
Brassica incana Ten. (Brassicaceae): Phenolic Constituents, Antioxidant and Cytotoxic Properties of the Leaf and Flowering Top Extracts

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Phytochemical Investigations
2.1.1. Determination of Total Phenolic Content
2.1.2. Identification of Phenolic Compounds by HPLC-PDA-ESI-MS
2.2. Antioxidant Activity
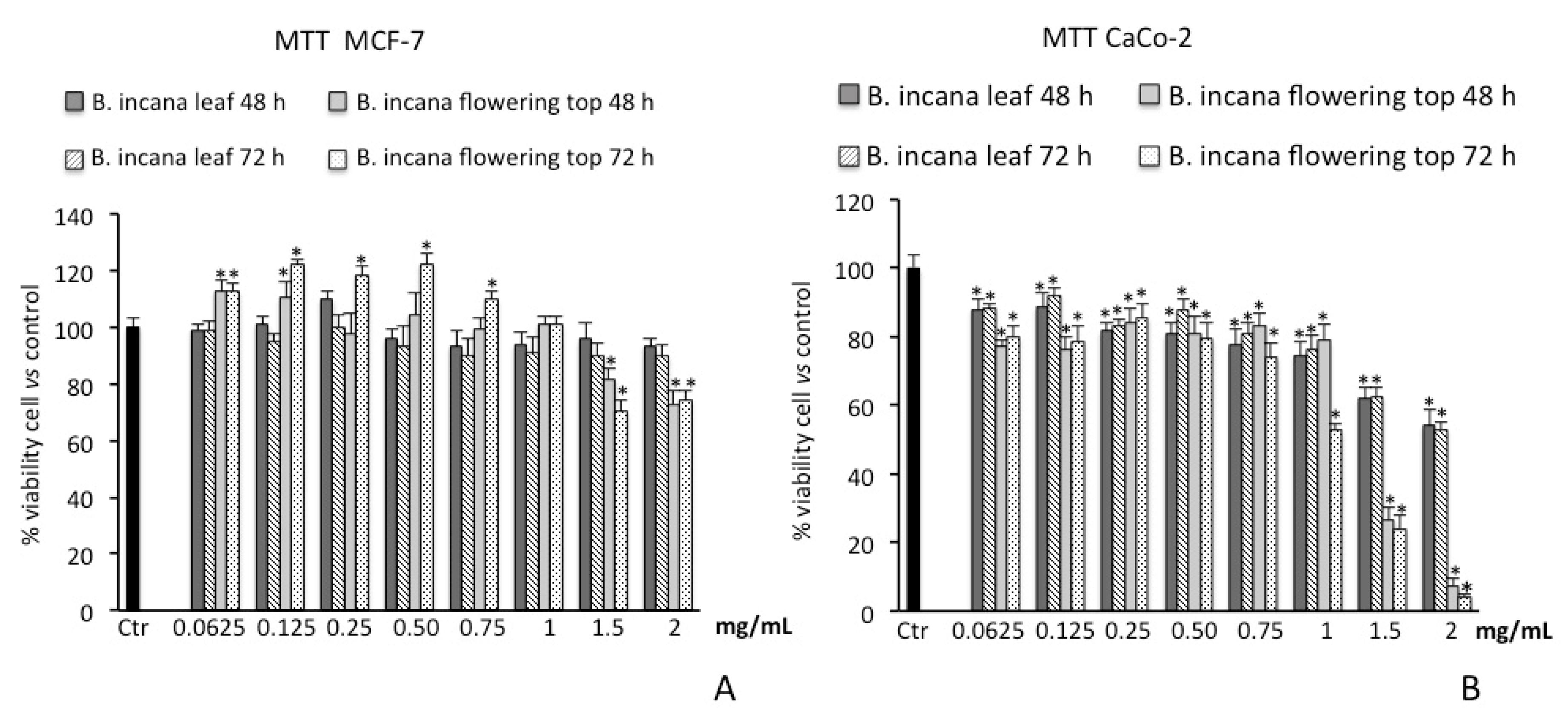
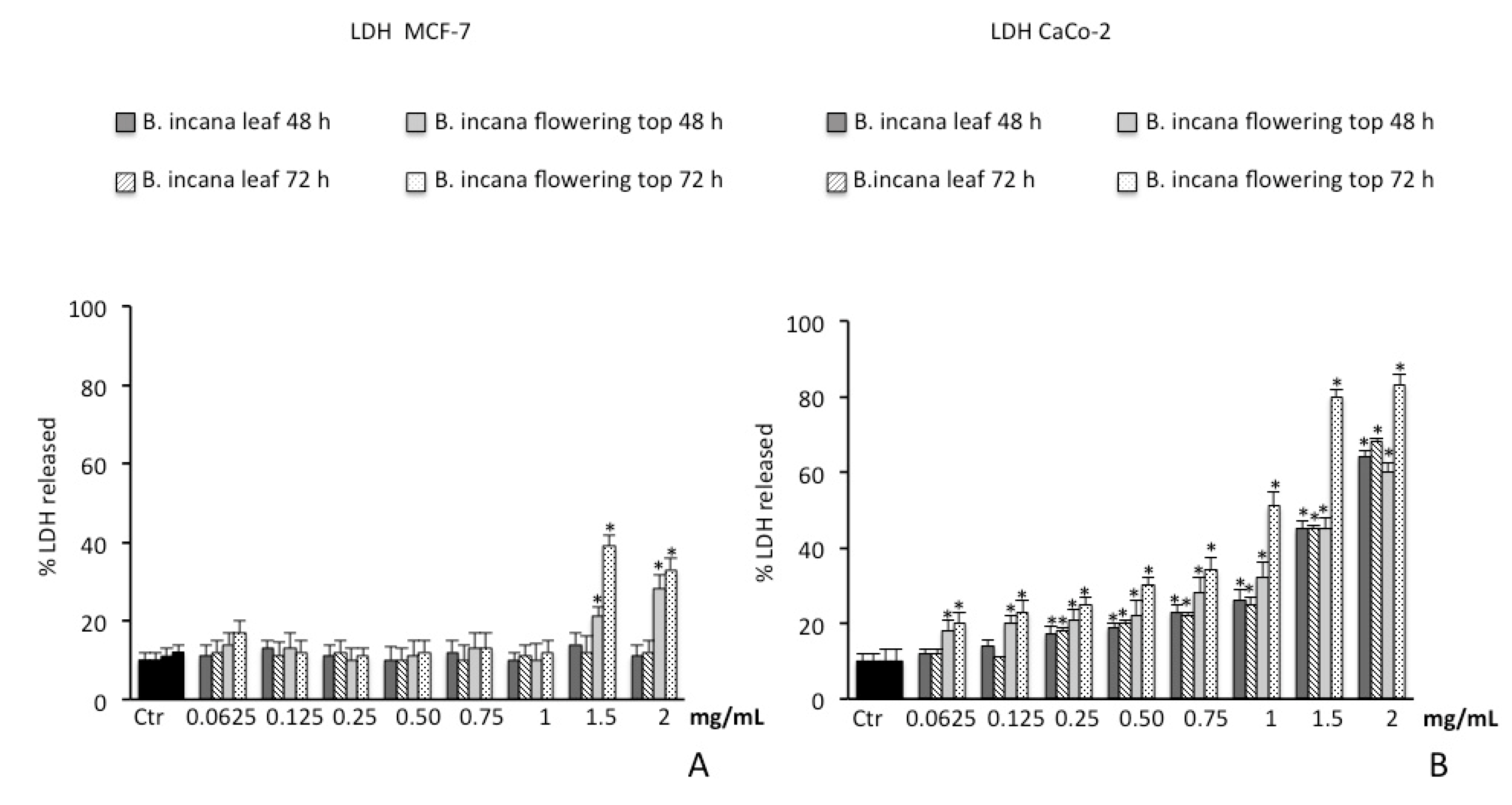
2.3. Cytotoxic Activity
2.3.1. Cell Viability Assay on Human Colorectal Adenocarcinoma (CaCo-2) and Breast Cancer (MCF-7) Cells
2.3.2. Brine Shrimp Lethality Bioassay
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Extraction Procedure
4.3. Phytochemical Investigations
4.3.1. Determination of Total Phenolic Content
4.3.2. Identification of Phenolic Compounds by HPLC-PDA-ESI-MS
4.4. Antioxidant Activity
4.4.1. Free Radical Scavenging Activity
4.4.2. Measurement of Reducing Power
4.4.3. Ferrous Ion (Fe2+) Chelating Activity
4.4.4. Protective Effect on Escherichia coli Growth and Survival under Peroxide Stress
4.5. Cytotoxic Activity
4.5.1. Cell viability Assay on Human Colorectal Adenocarcinoma (CaCo-2) and Breast Cancer (MCF-7) Cells
