
Species Diversity and Secondary Metabolites of Sarcophyton-Associated Marine Fungi


Abstract
:1. Soft Corals
2. Diversity of Sarcophyton
3. Sarcophyton-Fungal Associations
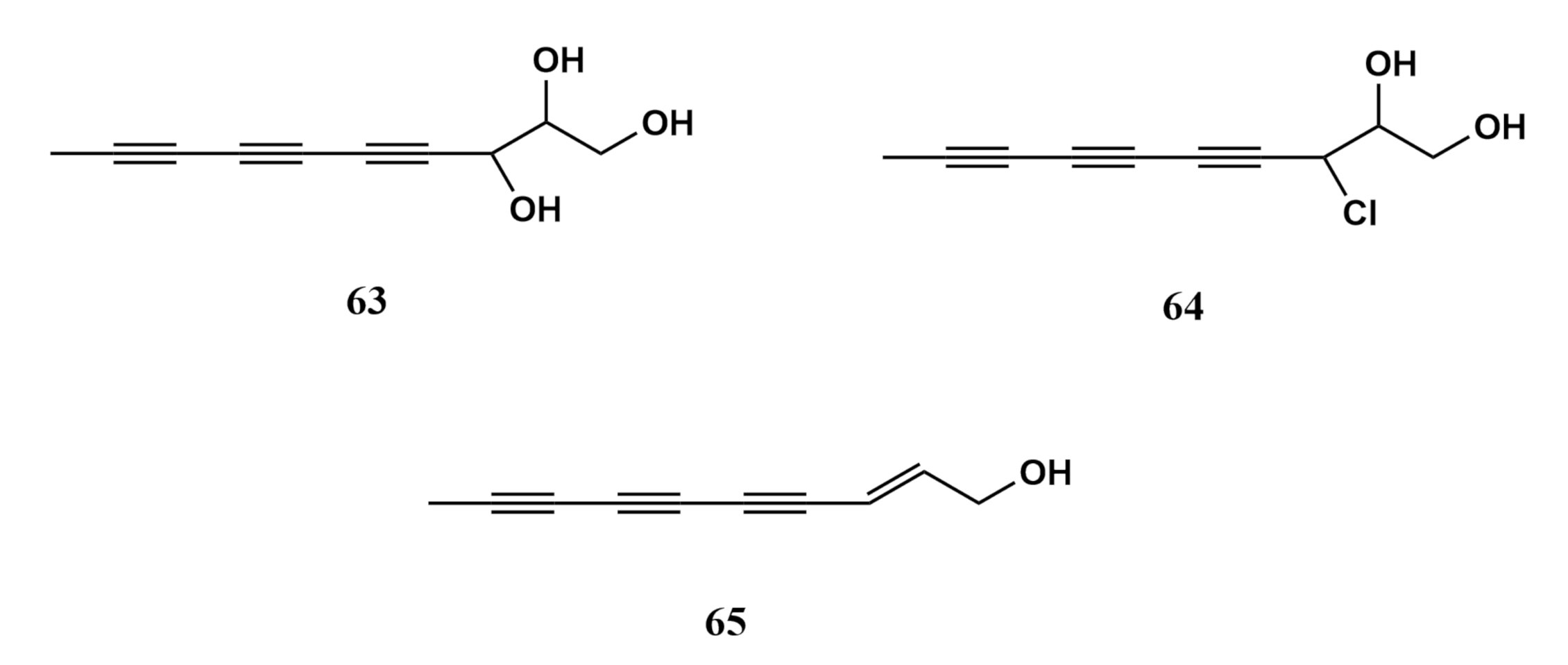
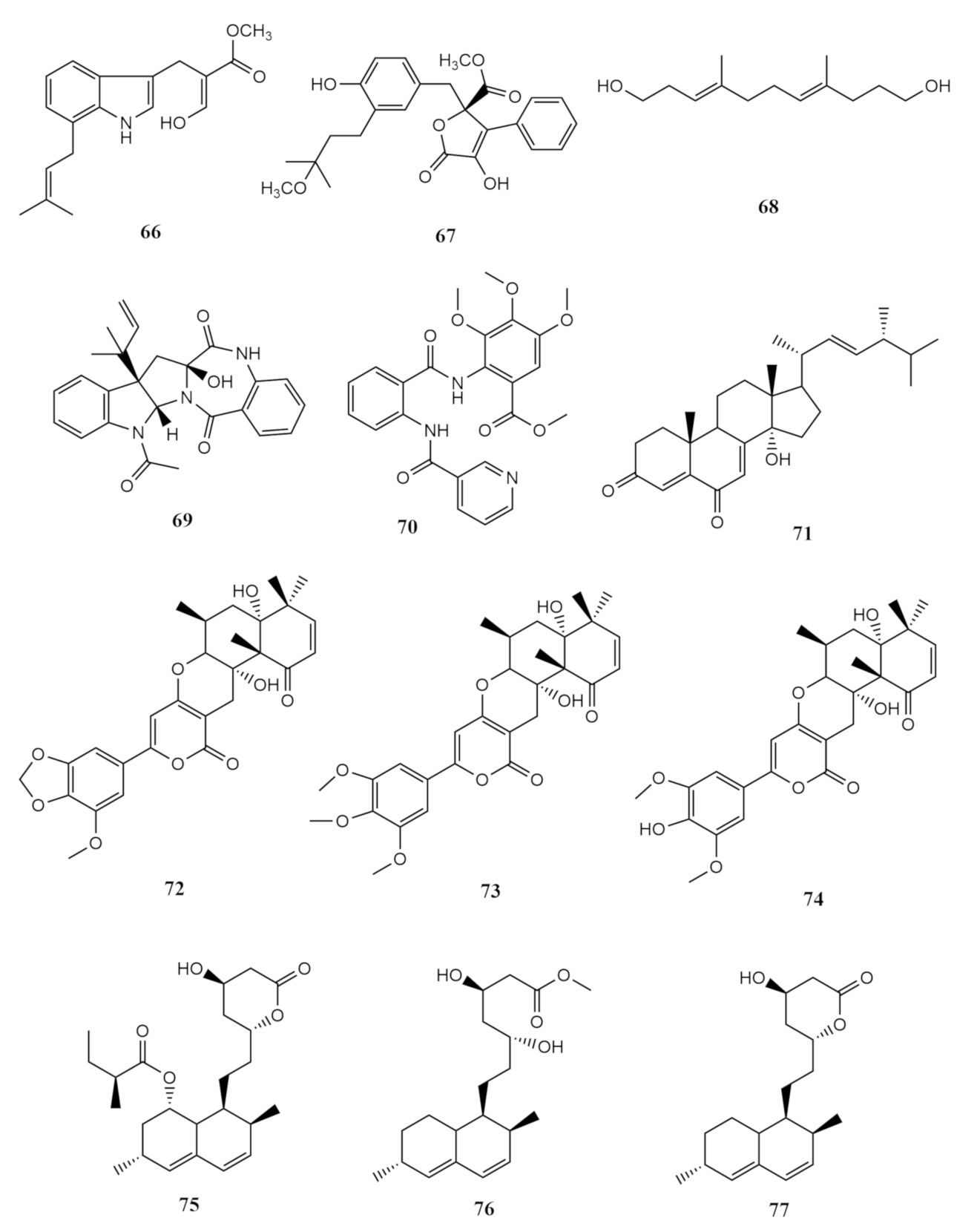
4. Metabolites of Marine Fungi Derived from Sarcophytons
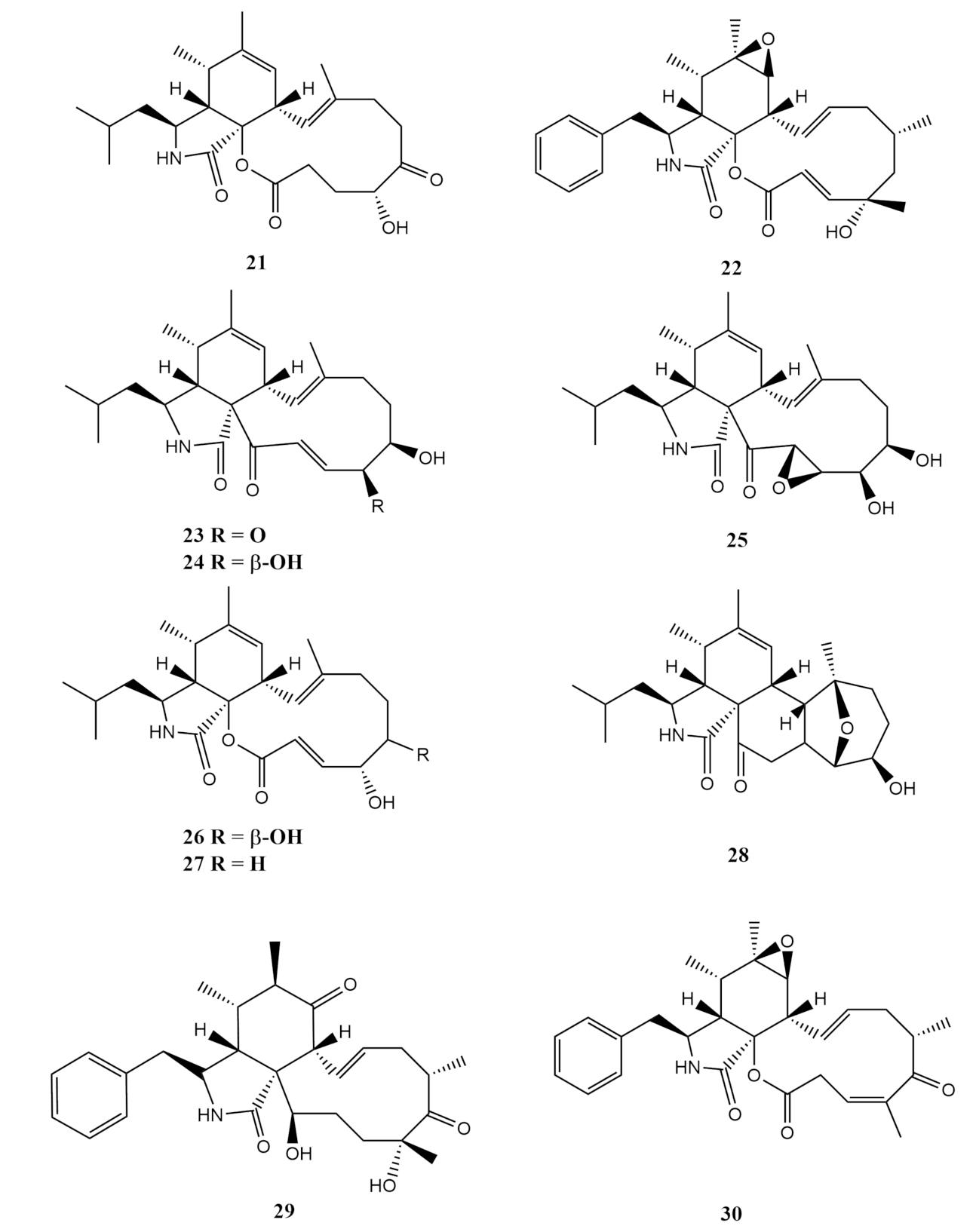
4.1. Terpenoids
Sesquiterpene
4.2. Alkaloids
Indole Type Alkaloid
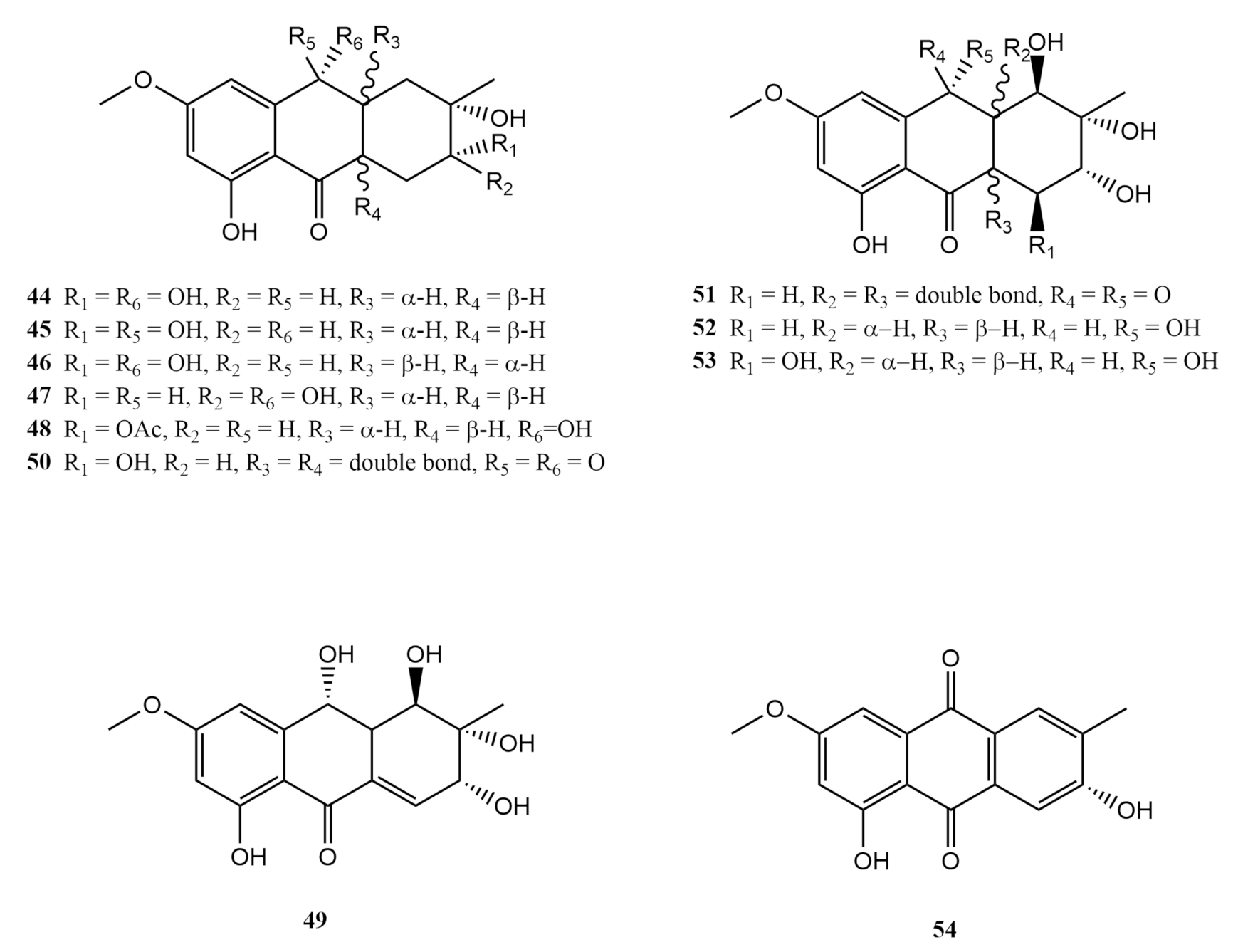
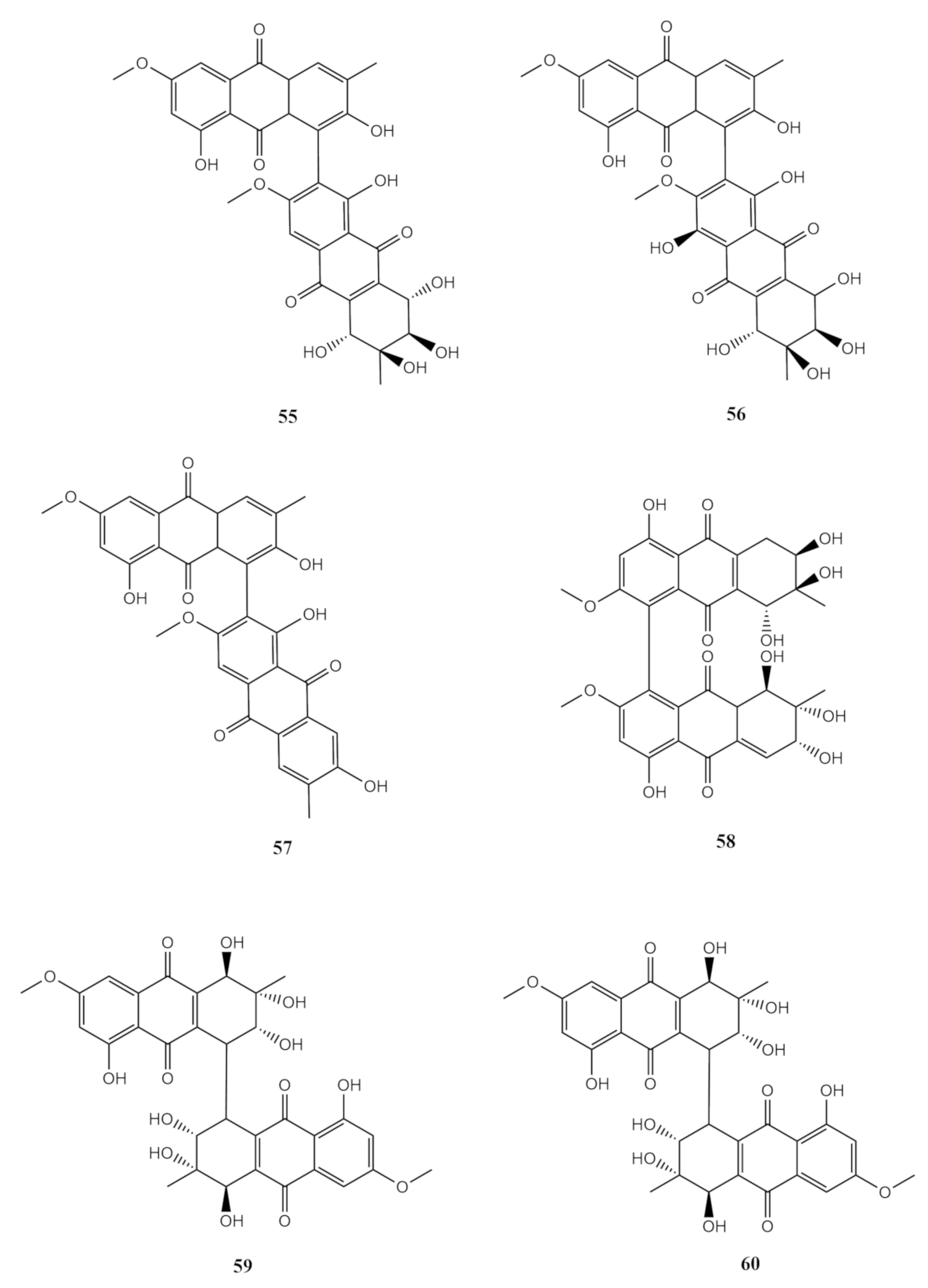
4.3. Anthraquinones Derivates
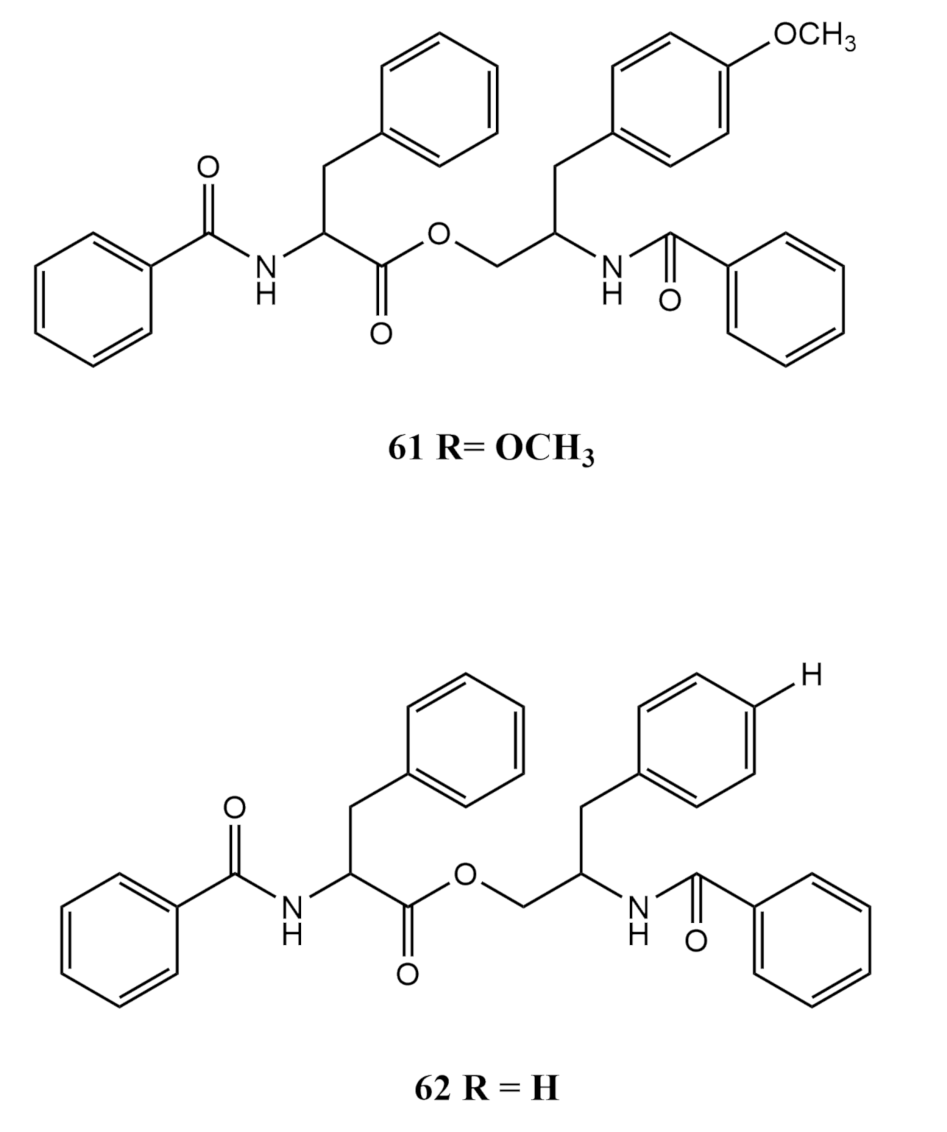
4.4. Amino Acid Derivates
4.5. Other Metabolites
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| UV | Ultra violet |
| MNP | Marine natural product |
| MAA | Mycosporin amino acid |
| Petr Eth | Petroleum Ether |
| EtOAc | Ethyl Acetate |
| MeOH | Methanol |
| RP-HPLC | Reverse Phased High-Performance Liquid Chromatography |
| HPLC | High-Performance Liquid Chromatography |
| NMR | Nuclear Magnetic Resonance |
| ROS | Reactive Oxygen Species |
| PDB | Potato Dextrose Broth |
| PDA | Potato Dextrose Agar |
| MeCN | Acetonitrile |
| ODS | Octadecylsilyl |
| A549 | Human lung adenocarcinoma cell line |
| CNE2 | Human nasopharyngeal carcinoma cell line |
| LoVo | Human colon cancer cell line |
| IC50 | Half maximal inhibition concentration |
| EC50 | Half maximal effective concentration |
| GPY | Glucose peptone yeast |
| COSY | Homonuclear correlation spectroscopy |
| HMBC | Heteronuclear Multiple Bond Correlation |
| SUNE1 | Human Nasopharyngeal carcinoma cell line |
| MCF-7 | Human breast cancer cell line |
| CNE1 | Human nasopharyngeal carcinoma cell line |
| Bel7402 | Human hepatic cancer cell line |
| KB | Human epidermoid carcinoma cell line |
| HONE1 | Epithelial tumour cell line |
| GLC-82 | Gejiu Lung Carcinoma-82 |
| SMMC-7721 | Human hepatocarcinoma cell line |
| HL7702 | Normal human liver cell |
| GLC82 | Human lung adenocarcinoma cell line |
| HL 7702 | Normal Human Liver Cells |
| HL-60 | Myeloid leukemic cell lines |
| Am-80 | RARα Specific Synthetic Retinoid |
| L5178Y | Mouse lymphoma cells |
| HCT-116 | Human colorectal carcinoma cell line |
| HeLa | Human cervical cancer cell line |
| PRRSV | porcine reproductive and respiratory syndrome virus |
| DEPT | Distortionless enhancement by polarisation transfer |
| HMBC | Heteronuclear multiple bond correlation |
| B16F10 | Human skin melanoma cell line |
| HeLa | Human cervical cancer cell line |
| HepG2 | Human liver cancer cell line |
| B16F10 | Human skin melanoma cell line |
| Vero | Normal kidney cells |
| HRESIMS | High-Resolution Electrospray Ionisation Mass Spectrometry |
| CHCl3 | Chloroform |
| OH | Hydroxyl |
| MIC | Minimum inhibition activity |
| LPS | Lipopolysaccharide |
| NO | Nitric oxide |
| HMGR-CoA | Hydroxymethylglutaryl-coenzyme A reductase |
| MPA | Mycophenolic acid |
| IMPDH2 | inosine-50-monophosphate dehydrogenase |
References
- Lau, Y.W.; Poliseno, A.; Kushida, Y.; Quere, G.; Reimer, J.D. The Classification, Diversity and Ecology of Shallow Water Octocorals. Encycl. World’s Biomes 2020, 4, 597–611. [Google Scholar]
- Yesson, C.; Taylor, M.L.; Tittensor, D.P.; Davies, A.J.; Guinotte, J.; Baco, A.; Black, J.; Hall-Spencer, J.M.; Rogers, A.D. Global habitat suitability of cold-water octocorals. J. Biogeogr. 2012, 39, 1278–1292. [Google Scholar] [CrossRef]
- Samimi-Namin, K.; van Ofwegen, L. The Octocoral Fauna of the Gulf. In Coral Reefs of the Gulf; Riegl, B.M., Purkis, S.J., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 225–252. [Google Scholar]
- Williams, G.C. The global diversity of sea pens (cnidaria: Octocorallia: Pennatulacea). PLoS ONE 2011, 6, e22747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabricius, K.; Alderslade, P. Soft Corals and Sea Fans: A Comprehensive Guide to the Tropical Shallow Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea; Fabricius, K., Alderslade, P., Eds.; Australian Institute of Marine Science: Queensland, Australia, 2001; p. 264. [Google Scholar]
