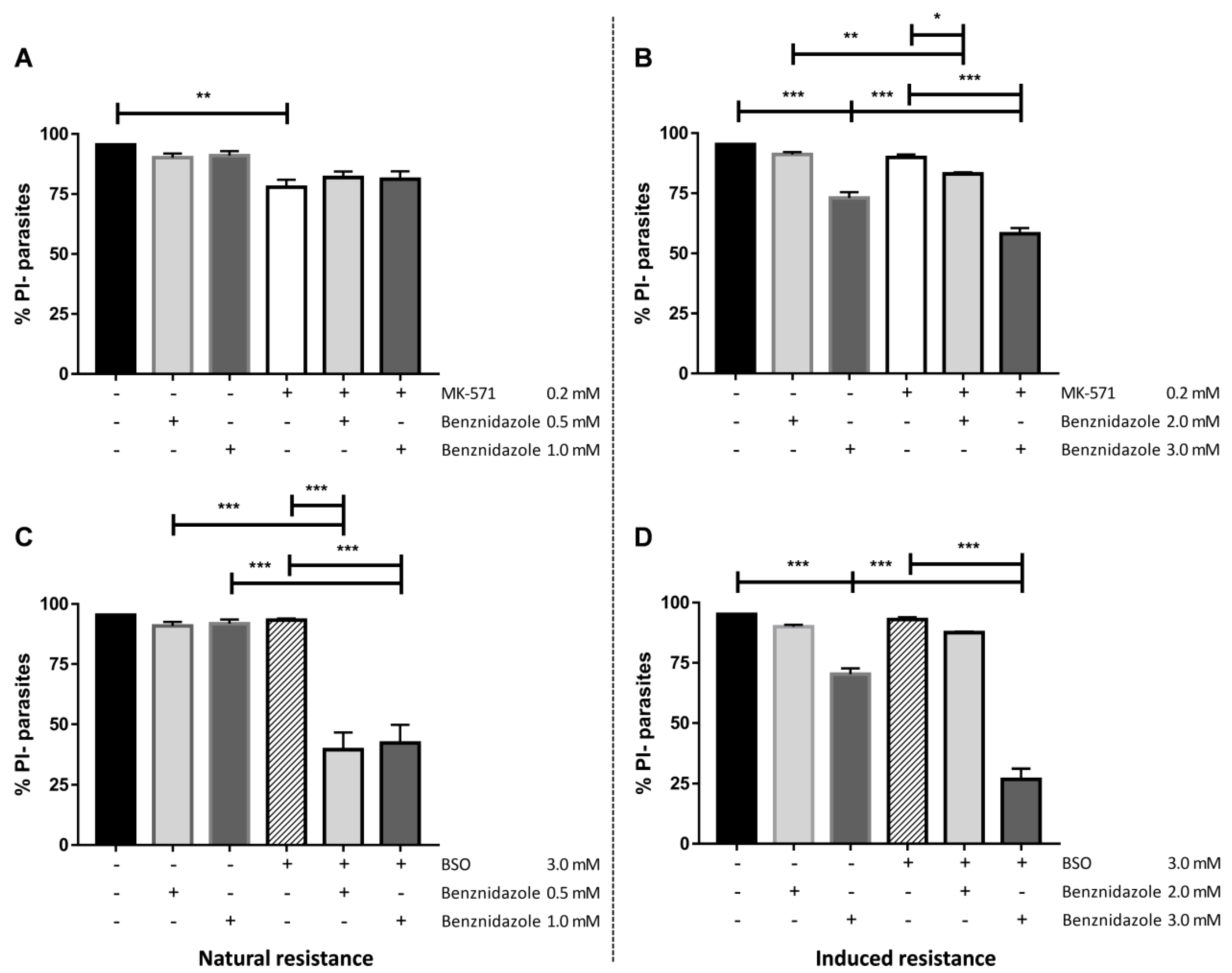
2.5. Natural Resistance to Benznidazole Does Not Relate to ABCC-Like Mediated Efflux
2.6. CF Efflux Increases after Acquired Resistance to Benznidazole
2.7. Resistance to Benznidazole Is Not Directly Dependent on ABCC-Like Efflux
3. Discussion
4. Materials and Methods
4.1. Cultures of Trypanosoma Cruzi Strains
4.2. MTT Reduction Assay
4.3. In Vitro Induction of Resistance to Benznidazole
4.4. ATP Depletion
4.5. ABC-Mediated Efflux Assay
4.6. Determination of Intracellular Thiols
4.7. Assessment of Cellular Viability
4.8. Flow Cytometry Analyses
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| Δ | inhibition index of transport |
| ABC | ATP-binding cassette |
| ABCB1 | subfamily B, member 1 (P-glycoprotein) |
| ABCC | subfamily C (MRP) |
| ABCG1 | subfamily G, member 1 |
| BSO | buthionine sulfoximine |
| Bz | benznidazole |
| C6-NBD-ceramide | N-[(E,2S,3R)-1,3-dihydroxyoctadec-4-en-2-yl]-6-[(4-nitro-2,1,3-benzoxadiazol-7-yl)amino]hexanamide |
| CF | carboxyfluorescein |
| CFDA | 5(6)-carboxyfluorescein diacetate |
| CMFDA | 5-chloromethylfluorescein diacetate |
| GSH | reduced glutathione |
| GSSG | glutathione disulfide form (oxidized glutathione) |
| IAA | Iodoacetic acid |
| MK-571 | (E)-3-[[[3-[2-(7-chloro-2-quinolinyl)ethenyl]phenyl][[3-(dimethylamino)-3-oxopropyl]thio]methyl]thio]-propanoic acid |
| MFI | median fluorescence intensity |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NEM | N-ethylmaleimide |
| PI | Propidium iodide |
| Rho 123 | rhodamine 123 |
| ROS | reactive oxygen species |
| TMF | thiol-conjugated methylfluorescein |
References
- Chagas, C. Nova tripanosomiase humana. Estudos sobre a morfologia e o ciclo evolutivo do Schizotrypanum cruzi n.g., n.sp., agente etiológico de nova entidade mórbida no homem. Mem. Inst. Oswaldo Cruz 1909, 1, 159–218. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Integrating Neglected Tropical Diseases into Global Health and Development: Fourth WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Perez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Pereira, K.S.; Schmidt, F.L.; Guaraldo, A.M.; Franco, R.M.; Dias, V.L.; Passos, L.A. Chagas’ disease as a foodborne illness. J. Food Prot. 2009, 72, 441–446. [Google Scholar] [CrossRef]
- Angheben, A.; Boix, L.; Buonfrate, D.; Gobbi, F.; Bisoffi, Z.; Pupella, S.; Gandini, G.; Aprili, G. Chagas disease and transfusion medicine: A perspective from non-endemic countries. Blood Transfus. Trasfus. Sangue 2015, 13, 540–550. [Google Scholar] [CrossRef]
- Carlier, Y.; Sosa-Estani, S.; Luquetti, A.O.; Buekens, P. Congenital Chagas disease: An update. Mem. Inst. Oswaldo Cruz 2015, 110, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.; Almeida, A.B.; Rosa, A.C.; Araujo, P.F.; Teixeira, A.R.L. American trypanosomiasis and Chagas disease: Sexual transmission. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2019, 81, 81–84. [Google Scholar] [CrossRef] [Green Version]