4.5.2. Brine Shrimp Lethality Bioassay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Katche, E.; Quezada-Martinez, D.; Katche, E.I.; Vasquez-Teuber, P.; Mason, A.S. Interspecific hybridization for Brassica crop improvement. Crop Breed. Genet. Genom. 2019, 1, e190007. [Google Scholar] [CrossRef] [Green Version]
- Rakow, G. Species origin and economic importance of Brassica. In Brassica. Biotechnology in Agriculture and Forestry; Pua, E.C., Douglas, C.J., Eds.; Springer: Berlin, Germany, 2004; Volume 54, pp. 3–11. ISBN 978-3-662-06164-0. [Google Scholar]
- Branca, F.; Cartea, E. Brassica. In Wild Crop Relatives: Genomic and Breeding Resources, Oilseeds; Kole, C., Ed.; Springer-Verlag: Berlin, Germany, 2011; pp. 17–36. ISBN 978-3-642-14871-2. [Google Scholar]
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–43. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Miceli, N.; Filocamo, A.; Ragusa, S.; Cacciola, F.; Dugo, P.; Mondello, L.; Celano, M.; Maggisano, V.; Taviano, M.F. Chemical characterization and biological activities of phenolic-rich fraction from cauline leaves of Isatis tinctoria L. (Brassicaceae) growing in Sicily, Italy. Chem. Biodivers. 2017, 14, e1700073. [Google Scholar] [CrossRef]