- Zapata-Guardiola, R.; Lopez-Gonzalez, P.J. Two new gorgonian genera (Octocorallia: Primnoidae) from Southern Ocean waters. Polar Biol. 2010, 33, 313–320. [Google Scholar] [CrossRef]
- McFadden, C.S.; Ofwegen, L.P. A second, cryptic species of the soft coral genus Incrustatus (Anthozoa: Octocorallia: Clavulariidae) from Tierra del Fuego, Argentina, revealed by DNA barcoding. Helgol. Mar. Res. 2013, 67, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.D.; de Moura Neves, B.; Cordeiro, R.T.; Williams, G.C.; Cairns, S.D. Distribution of Octocorallia. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzlerland, 2016; pp. 109–123. [Google Scholar]
- McFadden, C.S.; Sanchez, J.A.; France, S.C. Molecular phylogenetic insights into the evolution of Octocorallia: A review. Integr. Comp. Biol. 2010, 50, 389–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadden, C.S.; van Ofwegen, L.P. Stoloniferous octocorals (Anthozoa, Octocorallia) from South Africa, with descriptions of a new family of Alcyonacea, a new genus of Clavulariidae, and a new species of Cornularia (Cornulariidae). Invertebr. Syst. 2012, 26, 331–356. [Google Scholar]
- McFadden, C.S.; France, S.C.; Sanchez, J.A.; Alderslade, P. A molecular phylogenetic analysis of the Octocorallia (Cnidaria: Anthozoa) based on mitochondrial protein-coding sequences. Mol. Phylogenet. Evol. 2006, 41, 513–527. [Google Scholar] [CrossRef]
- Aratake, S.; Tomura, T.; Saitoh, S.; Yokokura, R.; Kawanishi, Y.; Shinjo, R.; Reimer, J.D.; Tanaka, J.; Maekawa, H. Soft Coral Sarcophyton (Cnidaria: Anthozoa: Octocorallia) Species Diversity and Chemotypes. PLoS ONE 2012, 7, e30410. [Google Scholar] [CrossRef]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive secondary metabolites from octocoral-Associated microbes—New chances for blue growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [Green Version]
- Verselveldt, J. A revision of the genus Sarcophyton Lesson (octocorallia, Alcyonacea). Zool. Verh. 1982, 192, 1–91. [Google Scholar]
- Feussner, K.-D.; Waqa, T. Five new species of Sarcophyton (Coelenterata: Octocorallia) from the Fiji Islands. South Pac. J. Nat. Appl. Sci. 2013, 31, 1–26. [Google Scholar] [CrossRef]
- van der Land, J. UNESCO-IOC Register of Marine Organisms (URMO). 2008. Available online: http://www.marinespecies.org/urmo (accessed on 4 March 2021).
- Simpson, W.M. Descriptions of some of the new marine invertebrata from the Chinese and Japanese Seas. Acad. Nat. Sci. 1855, 7, 375–378. [Google Scholar]
- Verseveldt, J.; Benayahu, Y. Descriptions of one old and five new species of Alcyonacea (Coelenterata: Octocorallia) from the Red Sea. Zool. Meded. 1978, 53, 57–74. [Google Scholar]
- Verseveldt, J. Alcyonaceans (Coelenterata: Octocorallia) from some Micronesian Islands. Zool. Meded. 1978, 53, 49–55. [Google Scholar]
- Liu, R. Checklist of Marine Biota of China Seas. In Checklist of Marine Biota of China Seas; Liu, R., Ed.; Science Press: Beijing, China, 2008; p. 1267. [Google Scholar]
- Lin, M.-C.; Dai, C.-F. Drag, morphology and mechanical properties of three species of octocorals. J. Exp. Mar. Bio. Ecol. 1996, 201, 13–22. [Google Scholar] [CrossRef]
- Tixier-Durivault, A. Octocoralliaires de Madagascar et des iles avoisinantes. Faune de Madagascar 1966, 21, 1–456. [Google Scholar]
- Tixier-Durivault, A. Les octocoralliaires de Nha-Trang (Viet-Nam). Cah du Pacifique 1970, 14, 115–236. [Google Scholar]
- Imahara, Y. Report on the Octocorallia from the Ryukyu Islands of Japan. Bull. Inst. Ocean Res. Dev. Tokai Univ. 1991, 11, 59–94. [Google Scholar]
- Verseveldt, J. Octocorallia from north-western Madagascar (Part II). Zool. Verh. Leiden 1971, 117, 1–73. [Google Scholar]
- Roxas, H.A. Two new species of Sarcophyton Less. from the Philippines. Univ. Philipp. Nat. Appl. Sci. Bull. 1932, 4, 73–80. [Google Scholar]
- Verseveldt, J. Octocorallia from New Caledonia. Zool. Meded. 1974, 48, 95–122. [Google Scholar]
- Benayahu, Y. Corals of the South-West Indian Ocean: I. Alcyonacea from Sodwana Bay, South Africa; The Oceanographic Research Institute: Durban, South Africa, 1993; Volume 67, pp. 1–15. [Google Scholar]
