A2E Distribution in RPE Granules in Human Eyes

 , ,
, ,
Abstract
:1. Introduction
2. Results
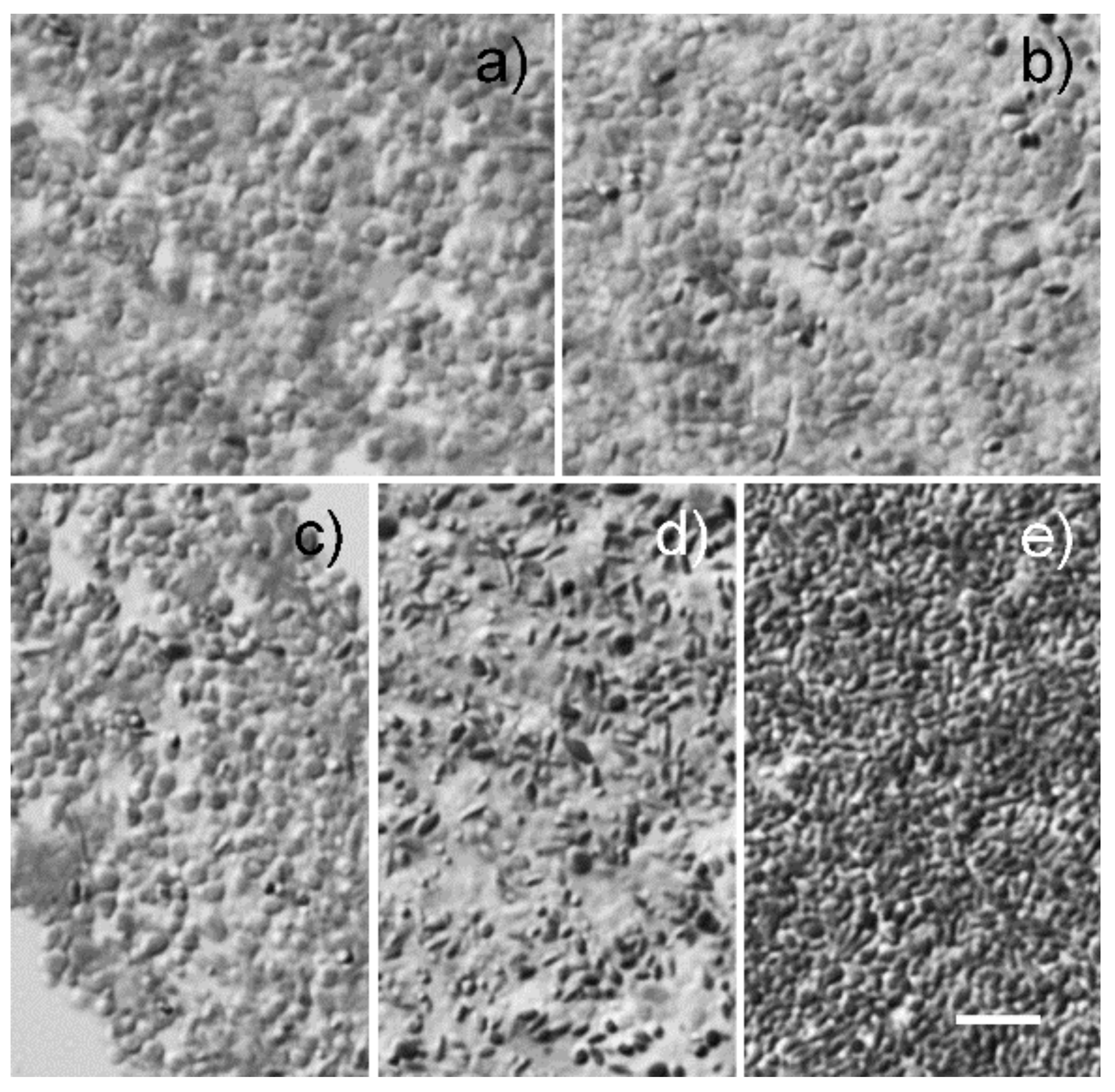
2.1. Morphological Analysis of Granules
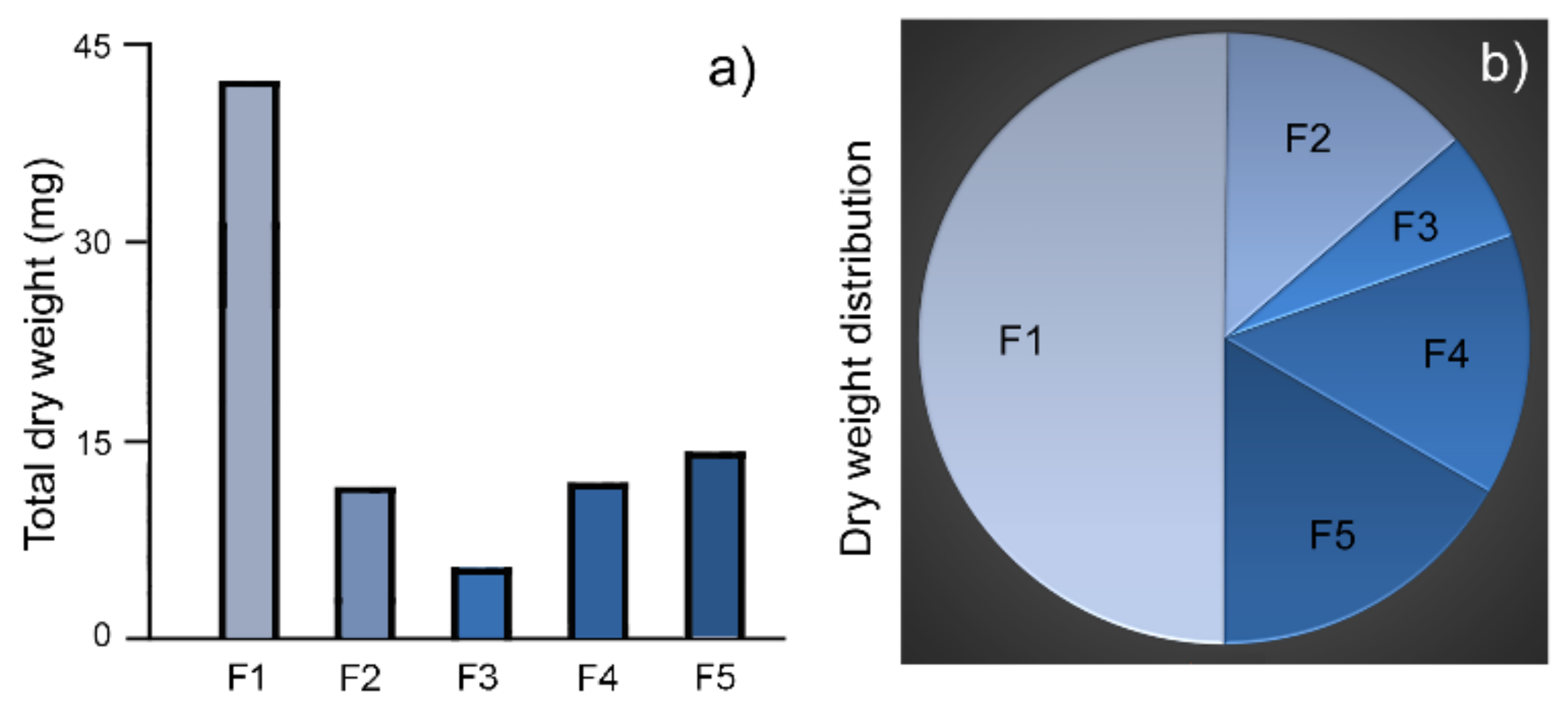