- Taviano, M.F.; Filocamo, A.; Ragusa, S.; Cacciola, F.; Dugo, P.; Mondello, L.; Paterniti Mastrazzo, G.; De Rose, R.F.; Celano, M.; Lombardo, G.E.; et al. Phenolic profile, antioxidant and cytotoxic properties of polar extracts from leaves and flowers of Isatis tinctoria L. (Brassicaceae) growing in Sicily. Plant Biosyst. 2018, 152, 795–803. [Google Scholar] [CrossRef]
- Miceli, N.; Cavò, E.; Ragusa, S.; Cacciola, F.; Dugo, P.; Mondello, L.; Marino, A.; Cincotta, F.; Condurso, C.; Taviano, M.F. Phytochemical characterization and biological activities of a hydroalcoholic extract obtained from the aerial parts of Matthiola incana (L.) R. Br. Subsp. Incana (Brassicaceae) growing wild in Sicily (Italy). Chem. Biodivers. 2019, 16, e1800677. [Google Scholar] [CrossRef]
- Heywood, V.H. Brassica L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge at the University Press: Cambridge, UK, 1964; Volume I, pp. 335–339. [Google Scholar]
- Heywood, V.H.; Zohary, D. A catalogue of the wild relatives of cultivated plants native to Europe. Flora Mediterr. 1995, 5, 375–415. [Google Scholar]
- Marhold, K. Brassicaceae. Euro+Med Plantbase-the Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://www.emplantbase.org/home.html (accessed on 1 February 2020).