- Macfadyen, L.M.I. Alcyonaria (Stolonifera, Alcyonacea, Telestacea and Gorgonacea). Gt Barrier Reef Exped 1928–1929. Sci. Rep. 1930, 5, 17. [Google Scholar]
- Utinomi, H. On some alcyonarians from the West-Pacific islands (Palau, Ponape and Bonins). Publ. Seto. Mar. Biol. Lab. 1956, 5, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Morton, B.; Morton, J. The Sea Shore Ecology of Hong Kong; Hong Kong University Press: Hong Kong, China, 1983. [Google Scholar]
- Roxas, H.A. Philippine Alcyonaria, II. The families Alcyoniidae and Nephthyidae. Philipp. J. Sci. 1933, 50, 1–5. [Google Scholar]
- van der Land, J. UNESCO-IOC Register of Marine Organisms, a common base for biodiversity inventories. In Families and Bibliography of Keyworks; van der Land, J., Ed.; NNM, Leiden and ETI: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Verseveldt, J. Report on the Octocorallia (Stolonifera and Alcyonacea) of the Israel south Red Sea expedition 1962, with notes on other collections from the Red Sea. Sea Fish Res. Stn Haifa Bull. 1965, 14, 27–47. [Google Scholar]
- Verseveldt, J.; Benayahu, Y. On two old and fourteen new species of Alcyonacea (Coelenterata, Octocorallia) from the Red Sea. Rijksmuseum van Natuurlijke Historie 1983, 208, 3–33. [Google Scholar]
- Verseveldt, J. Biological results of the Snellius expedition. XX. Octocorallia from the Malay Archipelago (Part I). Temminckia 1960, 10, 209–251. [Google Scholar]
- Verseveldt, J.; Tursch, A. Octocorallia from the Bismarck Sea (Part I). Zool. Meded. 1979, 54, 133–147. [Google Scholar]
- Samimi, N.K.; van Ofwegen, L.P. Some shallow water octocorals (Coelenterata: Anthozoa) of the Persian Gulf. Zootaxa 2009, 2058, 1–52. [Google Scholar]
- Benayahu, Y.; Perkol-finkel, S. Soft Corals (Octocorallia: Alcyonacea) from Southern Taiwan. I. Zool Stud. 2004, 43, 537–547. [Google Scholar]
- Pratt, E.M. The Alcyonaria of the Maldives. Part II. The genera Sarcophytum, Lobophytum, Sclerophytum and Alcyonium. Fauna Geogr. Maldive Laccadive Arch. 1903, 2, 503–509. [Google Scholar]
- Alderslade, P.; Shirwaiker, P. New species of soft corals (Coelenterata: Octocorallia) from the Laccadive Archipelago. Beagle 1991, 8, 189–233. [Google Scholar]
- Benayahu, Y.; Ofwegen, L.P. Van. New species of Sarcophyton and Lobophytum (Octocorallia: Alcyonacea) from Hong Kong. Zool. Meded. 2009, 83, 863–876. [Google Scholar]
- van de Water, J.A.J.M.; Allemand, D.; Ferrier-Pages, C. Host-microbe interactions in octocoral holobionts—Recent advances and perspectives. Microbiome 2018, 6, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Pernice, M.; Raina, J.B.; Radecker, N.; Cardenas, A.; Pogoreutz, C.; Voolstra, C.R. Down to the bone: The role of overlooked endolithic microbiomes in reef coral health. ISME J. 2020, 14, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, R.S.; Rosado, P.M.; Leite, D.C.D.A.; Rosado, A.S.; Bourne, D.G. Beneficial microorganisms for corals (BMC): Proposed mechanisms for coral health and resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef] [Green Version]
- Toledo-Hernandez, C.; Zuluaga-Montero, A.; Bones-Gonzalez, A.; Rodríguez, J.A.; Sabat, A.M.; Bayman, P. Fungi in healthy and diseased sea fans (Gorgonia ventalina): Is Aspergillus sydowii always the pathogen? Coral Reefs 2008, 27, 707–714. [Google Scholar] [CrossRef]
- Zuluaga-Montero, A.; Ramírez-Camejo, L.; Rauscher, J.; Bayman, P. Marine isolates of Aspergillus flavus: Denizens of the deep or lost at sea? Fungal Ecol. 2010, 3, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Soler-Hurtado, M.M.; Sandoval-Sierra, J.V.; Machordom, A.; Dieguez-Uribeondo, J. Aspergillus sydowii and other potential fungal pathogens in gorgonian octocorals of the Ecuadorian Pacific. PLoS ONE 2016, 11, e0165992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.J.; Shao, C.L.; Guo, Z.Y.; Chen, J.F.; Deng, D.S.; Yang, K.L.; Chen, Y.Y.; Fu, X.M.; She, Z.G.; Lin, Y.C. Bioactive hydroanthraquinones and anthraquinone dimers from a soft coral-derived Alternaria sp. fungus. J. Nat. Prod. 2012, 75, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.L.; Tan, T.K.; Chou, L.M.; Goh, N.K.C. In Proceedings of the The 12th International Coral Reef Symposium (ICRS 2012). Queensland, Australia, 9–13 July 2012; Volume 1, pp. 521–526. [Google Scholar]