- Salazar, R.; Castillo-Neyra, R.; Tustin, A.W.; Borrini-Mayori, K.; Naquira, C.; Levy, M.Z. Bed bugs (Cimex lectularius) as vectors of Trypanosoma cruzi. Am. J. Trop. Med. Hyg. 2015, 92, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Scarim, C.B.; Jornada, D.H.; Chelucci, R.C.; de Almeida, L.; Dos Santos, J.L.; Chung, M.C. Current advances in drug discovery for Chagas disease. Eur. J. Med. Chem. 2018, 155, 824–838. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.S.; Wilkinson, S.R. Activation of benznidazole by trypanosomal type I nitroreductases results in glyoxal formation. Antimicrob. Agents Chemother. 2012, 56, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardi, L.S.; Brener, Z. Susceptibility and natural resistance of Trypanosoma cruzi strains to drugs used clinically in Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 755–759. [Google Scholar] [CrossRef]
- Murta, S.M.; Romanha, A.J. In vivo selection of a population of Trypanosoma cruzi and clones resistant to benznidazole. Parasitology 1998, 116, 165–171. [Google Scholar] [CrossRef]
- Campos, M.C.; Leon, L.L.; Taylor, M.C.; Kelly, J.M. Benznidazole-resistance in Trypanosoma cruzi: Evidence that distinct mechanisms can act in concert. Mol. Biochem. Parasitol. 2014, 193, 17–19. [Google Scholar] [CrossRef]
- Graca-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.; Paes, M.C.; Sorgine, M.H.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef]
- Nogueira, N.P.; Saraiva, F.M.S.; Oliveira, M.P.; Mendonca, A.P.M.; Inacio, J.D.F.; Almeida-Amaral, E.E.; Menna-Barreto, R.F.; Laranja, G.A.T.; Torres, E.J.L.; Oliveira, M.F.; et al. Heme modulates Trypanosoma cruzi bioenergetics inducing mitochondrial ROS production. Free Radic. Biol. Med. 2017, 108, 183–191. [Google Scholar] [CrossRef]
- Paes, M.C.; Cosentino-Gomes, D.; de Souza, C.F.; Nogueira, N.P.; Meyer-Fernandes, J.R. The Role of Heme and Reactive Oxygen Species in Proliferation and Survival of Trypanosoma cruzi. J. Parasitol. Res. 2011, 2011, 174614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, S.; Pena, L.A.; Kolesnick, R.N. Signal transduction of stress via ceramide. Biochem. J. 1998, 335, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Author Correction: Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannun, Y.A. Functions of ceramide in coordinating cellular responses to stress. Science 1996, 274, 1855–1859. [Google Scholar] [CrossRef]
- Won, J.S.; Singh, I. Sphingolipid signaling and redox regulation. Free Radic. Biol. Med. 2006, 40, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.P. Targeting multidrug resistance protein 1 (MRP1, ABCC1): Past, present, and future. Ann. Rev. Pharmacol. Toxicol. 2014, 54, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Mitra, P.; Oskeritzian, C.A.; Payne, S.G.; Beaven, M.A.; Milstien, S.; Spiegel, S. Role of ABCC1 in export of sphingosine-1-phosphate from mast cells. Proc. Natl. Acad. Sci. USA 2006, 103, 16394–16399. [Google Scholar] [CrossRef] [Green Version]
- Raggers, R.J.; van Helvoort, A.; Evers, R.; van Meer, G. The human multidrug resistance protein MRP1 translocates sphingolipid analogs across the plasma membrane. J. Cell Sci. 1999, 112, 415–422. [Google Scholar] [CrossRef]
- Salustiano, E.J.; da Costa, K.M.; Freire-de-Lima, L.; Mendonca-Previato, L.; Previato, J.O. Inhibition of glycosphingolipid biosynthesis reverts multidrug resistance by differentially modulating ABC transporters in chronic myeloid leukemias. J. Biol. Chem. 2020, 295, 6457–6471. [Google Scholar] [CrossRef]