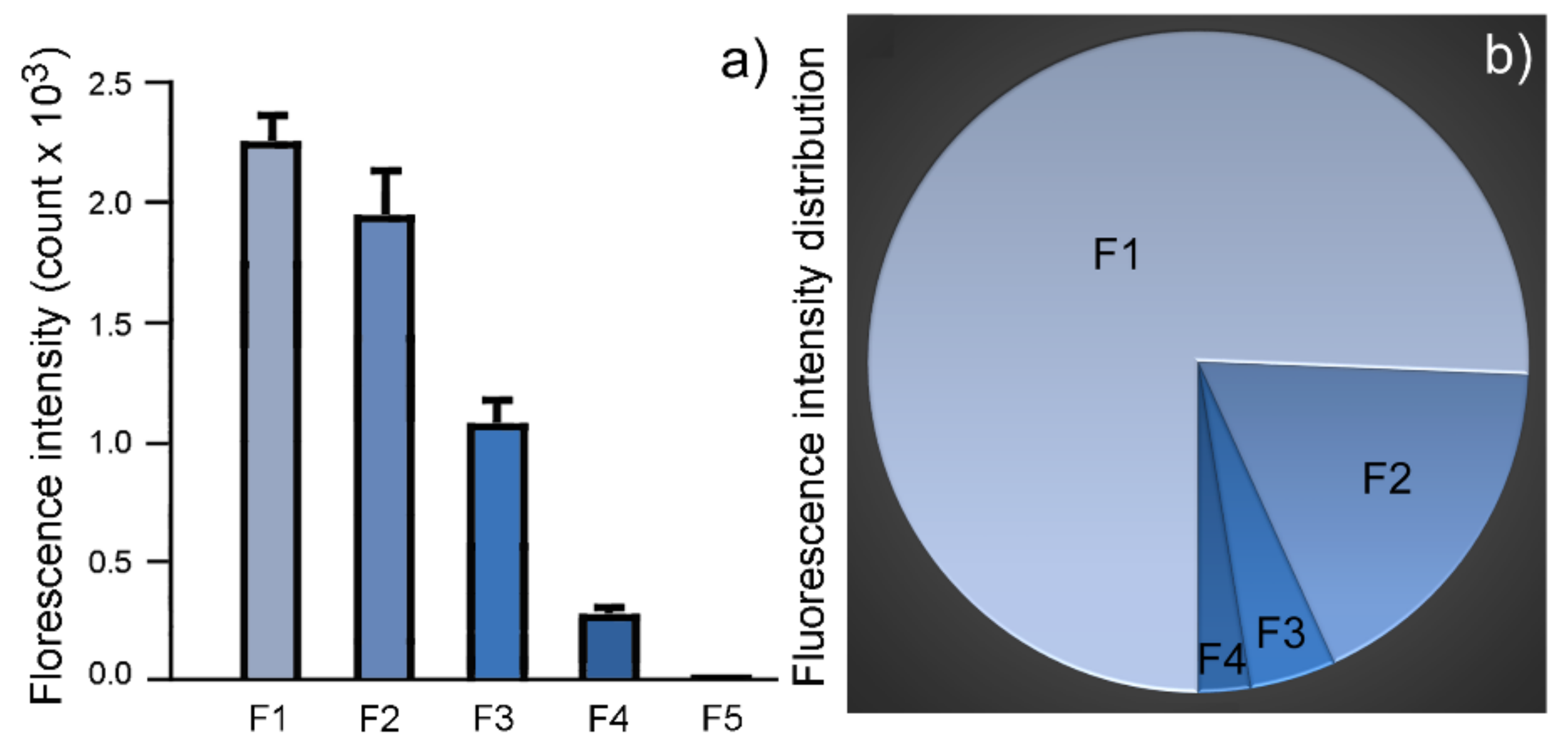
2.2. Weight Distribution in Granule Fractions
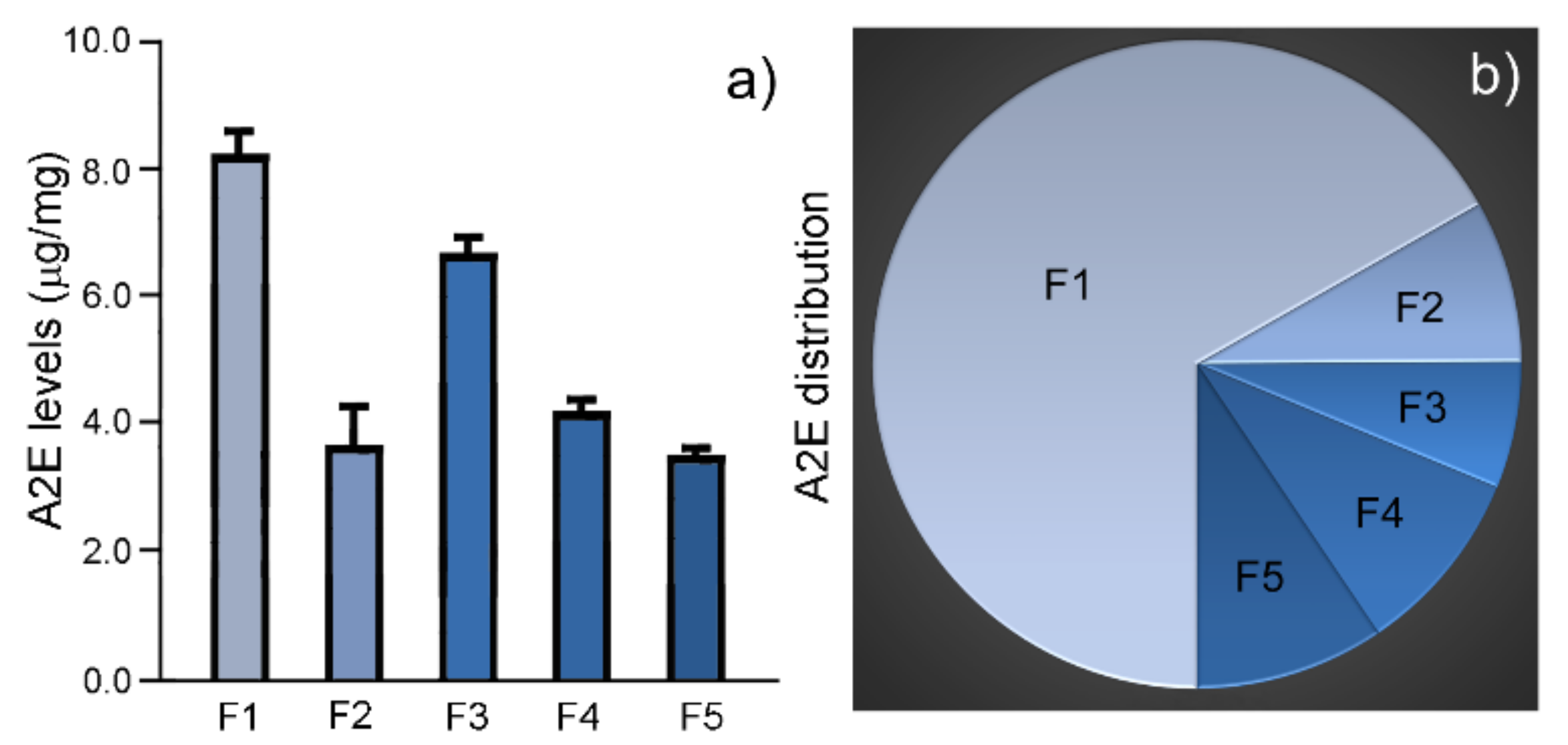