- Zhang, X.-Y.; Bao, J.; Wang, G.-H.; He, F.; Xu, X.-Y.; Qi, S.-H. Diversity and antimicrobial activity of Culturable fungi isolated from six species of the South China Sea gorgonians. Microb. Ecol. 2012, 64, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, W.; Wang, J.; He, Y.; Zhang, J.; Li, F.; Zhang, Y. Bioactive secondary metabolites from the marine-associated fungus Aspergillus terreus. Bioorg. Chem. 2018, 80, 525–530. [Google Scholar] [CrossRef]
- El-Demerdash, A.; Kumla, D.; Kijjoa, A. Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update. Mar. Drugs 2020, 18, 317. [Google Scholar] [CrossRef]
- Li, H.-J.; Xie, Y.-L.; Xie, Z.-L.; Chen, Y.; Lam, C.-K.; Lan, W.-J. Chondrosterins A–E, Triquinane-Type Sesquiterpenoids from Soft Coral-Associated Fungus Chondrostereum sp. Mar. Drugs 2012, 10, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Lan, W.J.; Li, H.J. Two new hirsutane-type sesquiterpenoids chondrosterins N and O from the marine fungus Chondrostereum sp. Nat. Prod. Res 2018, 32, 1578–1582. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Hao, H.L.; Lau, S.C.K.; Wang, H.Y.; Han, Y.; Dong, L.M.; Huang, R.M. Biodiversity and antifouling activity of fungi associated with two soft corals from the South China Sea. Arch. Microbiol. 2019, 201, 757–767. [Google Scholar] [CrossRef]
- Agrawal, S.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. In-vitro evaluation of marine derived fungi against Cutibacterium acnes. Anaerobe 2018, 49, 5–13. [Google Scholar] [CrossRef]
- Zheng, C.J.; Shao, C.L.; Wu, L.Y.; Chen, M.; Wang, K.L.; Zhao, D.L.; Sun, X.P.; Chen, G.Y.; Wang, C.Y. Bioactive phenylalanine derivatives and cytochalasins from the soft coral-derived fungus, Aspergillus elegans. Mar. Drugs 2013, 11, 2054–2068. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.X.; Guo, Q.; Ran, Y.Q.; Qiu, Y.; Lan, W.J.; Li, H.J. New Aromadendrane Sesquiterpenoid Pseuboydone F from the Marine-derived Fungus Pseudallescheria boydii F44-1. Rec. Nat. Prod. 2019, 14, 166–170. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, C. Marine natural products in medicinal chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Changyun, W.; Haiyan, L.; Changlun, S.; Yanan, W.; Liang, L.; Huashi, G. Chemical defensive substances of soft corals and gorgonians. Acta Ecol. Sin. 2008, 28, 2320–2328. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the Coral Microbiome: Underpinning the Health and Resilience of Reef Ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Glasl, B.; Herndl, G.J.; Frade, P.R. The microbiome of coral surface mucus has a key role in mediating holobiont health and survival upon disturbance. ISME J. 2016, 10, 2280–2292. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hentschel, U.; Webster, N.; Olson, J.; Haggblom, M. Editorial: Special issue on sponge microbiome. FEMS Microbiol. Ecol. 2020, 96, fiaa075. [Google Scholar] [CrossRef]
- de la Coba, F.; Aguilera, J.; Korbee, N.; de Galvez, M.V.; Herrera-Ceballos, E.; Álvarez-Gomez, F. UVA and UVB Photoprotective Capabilities of Topical Formulations Containing Mycosporine-like Amino Acids (MAAs) through Different Biological Effective Protection Factors (BEPFs). Mar. Drugs 2019, 17, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-J.; Lan, W.-J.; Lam, C.-K.; Yang, F.; Zhu, X.-F. Hirsutane Sesquiterpenoids from the Marine-Derived Fungus Chondrostereum sp. Chem. Biodivers. 2011, 8, 317–324. [Google Scholar] [CrossRef]