- Sauvage, V.; Aubert, D.; Escotte-Binet, S.; Villena, I. The role of ATP-binding cassette (ABC) proteins in protozoan parasites. Mol. Biochem. Parasitol. 2009, 167, 81–94. [Google Scholar] [CrossRef]
- Dallagiovanna, B.; Castanys, S.; Gamarro, F. Trypanosoma cruzi: Sequence of the ATP-binding site of a P-glycoprotein gene. Exp. Parasitol. 1994, 79, 63–67. [Google Scholar] [CrossRef]
- Dallagiovanna, B.; Gamarro, F.; Castanys, S. Molecular characterization of a P-glycoprotein-related tcpgp2 gene in Trypanosoma cruzi. Mol. Biochem. Parasitol. 1996, 75, 145–157. [Google Scholar] [CrossRef]
- Leprohon, P.; Legare, D.; Girard, I.; Papadopoulou, B.; Ouellette, M. Modulation of Leishmania ABC protein gene expression through life stages and among drug-resistant parasites. Eukaryot. Cell 2006, 5, 1713–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, K.M.; Valente, R.C.; Salustiano, E.J.; Gentile, L.B.; Freire-de-Lima, L.; Mendonca-Previato, L.; Previato, J.O. Functional Characterization of ABCC Proteins from Trypanosoma cruzi and Their Involvement with Thiol Transport. Front. Microbiol. 2018, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slot, A.J.; Molinski, S.V.; Cole, S.P. Mammalian multidrug-resistance proteins (MRPs). Essays Biochem. 2011, 50, 179–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mina, J.G.M.; Denny, P.W. Everybody needs sphingolipids, right! Mining for new drug targets in protozoan sphingolipid biosynthesis. Parasitology 2018, 145, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Olin-Sandoval, V.; Moreno-Sanchez, R.; Saavedra, E. Targeting trypanothione metabolism in trypanosomatid human parasites. Curr. Drug Targets 2010, 11, 1614–1630. [Google Scholar] [CrossRef]
- Flohe, L. The trypanothione system and its implications in the therapy of trypanosomatid diseases. Int. J. Med. Microbiol. IJMM 2012, 302, 216–220. [Google Scholar] [CrossRef]
- Deeley, R.G.; Cole, S.P. Substrate recognition and transport by multidrug resistance protein 1 (ABCC1). FEBS Lett. 2006, 580, 1103–1111. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, N.M.; Myler, P.J.; Bartholomeu, D.C.; Nilsson, D.; Aggarwal, G.; Tran, A.N.; Ghedin, E.; Worthey, E.A.; Delcher, A.L.; Blandin, G.; et al. The genome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science 2005, 309, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, N.P.; de Souza, C.F.; Saraiva, F.M.; Sultano, P.E.; Dalmau, S.R.; Bruno, R.E.; Goncalves Rde, L.; Laranja, G.A.; Leal, L.H.; Coelho, M.G.; et al. Heme-induced ROS in Trypanosoma cruzi activates CaMKII-like that triggers epimastigote proliferation. One helpful effect of ROS. PLoS ONE 2011, 6, e25935. [Google Scholar] [CrossRef]
- Nogueira, N.P.; Saraiva, F.M.; Sultano, P.E.; Cunha, P.R.; Laranja, G.A.; Justo, G.A.; Sabino, K.C.; Coelho, M.G.; Rossini, A.; Atella, G.C.; et al. Proliferation and differentiation of Trypanosoma cruzi inside its vector have a new trigger: Redox status. PLoS ONE 2015, 10, e0116712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, T.H.; Frezzatti, W.A., Jr.; Schreier, S. Hemin-induced lipid membrane disorder and increased permeability: A molecular model for the mechanism of cell lysis. Arch. Biochem. Biophys. 1993, 307, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.R.; Dang, T.N.; Dringen, R.; Bishop, G.M. Hemin toxicity: A preventable source of brain damage following hemorrhagic stroke. Redox Rep. Commun. Free Radical Res. 2009, 14, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. The Ceramide-centric universe of lipid-mediated cell regulation: Stress encounters of the lipid kind. J. Biol. Chem. 2002, 277, 25847–25850. [Google Scholar] [CrossRef] [Green Version]