- Castellano, G.; Bazan, G. Aspetti ӱistributive e fitosociologici di Brassica incana (Brassicaceae, Magnoliophyta) in Sicilia. Quad. Bot. Amb. Appl. 2009, 20, 263–268. [Google Scholar]
- Pignatti, S. Brassica L. In Flora d’Italia; Edagricole-New Business Media, Ed.; Edagricole: Milano, Italy, 2017; Volume 2, pp. 1016–1028. ISBN 8850652437. [Google Scholar]
- Do Nurb, A. Piante Spontanee d’uso Alimentare-Riconoscere, Raccogliere; Rifletto & Rifrango, Ed.; Lulu.com®: Rome, Italy, 2018; pp. 87–89. ISBN 9780244445331. [Google Scholar]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Kumar, S.; Andy, A. Health promoting bioactive phytochemicals from Brassica. Int. Food Res. J. 2012, 19, 141–152. [Google Scholar]
- Sobrinho Santos, E.M.; Almeida, A.C.; Santos, H.O.; Cangussu, A.R.; Costa, K.S.; Alves, J.N.; Bertucci Barbosa, L.C.; Souza Aguiar, R.W. Mechanism of Brassica oleracea performance in bovine infectious mastitis by bioinformatic analysis. Microb. Pathog. 2019, 129, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, G.; Verzera, A.; Dima, G.; Condurso, C.; Ragusa, S. Brassica fruticulosa Cyr. and Brassica incana Ten. (Brassicaceae) as Mediterranean traditional wild vegetables: A valuable source of bioactive compounds. J. Essent. Oil Res. 2012, 24, 539–545. [Google Scholar] [CrossRef]
- Horn, P.J.; Vaughan, J.G. Seed glucosinolates of fourteen wild Brassica species. Phytochemistry 1983, 22, 465–471. [Google Scholar] [CrossRef]
- Heaney, R.K.; Fenwick, G.R.; Mithen, R.F.; Lewis, B.G. Glucosinolates of wild and cultivated Brassica species. Phytochemistry 1987, 26, 1969–1973. [Google Scholar] [CrossRef]
- Velasco, L.; Becker, H.C. Variability for seed glucosinolates in a germplasm collection of the genus Brassica. Genet. Resour. Crop Evol. 2000, 47, 231–238. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT-Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. A comparative study on the polyphenolic content, antibacterial activity and antioxidant capacity of different solvent extracts of Brassica oleracea vegetables. Int. J. Food Sci. Technol. 2012, 47, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Ferreres, F.; Sousa, C.; Vrchovská, V.; Valentão, P.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Chemical composition and antioxidant activity of tronchuda cabbage internal leaves. Eur. Food Res. Technol. 2006, 222, 88–98. [Google Scholar] [CrossRef]
- Zietz, M.; Weckmüller, A.; Schmidt, S.; Rohn, S.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Genotypic and climatic influence on the antioxidant activity of flavonoids in Kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2010, 58, 2123–2130. [Google Scholar] [CrossRef]
- Fiol, M.; Adermann, S.; Neugart, S.; Rohn, S.; Mügge, C.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Highly glycosylated and acylatedflavonols isolated from kale (Brassica oleracea var. sabellica)—Structure–antioxidant activity relationship. Food Res. Int. 2012, 47, 80–89. [Google Scholar] [CrossRef]