- Li, H.J.; Chen, T.; Xie, Y.L.; Chen, W.D.; Zhu, X.F.; Lan, W.J. Isolation and structural elucidation of chondrosterins F–H from the marine fungus Chondrostereum sp. Mar. Drugs 2013, 11, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Li, H.J.; Jiang, W.H.; Liang, W.L.; Huang, J.X.; Mo, Y.F.; Ding, Y.Q.; Lam, C.K.; Qian, X.J.; Zhu, X.F.; Lan, W.J. Induced marine fungus Chondrostereum sp. as a means of producing new sesquiterpenoids chondrosterins I and J by using glycerol as the carbon source. Mar. Drugs 2014, 12, 167–175. [Google Scholar] [CrossRef]
- Huang, L.; Lan, W.J.; Deng, R.; Feng, G.K.; Xu, Q.Y.; Hu, Z.Y.; Zhu, X.F.; Li, H.J. Additional new cytotoxic triquinane-type sesquiterpenoids chondrosterins K–M from the marine fungus Chondrostereum sp. Mar. Drugs 2016, 14, 157. [Google Scholar] [CrossRef] [Green Version]
- Yagi, A.; Okamura, N.; Haraguchi, H.; Abot, T.; Hashimoto, K. Antimicrobial tetrahydroanthraquinones from a strain of Alternaria solani. Phytochemistry 1993, 33, 87–91. [Google Scholar] [CrossRef]
- Aly, A.H.; Edrada-Ebel, R.; Wray, V.; Muller, W.E.; Kozytska, S.; Hentschel, U.; Proksch, P.; Ebel, R. Bioactive metabolites from the endophytic fungus Ampelomyces sp. isolated from the medicinal plant Urospermum picroides. Phytochemistry 2008, 69, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Y.S.; Abrell, L.M.; Avelar, A.; Borgeson, B.M.; Crews, P. New hirsutane based sesquiterpenes from salt water cultures of a marine sponge-derived fungus and the terrestrial fungus Coriolus consors. Tetrahedron 1998, 54, 7335–7342. [Google Scholar] [CrossRef]
- Yang, F.; Chen, W.D.; Deng, R.; Li, D.D.; Wu, K.W.; Feng, G.K.; Zhu, X.F. Hirsutanol A induces apoptosis and autophagy via reactive oxygen species accumulation in breast cancer MCF-7 cells. J. Pharmacol. Sci. 2012, 119, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Hellwig, V.; Dasenbrock, J.; Schumann, S.; Steglich, W.; Leonhardt, K.; Anke, T. New triquinane-type sesquiterpenoids from Macrocystidia cucumis (basidiomycetes). Eur. J. Org. Chem. 1998, 1998, 73–79. [Google Scholar] [CrossRef]
- Etchri, A.; William, A.A.; Lois, M.B. Antifungal sesquiterpenoids from an arthroconidial fungus. J. Nat. Prod. 1989, 52, 1042–1054. [Google Scholar]
- Gebhardt, K.; Schimana, J.; Hoeltzel, A.; Dettner, K.; Draeger, S.; Beil, W.; Rheinheimer, J.; Fiedler, H.P. Aspochalamins AD and Aspochalasin Z Produced by the Endosymbiotic Fungus Aspergillus niveus LU 9575 II. Structure Elucidation. J. Antibiot. 2004, 57, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, T.; Shin-Ya, K.; Seto, H.; Okusa, N.; Kajiura, T.; Hayakawa, Y. Structure of aspochalasin H, a new member of the aspochalasin family. J. Antibiot. 2002, 55, 666–668. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.X.; Wijeratne, E.K.; Bigelow, D.; Pierson, L.S.; VanEtten, H.D.; Gunatilaka, A.L. Aspochalasins I, J, and K: Three new cytotoxic cytochalasans of Aspergillus f lavipes from the rhizosphere of Ericameria laricifolia of the Sonoran Desert. J. Nat. Prod. 2004, 67, 328–332. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, T.; Pei, Y.; Hua, H.; Feng, B. Aspergillin PZ, a novel isoindole-alkaloid from Aspergillus awamori. J. Antibiot. 2002, 55, 693–695. [Google Scholar] [CrossRef] [Green Version]
- Fujii, Y.; Tani, H.; Ichinoe, M.; Nakajima, H. Zygosporin D and two new cytochalasins produced by the fungus Metarrhizium anisopliae. J. Nat. Prod. 2000, 63, 132–135. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, H.; Wu, N.; Liu, M.; Wei, J.; Zhang, Y.; Lin, X. Characterization of the high cytochalasin E and rosellichalasin producing-Aspergillus sp. nov. F1 isolated from marine solar saltern in China. World J. Microbiol. Biotechnol. 2013, 29, 11–17. [Google Scholar] [CrossRef]
- Veluri, R.; Oka, I.; Wagner-Dobler, I.; Laatsch, H. New indole alkaloids from the North Sea bacterium Vibrio parahaemolyticus Bio249. J. Nat. Prod. 2003, 66, 1520–1523. [Google Scholar] [CrossRef]
- Cai, S.X.; Li, D.H.; Zhu, T.J.; Wang, F.P.; Xiao, X.; Gu, Q.Q. Two New Indole Alkaloids from the Marine-Derived Bacterium Aeromonas sp. CB101. Helv. Chim. Acta 2010, 93, 791–795. [Google Scholar] [CrossRef]