- Mendonca-Previato, L.; Todeschini, A.R.; Heise, N.; Previato, J.O. Protozoan parasite-specific carbohydrate structures. Curr. Opin. Struct. Biol. 2005, 15, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Andrieu-Abadie, N.; Levade, T.; Zhang, P.; Obeid, L.M.; Hannun, Y.A. Glutathione regulation of neutral sphingomyelinase in tumor necrosis factor-alpha-induced cell death. J. Biol. Chem. 1998, 273, 11313–11320. [Google Scholar] [CrossRef] [Green Version]
- Pankova-Kholmyansky, I.; Dagan, A.; Gold, D.; Zaslavsky, Z.; Skutelsky, E.; Gatt, S.; Flescher, E. Ceramide mediates growth inhibition of the Plasmodium falciparum parasite. Cell. Mol. Life Sci. CMLS 2003, 60, 577–587. [Google Scholar] [CrossRef]
- Morad, S.A.; Cabot, M.C. Tamoxifen regulation of sphingolipid metabolism–Therapeutic implications. Biochim. Biophys. Acta 2015, 1851, 1134–1145. [Google Scholar] [CrossRef] [Green Version]
- Trinconi, C.T.; Miguel, D.C.; Silber, A.M.; Brown, C.; Mina, J.G.M.; Denny, P.W.; Heise, N.; Uliana, S.R.B. Tamoxifen inhibits the biosynthesis of inositolphosphorylceramide in Leishmania. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 475–487. [Google Scholar] [CrossRef]
- Cabot, M.C.; Giuliano, A.E.; Volner, A.; Han, T.Y. Tamoxifen retards glycosphingolipid metabolism in human cancer cells. FEBS Lett. 1996, 394, 129–131. [Google Scholar] [CrossRef] [Green Version]
- Miguel, D.C.; Zauli-Nascimento, R.C.; Yokoyama-Yasunaka, J.K.; Katz, S.; Barbieri, C.L.; Uliana, S.R. Tamoxifen as a potential antileishmanial agent: Efficacy in the treatment of Leishmania braziliensis and Leishmania chagasi infections. J. Antimicrob. Chemother. 2009, 63, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Daleke, D.L. Phospholipid flippases. J. Biol. Chem. 2007, 282, 821–825. [Google Scholar] [CrossRef] [Green Version]
- Mejia-Jaramillo, A.M.; Fernandez, G.J.; Palacio, L.; Triana-Chavez, O. Gene expression study using real-time PCR identifies an NTR gene as a major marker of resistance to benzonidazole in Trypanosoma cruzi. Parasit Vectors 2011, 4, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piacenza, L.; Peluffo, G.; Alvarez, M.N.; Martinez, A.; Radi, R. Trypanosoma cruzi antioxidant enzymes as virulence factors in Chagas disease. Antioxid. Redox Signal. 2013, 19, 723–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faundez, M.; Pino, L.; Letelier, P.; Ortiz, C.; Lopez, R.; Seguel, C.; Ferreira, J.; Pavani, M.; Morello, A.; Maya, J.D. Buthionine sulfoximine increases the toxicity of nifurtimox and benznidazole to Trypanosoma cruzi. Antimicrob. Agents Chemother. 2005, 49, 126–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faundez, M.; Lopez-Munoz, R.; Torres, G.; Morello, A.; Ferreira, J.; Kemmerling, U.; Orellana, M.; Maya, J.D. Buthionine sulfoximine has anti-Trypanosoma cruzi activity in a murine model of acute Chagas’ disease and enhances the efficacy of nifurtimox. Antimicrob. Agents Chemother. 