- Nićiforović, N.; Abramovič, H. Sinapic acid and its derivatives: Natural sources and bioactivity. Compr. Rev. Food Sci. Food Saf. 2014, 13, 34–51. [Google Scholar] [CrossRef]
- Li, Z.; Lee, H.W.; Liang, X.; Liang, D.; Wang, Q.; Huang, D.; Ong, C.N. Profiling of phenolic compounds and antioxidant activity of 12 cruciferous vegetables. Molecules 2018, 23, E1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenbaek, M.; Tybirk, E.; Neugart, S.; Sundekilde, U.K.; Schreiner, M.; Kristensen, H.L. Flavonoid glycosides and hydroxycinnamic acid derivatives in baby leaf rapeseed from white and yellow flowering cultivars with repeated harvest in a 2-years field study. Front. Plant Sci. 2019, 10, 355. [Google Scholar] [CrossRef]
- Plumb, G.W.; Price, K.R.; Rhodes, M.J.; Williamson, G. Antioxidant properties of the major polyphenolic compounds in broccoli. Free Radic. Res. 1997, 27, 429–435. [Google Scholar] [CrossRef]
- Braca, A.; Fico, G.; Morelli, I.; De Simone, F.; Tomé, F.; De Tommasi, N. Antioxidant and free radical scavenging activity of flavonol glycosides from different Aconitum species. J. Ethnopharmacol. 2003, 86, 63–67. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Csepregi, K.; Neugart, S.; Schreine, M.; Hideg, É. Comparative evaluation of total antioxidant capacities of plant polyphenols. Molecules 2016, 21, E208. [Google Scholar] [CrossRef] [Green Version]
- Bidchol, A.M.; Wilfred, A.; Abhijna, P.; Harish, R. Free radical scavenging activity of aqueous and ethanolic extract of Brassica oleracea L. var. italica. Food Bioprocess Tech. 2011, 4, 1137–1143. [Google Scholar] [CrossRef]
- Anwar, F.; Kalsoom, U.; Sultana, B.; Mushtaq, M.; Mehmood, T.; Arshad, H.A. Effect of drying method and extraction solvent on the total phenolics and antioxidant activity of cauliflower (Brassica oleracea L.) extracts. Int. Food Res. J. 2013, 20, 653–659. [Google Scholar]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Augusto, T.R.; Salinas, E.S.S.; Alencar, S.M.; D’arce, M.A.B.R.; de Camargo, A.C.; Vieira, T.M.F.D.S. Phenolic compounds and antioxidant activity of hydroalcoholic extracts of wild and cultivated murtilla (Ugni molinae Turcz.). Food Sci. Technol. 2014, 34, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-H.; Lee, H.-J.; Cho, M.R.; Chang, N.; Kim, Y.; Oh, S.-Y.; Kang, M.-H. Total antioxidant capacity of the Korean diet. Nutr. Res. Pract. 2014, 8, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokozawa, T.; Kim, H.Y.; Cho, E.J.; Choi, J.S.; Chung, H.Y. Antioxidant effects of isorhamnetin 3,7-di-O-beta-d-glucopyranoside isolated from mustard leaf (Brassica juncea) in rats with streptozotocin-induced diabetes. J. Agric. Food. Chem. 2002, 50, 5490–5495. [Google Scholar] [CrossRef]
- Anupama, M.; Murgan, S.S.; Murthy, P.B. Broccoli flower head extract reduces mitomycin-C induced sister chromatid exchange in cultured human lymphocytes. Food Chem. Toxicol. 2008, 46, 3351–3353. [Google Scholar] [CrossRef]