- Yang, C.L.; Han, Y.; Wang, Y.; Zhang, X.H.; Zhu, W.M. Bis- and tris- indole alkaloids from Edwardsiella tarda. Microbiol. China 2010, 37, 1325–1330. [Google Scholar]
- Yuan, M.-X.; Qiu, Y.; Ran, Y.-Q.; Feng, G.-K.; Deng, R.; Zhu, X.-F. Exploration of Indole Alkaloids from Marine Fungus Pseudallescheria boydii F44-1 Using an Amino Acid-Directed Strategy. Mar. Drugs 2019, 17, 77. [Google Scholar] [CrossRef] [Green Version]
- Liew, L.P.; Fleming, J.M.; Longeon, A.; Mouray, E.; Florent, I.; Bourguet-Kondracki, M.L.; Copp, B.R. Synthesis of 1-indolyl substituted β-carboline natural products and discovery of antimalarial and cytotoxic activities. Tetrahedron 2014, 70, 4910–4920. [Google Scholar] [CrossRef]
- Machowinski, A.; Kramer, H.J.; Hort, W.; Mayser, P. Pityriacitrin—A potent UV filter produced by Malassezia furfur and its effect on human skin microflora. Mycoses 2006, 49, 388–392. [Google Scholar] [CrossRef]
- Jiao, W.H.; Gao, H.; Li, C.Y.; Zhou, G.X.; Kitanaka, S.; Ohmura, A.; Yao, X.S. β-Carboline alkaloids from the stems of Picrasma quassioides. Magn. Reson. Chem. 2010, 48, 490–495. [Google Scholar] [PubMed]
- Anouhe, J.B.S.; Adima, A.A.; Niamke, F.B.; Stien, D.; Amian, B.K.; Blandinieres, P.A.; Virieux, D.; Pirat, J.L.; Kati-Coulibaly, S.; Amusant, N. Dicorynamine and harmalan-N-oxide, two new β-carboline alkaloids from Dicorynia guianensis Amsh heartwood. Phytochem. Lett. 2010, 12, 158–163. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.F.; Kim, D.S.; Choi, H.D.; Son, B.W. Indolyl alkaloid derivatives, N b-acetyltryptamine and oxaline from a marine-derived fungus. Arch. Pharm. Res. 2003, 26, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Edrada-Ebel, R.; Wray, V.; Muller, W.E.; Totzke, F.; Zirrgiebel, U.; Schachtele, C.; Kubbutat, M.H.; Lin, W.H. Bioactive metabolites from the endophytic fungus Stemphylium globuliferum isolated from Mentha pulegium. J. Nat. Prod. 2009, 72, 626–631. [Google Scholar] [CrossRef]
- Suemitsu, R.; Ueshima, T.; Yamamoto, T.; Yanagawase, S. Alterporriol C: A modified bianthraquinone from Alternaria porri. Phytochemistry 1988, 27, 3251–3254. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, J.; Liu, P.; Wang, W.; Zhu, W. Three new compounds from Aspergillus terreus PT06-2 grown in a high salt medium. Mar. Drugs 2011, 9, 1368–1378. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Z.; Wei, H.J.; Zhu, T.J.; Li, D.H.; Lin, Z.J.; Gu, Q.Q. Three New Cytochalasins from the Marine-Derived Fungus Spicaria elegans KLA03 by Supplementing the Cultures with L-and D-Tryptophan. Chem. Biodivers. 2011, 8, 887–894. [Google Scholar] [CrossRef]
- Bladon, P.; Sleigh, T. Photo-oxygenation of 3-acetoxyergosta-3,5,7,22-tetraene and related compounds. J. Chem. Soc. 1965, 6991–7000. [Google Scholar] [CrossRef]
- Nong, X.H.; Wang, Y.F.; Zhang, X.Y.; Zhou, M.P.; Xu, X.Y.; Qi, S.H. Territrem and butyrolactone derivatives from a marine-derived fungus aspergillus terreus. Mar. Drugs 2014, 12, 6113–6124. [Google Scholar] [CrossRef]
- Phainuphong, P.; Rukachaisirikul, V.; Saithong, S.; Phongpaichit, S.; Bowornwiriyapan, K.; Muanprasat, C. Lovastatin Analogues from the Soil-Derived Fungus Aspergillus sclerotiorum PSU-RSPG178. J. Nat. Prod. 2016, 79, 1500–1507. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Ye, Q.; Li, J.; Hua, Y.; Ju, D. Constituents of red yeast rice, a traditional Chinese food and medicine. J. Agric. Food Chem. 2000, 48, 5220–5225. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, B.; Li, F.; Liu, M.; Lin, S.; Wang, J. Mycophenolic acid derivatives with immunosuppressive activity from the coral-derived fungus penicillium bialowiezense. Mar. Drugs 2018, 16, 230. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Region | Reference |
|---|---|---|
| Sarcophyton aalbersbergi Feussner and Waqa, 2013 | Fiji Island | [15] |
| Sarcophyton acutum Tixier-Durivault, 1970 | Western Central Pacific | [16] |
| Sarcophyton agaricum (Stimpson, 1855) | Chinese, Japanese Sea | [17] |
| Sarcophyton aldersladei Feussner and Waqa, 2013 | Fiji Islands | [15] |
| Sarcophyton alexanderi Feussner and Waqa, 2013 | Fiji Islands | [15] |