2.3. A2E Levels and Distribution in Granule Fractions
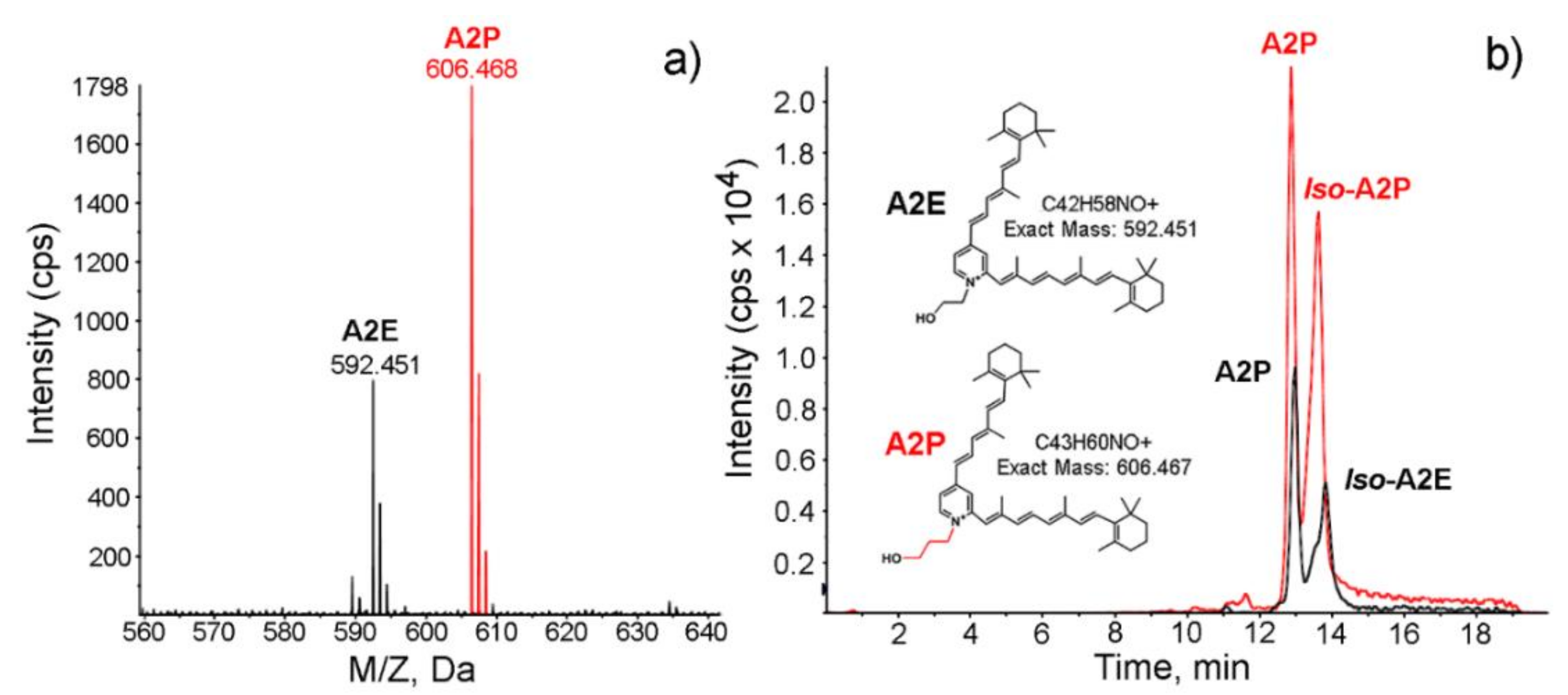