- Lam, T.K.; Gallicchio, L.; Lindsley, K.; Shiels, M.; Hammond, E.; Tao, X.G.; Chen, L.; Robinson, K.A.; Caulfield, L.E.; Herman, J.G.; et al. Cruciferous vegetable consumption and lung cancer risk: A systematic review. Cancer Epidemiol. Biomarkers. Prev. 2009, 18, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Chan, F.K.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. Methods Mol. Biol. 2013, 979, 65–70. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula aetnensis Raf. (Betulaceae) extract induced HO-1 Expression and ferroptosis cell death in human colon cancer cells. Int. J. Mol. Sci. 2019, 20, E2723. [Google Scholar] [CrossRef] [Green Version]
- Tadić, V.M.; Jeremic, I.; Dobric, S.; Isakovic, A.; Markovic, I.; Trajkovic, V.; Bojovic, D.; Arsic, I. Anti-inflammatory, gastroprotective, and cytotoxic effects of Sideritis scardica extracts. Planta Med. 2012, 78, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Sargent, J.M.; Williamson, C.J.; Maliepaard, M.; Elgie, A.W.; Scheper, R.J.; Taylor, C.G. Breast cancer resistance protein expression and resistance to daunorubicin in blast cells from patients with acute myeloid leukaemia. Br. J. Haematol. 2001, 115, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Miret, S.; Abrahamse, L.; de Groene, E.M. Comparison of in vitro models for the prediction of compound absorption across the human intestinal mucosa. J. Biomol. Screen. 2004, 9, 598–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaur, K.; Vázquez-Salgado, A.M.; Duran-Camacho, G.; Dominguez-Martinez, I.; Benjamín-Rivera, J.A.; Fernández-Vega, L.; Carmona Sarabia, L.; Cruz García, A.; Pérez-Deliz, F.; Méndez Román, J.A.; et al. Iron and copper intracellular chelation as an anticancer drug strategy. Inorganics 2018, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Veni, T.; Pushpanathan, T. Comparison of the Artemia salina and Artemia fransiscana bioassays for toxicity of Indian medicinal plants. J. Coast. Life Med. 2014, 2, 453–457. [Google Scholar] [CrossRef]
- Vitali, F.; Pennisi, C.; Tomaino, A.; Bonina, F.; Pasquale, A.; Saija, A.; Tita, B. Effect of a standardized extract of red orange juice on proliferation of human prostate cells in vitro. Fitoterapia 2006, 77, 151–155. [Google Scholar] [CrossRef]
- Hong, L.S.; Ibrahim, D.; Kassim, J. Assessment of in vivo and in vitro cytotoxic activity of hydrolysable tannin extracted from Rhizophora apiculata barks. World J. Microbiol. Biotechnol. 2011, 27, 2737–2740. [Google Scholar] [CrossRef]
- Mohti, H.; Taviano, M.F.; Cacciola, F.; Dugo, P.; Mondello, L.; Zaid, A.; Cavò, E.; Miceli, N. Silene vulgaris subsp. macrocarpa leaves and roots from Morocco: Assessment of the efficiency of different extraction techniques and solvents on their antioxidant capacity, brine shrimp toxicity and phenolic characterization. Plant Biosyst. 2019. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Sinatra, F.; Villaggio, G.; Amenta, F.; Avola, R.; Renis, M. “Reactive” response evaluation of primary human astrocytes after methylmercury exposure. J. Neurosci. Res. 2014, 92, 95–103. [Google Scholar] [CrossRef]