| Sarcophyton auritum Verseveldt and Benayahu, 1978 | Red Sea | [18] |
| Sarcophyton birkelandi Verseveldt, 1978 | Micronesian Islands | [19] |
| Sarcophyton boettgeri Schenk, 1896 | Indonesia | [14] |
| Sarcophyton boletiforme Tixier-Durivault, 1958 | China seas | [20] |
| Sarcophyton buitendijki Verseveldt, 1982 | Southern Taiwan | [21] |
| Sarcophyton cherbonnieri Tixier-Durivault, 1958 | Madagascar | [22] |
| Sarcophyton cinereum Tixier-Durivault, 1946 | Madagascar, Vietnam, Japan | [22,23,24] |
| Sarcophyton cornispiculatum Verseveldt, 1971 | Madagascar, China Seas | [20,25] |
| Sarcophyton crassocaule Moser, 1919 | Philippines, Madagascar, Vietnam, China seas | [20,22,23,26] |
| Sarcophyton crassum Tixier-Durivault, 1946 | Madagascar, New Caledonia, South-West Indian Ocean | [22,27,28] |
| Sarcophyton digitatum Moser, 1919 | Philippines, Great Barrier Reef, West-Pacific islands, Madagascar, New Caledonia | [22,23,26,29,30] |
| Sarcophyton ehrenbergi von Marenzeller, 1886 | Philippines, Madagascar, Vietnam, China seas | [20,22,23] |
| Sarcophyton elegans Moser, 1919 | Philippines, Madagascar, New Caledonia, Hong Kong, Japan, China seas | [20,22,23,24,31,32] |
| Sarcophyton expandum Kolliker | Samoa Islands, South Pacific Ocean | [33] |
| Sarcophyton flexuosum Tixier-Durivault, 1966 | Madagascar, South-West Indian Ocean | [22,28] |
| Sarcophyton furcatum Li, 1984 | China Seas | [20] |
| Sarcophyton gemmatum Verseveldt and Benayahu, 1978 | Red Sea | [18] |
| Sarcophyton glaucum (Quoy and Gaimard, 1833) | Philippines, Madagascar, Red Sea, West-Pacific islands, China seas | [20,22,30,32,34] |
| Sarcophyton globoverruccatum Benayahu and Verseveldt, 1983 | Red Sea | [35] |
| Sarcophyton griffini Moser, 1919 | Papua New Guinea | [33] |
| Sarcophyton infundibuliforme Tixier-Durivault, 1958 | Madagascar, New Caledonia, South-West Indian ocean, China seas | [20,22,23,28] |
| Sarcophyton latum (Dana, 1846) | Philippines, Malay Archipelago, Madagascar, China Seas | [20,22,32,36] |
| Sarcophyton mililatensis Verseveldt and Tursch, 1979 | Bismarck Sea | [37] |
| Sarcophyton minusculum Samimi Namin and van Ofwegen, 2009 | Persian Gulf | [38] |
| Sarcophyton nanwanensis Benayahu and Perkol-Finkel, 2004 | southern Taiwan | [39] |
| Sarcophyton nigrum May, 1899 | Marshall Islands, North Pacific Ocean | [33] |
| Sarcophyton pauciplicatum Verseveldt and Benayahu, 1978 | Red Sea | [18] |
| Sarcophyton portentosum Tixier-Durivault, 1970 | New Caledonia | [22] |
| Sarcophyton pulchellum (Tixier-Durivault, 1957) | Indian Waters, Japan | [33] |
| Sarcophyton regulare Tixier-Durivault, 1946 | Madagascar, New Caledonia | [22,23] |
| Sarcophyton roseum Pratt, 1903 | Maldives | [40] |
| Sarcophyton serenei Tixier-Durivault, 1958 | Vietnam | [23] |
| Sarcophyton skeltoni Feussner and Waqa, 2013 | Fiji Islands | [15] |
| Sarcophyton soapiae Feussner and Waqa, 2013 | Fiji Islands | [15] |
| Sarcophyton solidum Tixier-Durivault, 1958 | Madagascar | [22] |
| Sarcophyton spinospiculatum Alderslade and Shirwaiker, 1991 | Laccadive Archipelago | [41] |
| Sarcophyton spongiosum Thomson and Dean, 1931 | Malay Archipelago, Madagascar | [22,36] |
| Sarcophyton stellatum Kükenthal, 1911 | China Seas | [20] |
| Sarcophyton stolidotum Verseveldt, 1971 | Madagascar | [25] |
| Sarcophyton subviride Tixier-Durivault, 1958 | Madagascar | [22] |
| Sarcophyton tenuispiculatum Thomson and Dean, 1931 | West-Pacific islands, Malay Archipelago, New Caledonia | [23,30,36] |
| Sarcophyton tortuosum Tixier-Durivault, 1946 | [33] | |
| Sarcophyton trocheliophorum von Marenzeller, 1886 | Philippines, West-Pacific islands, Malay Archipelago, Madagascar, New Caledonia, Vietnam, Japan, China seas | [20,22,23,24,30,32,36] |
| Sarcophyton tumulosum Benayahu and van Ofwegen, 2009 | Hong Kong | [42] |
| Sarcophyton turschi Verseveldt, 1976 | Red Sea | [35] |
| Soft Coral Species | Fungi | Reference |
|---|---|---|
| Sarcophyton subviride | Aspergillus terreus | [52] |
| Penicillium bialowiezense | [53] | |