2008, 52, 1837–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, F.B.; Ruiz, J.C.; Robello, C.; Romanha, A.J.; Murta, S.M. Molecular characterization of cytosolic and mitochondrial tryparedoxin peroxidase in Trypanosoma cruzi populations susceptible and resistant to benznidazole. Parasitol. Res. 2009, 104, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, F.B.; Krieger, M.A.; Nirde, P.; Goldenberg, S.; Romanha, A.J.; Murta, S.M. Increased expression of iron-containing superoxide dismutase-A (TcFeSOD-A) enzyme in Trypanosoma cruzi population with in vitro-induced resistance to benznidazole. Acta Trop. 2006, 100, 119–132. [Google Scholar] [CrossRef]
- Zingales, B.; Araujo, R.G.; Moreno, M.; Franco, J.; Aguiar, P.H.; Nunes, S.L.; Silva, M.N.; Ienne, S.; Machado, C.R.; Brandao, A. A novel ABCG-like transporter of Trypanosoma cruzi is involved in natural resistance to benznidazole. Mem. Inst. Oswaldo Cruz 2015, 110, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Franco, J.; Ferreira, R.C.; Ienne, S.; Zingales, B. ABCG-like transporter of Trypanosoma cruzi involved in benznidazole resistance: Gene polymorphisms disclose inter-strain intragenic recombination in hybrid isolates. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 31, 198–208. [Google Scholar] [CrossRef]
- Angelani, C.R.; Carabias, P.; Cruz, K.M.; Delfino, J.M.; de Sautu, M.; Espelt, M.V.; Ferreira-Gomes, M.S.; Gomez, G.E.; Mangialavori, I.C.; Manzi, M.; et al. A metabolic control analysis approach to introduce the study of systems in biochemistry: The glycolytic pathway in the red blood cell. Biochem. Mol. Biol. Educ. Bimon. Publ. Int. Union Biochem. Mol. Biol. 2018, 46, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Mandal, G.; Singh, N.; Sundar, S.; Chatterjee, M. Flow cytometric determination of intracellular non-protein thiols in Leishmania promastigotes using 5-chloromethyl fluorescein diacetate. Exp. Parasitol. 2009, 122, 299–305. [Google Scholar] [CrossRef]
- Echevarria-Lima, J.; Kyle-Cezar, F.; DF, P.L.; Capella, L.; Capella, M.A.; Rumjanek, V.M. Expression and activity of multidrug resistance protein 1 in a murine thymoma cell line. Immunology 2005, 114, 468–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa, K.M.; Salustiano, E.J.; Valente, R.d.C.; Freire-de-Lima, L.; Mendonça-Previato, L.; Previato, J.O. Intrinsic and Chemotherapeutic Stressors Modulate ABCC-Like Transport in Trypanosoma cruzi. Molecules 2021, 26, 3510. https://doi.org/10.3390/molecules26123510
da Costa KM, Salustiano EJ, Valente RdC, Freire-de-Lima L, Mendonça-Previato L, Previato JO. Intrinsic and Chemotherapeutic Stressors Modulate ABCC-Like Transport in Trypanosoma cruzi. Molecules. 2021; 26(12):3510. https://doi.org/10.3390/molecules26123510
Chicago/Turabian Styleda Costa, Kelli Monteiro, Eduardo J. Salustiano, Raphael do Carmo Valente, Leonardo Freire-de-Lima, Lucia Mendonça-Previato, and José Osvaldo Previato. 2021. "Intrinsic and Chemotherapeutic Stressors Modulate ABCC-Like Transport in Trypanosoma cruzi" Molecules 26, no. 12: 3510. https://doi.org/10.3390/molecules26123510