- Acquaviva, R.; Sorrenti, V.; Santangelo, R.; Cardile, V.; Tomasello, B.; Malfa, G.; Vanella, L.; Amodeo, A.; Genovese, C.; Mastrojeni, S.; et al. Effects of an extract of Celtis aetnensis (Tornab.) Strobl twigs on human colon cancer cell cultures. Oncol. Rep. 2016, 36, 2298–2304. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound | tR (min) | [M − H]− | mg/g ± %RSD (A) | mg/g ± %RSD (B) |
|---|---|---|---|---|---|
| a | Kaempferol-3-O-diglucoside-7-O-glucoside | 8.4 | 773 (609) | - | 1.11 ± 1.44 |
| b | Quercetin-3-sophoroside-7-glucoside | 10.7 | 787 (625) | - | 1.32 ± 1.32 |
| 1 | Quercetin-3-hydroxyferuloylsophoroside-7-glucoside | 13.9 | 979 (625) | 1.91 ± 0.52 | 1.59 ± 0.57 |
| 2 | Quercetin-3-caffeoylsophoroside-7-glucoside | 15.2 | 949 (625) | 1.55 ± 1.23 | 1.22 ± 1.41 |
| 3 | Kaempferol-3-hydroxyferuloylsophoroside-7-glucoside | 15.6 | 963 (801) | 1.43 ± 1.21 | 1.41 ± 1.39 |
| 4 | Quercetin-3-sinapoyltriglucoside-7-glucoside | 16.3 | 1155 (831) | 1.22 ± 1.11 | 0.62 ± 1.87 |
| 5 | Quercetin-3-feruloyl-diglucoside-7-glucoside | 17.6 | 963 (801) | 1.52± 1.32 | 1.17 ± 1.11 |
| 6 | Kaempferol-3-sinapoylsophoroside-7-glucoside | 18.5 | 977 (817) | 2.84 ± 1.52 | 0.59 ± 1.98 |
| 7 | Kaempferol-3-feruloylsophoroside-7-glucoside | 19.9 | 947 (609) | 2.11 ± 0.98 | 2.14 ± 0.48 |
| 8 | Isorhamnetin-3-glucoside-7-glucoside | 20.7 | 639 (747) | 3.33 ± 0.77 | 1.79 ± 0.32 |
| c | Feruloylmalate | 21.3 | 309 | - | N.Q. |
| 9 | Sinapoylmalic acid | 25.2 | 339 (223) | N.Q. | N.Q. |
| 10 | Sinapoyl-hydroxyferuloyldiglycoside | 28.6 | 739 (515) | N.Q. | N.Q. |
| 11 | Isorhamnetinglycoside | 29.8 | 477 (315) | 1.28 ± 0.54 | 0.57 ± 2.01 |
| 12 | Kaempferolglycoside | 30.7 | 447 (285) | 0.64 ± 1.08 | 0.42 ± 1.94 |
| 13 | Disinapoylgentiobiose | 33.6 | 753 (529) | N.Q. | N.Q. |
| 14 | Sinapoylferuloylgentiobiose | 34.8 | 723 (529) | N.Q. | N.Q. |
| 15 | Diferuloyldiglucoside | 35.4 | 693 (499) | N.Q. | N.Q. |
| 16 | Trisinapoylgentiobiose | 36.7 | 959 (735, 529) | N.Q. | N.Q. |
| 17 | Feruloyldisinapoylgentiobiose | 37.7 | 929 (705, 511) | N.Q. | N.Q. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miceli, N.; Cavò, E.; Ragusa, M.; Cacciola, F.; Mondello, L.; Dugo, L.; Acquaviva, R.; Malfa, G.A.; Marino, A.; D’Arrigo, M.; et al. Brassica incana Ten. (Brassicaceae): Phenolic Constituents, Antioxidant and Cytotoxic Properties of the Leaf and Flowering Top Extracts. Molecules 2020, 25, 1461. https://doi.org/10.3390/molecules25061461
Miceli N, Cavò E, Ragusa M, Cacciola F, Mondello L, Dugo L, Acquaviva R, Malfa GA, Marino A, D’Arrigo M, et al. Brassica incana Ten. (Brassicaceae): Phenolic Constituents, Antioxidant and Cytotoxic Properties of the Leaf and Flowering Top Extracts. Molecules. 2020; 25(6):1461. https://doi.org/10.3390/molecules25061461
Chicago/Turabian StyleMiceli, Natalizia, Emilia Cavò, Monica Ragusa, Francesco Cacciola, Luigi Mondello, Laura Dugo, Rosaria Acquaviva, Giuseppe Antonio Malfa, Andreana Marino, Manuela D’Arrigo, and et al. 2020. "Brassica incana Ten. (Brassicaceae): Phenolic Constituents, Antioxidant and Cytotoxic Properties of the Leaf and Flowering Top Extracts" Molecules 25, no. 6: 1461. https://doi.org/10.3390/molecules25061461