| Sarcophyton tortuosum | Chondrostereum sp. | [54,55] |
| Alternaria alternata | ||
| Aspergillus versicolor | ||
| Chaunopycnis sp. | ||
| Cladosporium cladosporioides | ||
| Cladosporium dominicanum | ||
| Cladosporium sphaerospermum | ||
| Didymella sp. | ||
| Hypocrea lixii | ||
| Microsphaeropsis sp. | ||
| Paraconiothyrium cyclothyrioides | [56] | |
| Penicillium citrinum | ||
| Tritirachium sp. | ||
| Penicillium janthinellum | ||
| Penicillium oxalicum | ||
| Phoma putaminum | ||
| Phoma sp. | ||
| Pseudocercospora sp. | ||
| Stagonosporopsis cucurbitacearum | ||
| Talaromyces allahabadensis | ||
| Sarcophyton sp. | Aspergillus elegans | [57,58] |
| Pseudallescheria boydii | [59] |
| Soft Coral Species | Fungi | Metabolites | Bioactivities | Reference |
|---|---|---|---|---|
| Sarcophyton tortuosum | Chondrostereum sp. | chondrosterin A (5) | cytotoxic activities against cancer lines A549, CNE2, and LoVo | [54] |
| hirsutanol A (1) | potent cytotoxic activities against various cancer cell lines | [67] | ||
| incarnal (11) | potent cytotoxic activity against various cancer cell lines | [68] | ||
| chondrosterin J (14) | potent cytotoxic activities against the cancer cell lines CNE-1 and CNE-2 | [69] | ||
| chondrosterin K (15) | ||||
| chondrosterins L (16) | significant cytotoxicity against various cancer cell lines in vitro | [70] | ||
| chondrosterins M (17) | ||||
| Sarcophyton sp. | Pseudallescheria boydii | 3,3′-cyclohexylidenebis(1H-indole) (33) | significant cytotoxic activity against various cancer cell lines | [59] |
| Aspergillus elegans | 4′-OMe-asperphenamate (61) | antibacterial activity against Staphylococcus epidermidis | [57] | |
| Alternaria sp. | altersolanol B (50) | potent inhibitory activity against Gram-negative bacteria | [71] | |
| altersolanol C (51) | ||||
| Alternaria sp. | ampelanol (53) | mild toxicity against the L5178Y mouse lymphoma cells | [72] | |
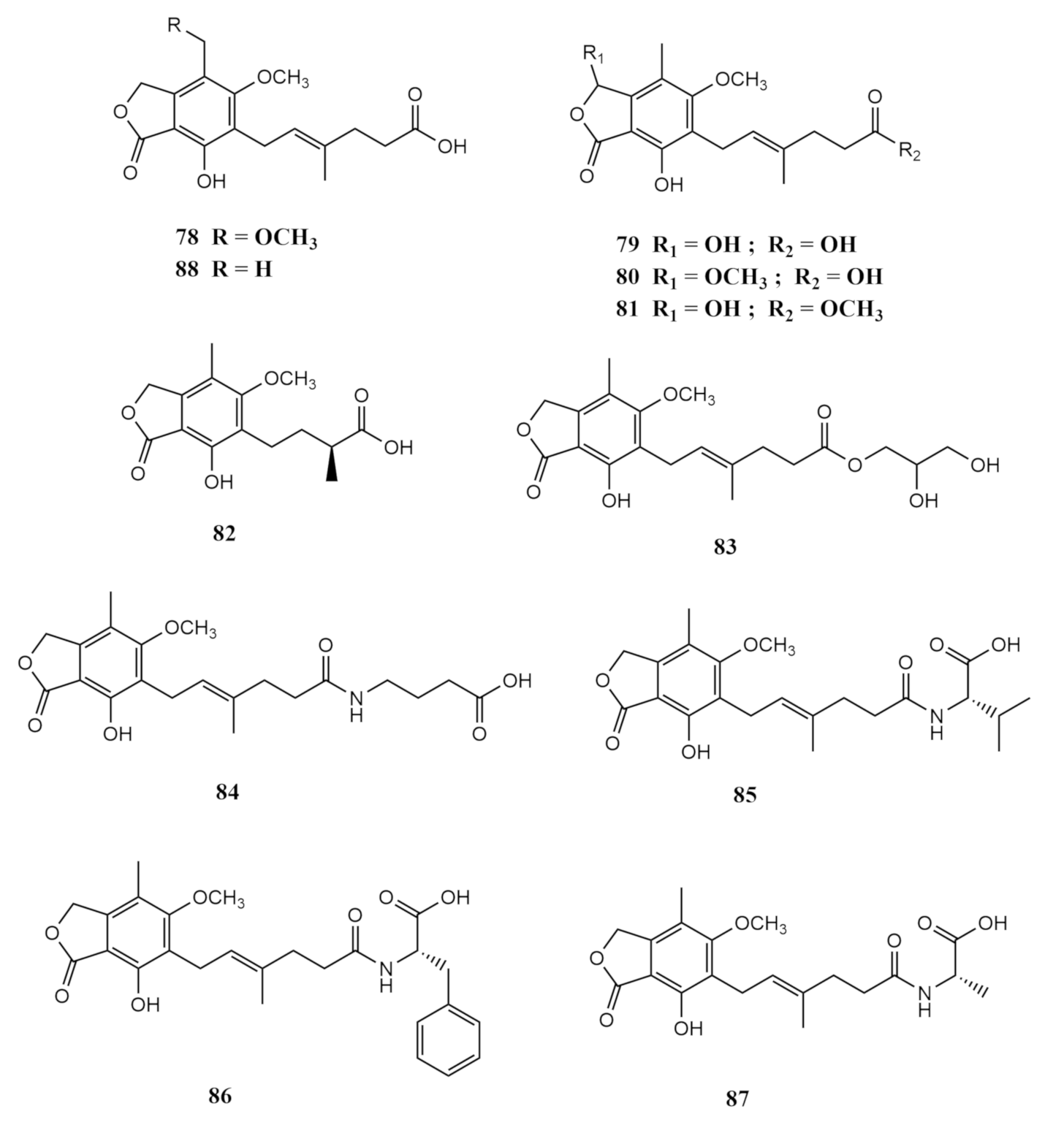
| Sarcophyton subviride | Aspergillus terreus | versicolactone G (67) | potent α-glucosidase inhibitory activity | [52] |
| luteoride E (66) | ||||
| (3E,7E)-4,8-dimethyl-undecane-3,7-diene-1,11-diol (68) | ||||
| methyl 3,4,5-trimethoxy-2-(2 -(nicotinamido)benzamido)benzoate (70) | significant anti-inflammatory activity against NO production | [52] | ||
| territrem A (72) | ||||
| lovastatin (75) | ||||
| Penicillium bialowiezense | 8-O-methyl mycophenolic acid (78) | |||
| 3-hydroxy mycophenolic acid (79) | ||||
| 6-(5-carboxy-3-methylpent-2-enyl)-7-hydroxy-3,5-dimethoxy-4-methylphthalan-1-one (80) | inhibitory activity against inosine-50-monophosphate dehydrogenase (IMPDH2) | |||
| 6-(5-methoxycarbonyl-3-methylpent-2-enyl)-3,7-dihydroxy-5-methoxy-4-methylphthalan-1-one (81) | ||||
| 6-(3-carboxybutyl)-7-hydroxy-5-methoxy-4-methylphthalan-1-one (82) | and | [53] | ||
| 6-[5-(2,3-dihydroxy-l-carboxyglyceride)-3-methylpent-2-enyl]-7-hydroxy-5-methoxy-4-methylphthalan-1-one (83) | in vitro immunosuppressive activity against the proliferation of T-lymphocytes | |||
| 6-[5-(1-carboxy-4-N-carboxylate)-3-methylpent-2-enyl]-7-hydroxy-5-methoxy-4-methylphthalan-1-one (84) | ||||
| N-mycophenoyl-l-valine (85) | ||||
| N-mycophenoyl–l-phenyloalanine (86) | ||||
| N-mycophenoyl–l-alanine (87) | ||||
| mycophenolic acid (MPA) (88) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Palaniveloo, K.; Alias, S.A.; Sathiya Seelan, J.S. Species Diversity and Secondary Metabolites of Sarcophyton-Associated Marine Fungi. Molecules 2021, 26, 3227. https://doi.org/10.3390/molecules26113227
Liu Y, Palaniveloo K, Alias SA, Sathiya Seelan JS. Species Diversity and Secondary Metabolites of Sarcophyton-Associated Marine Fungi. Molecules. 2021; 26(11):3227. https://doi.org/10.3390/molecules26113227
Chicago/Turabian StyleLiu, Yuanwei, Kishneth Palaniveloo, Siti Aisyah Alias, and Jaya Seelan Sathiya Seelan. 2021. "Species Diversity and Secondary Metabolites of Sarcophyton-Associated Marine Fungi" Molecules 26, no. 11: 3227. https://doi.org/10.3390/molecules26113227