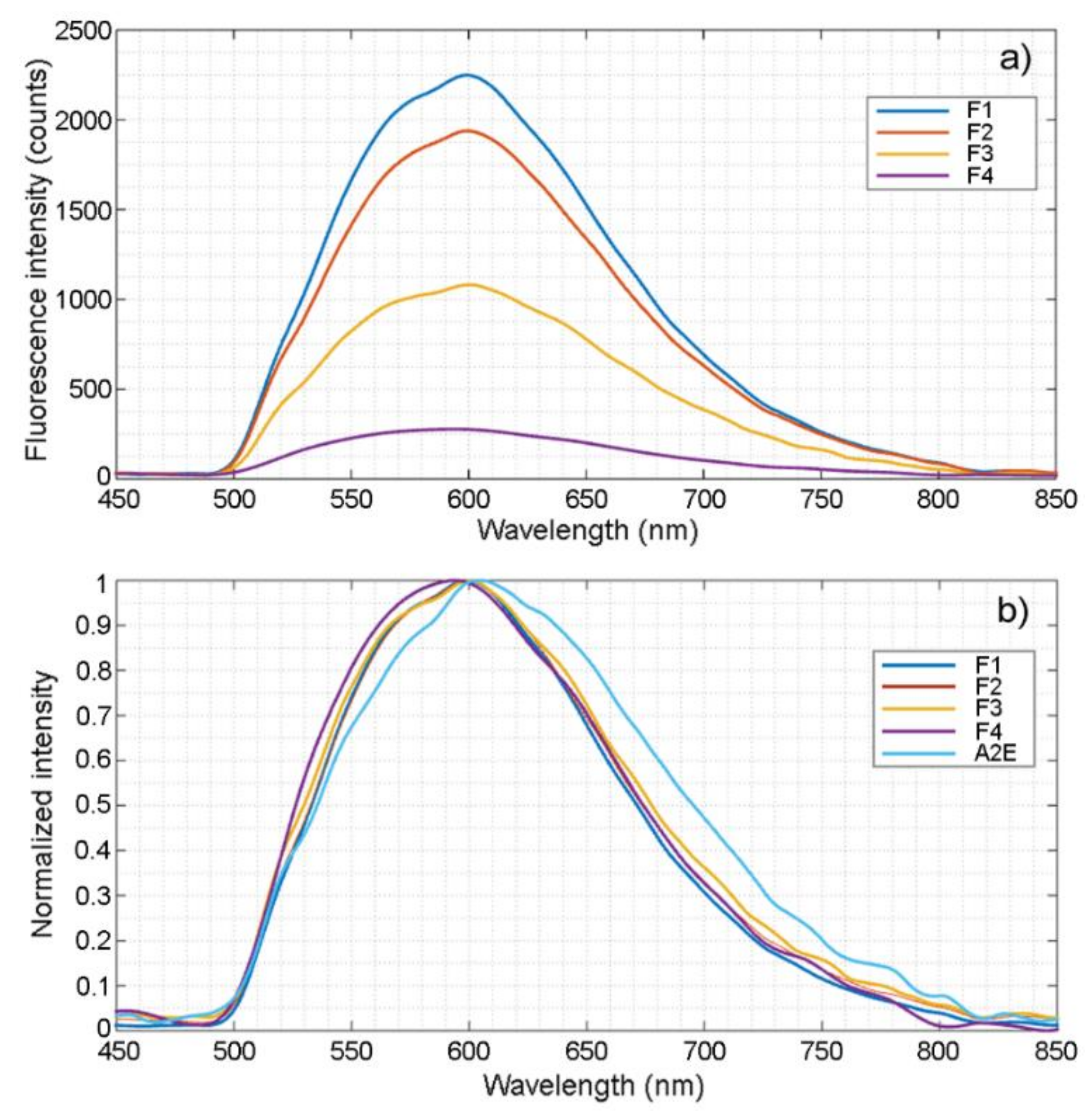
2.4. Fluorescence Spectroscopic Analysis
3. Discussion
4. Materials and Methods
4.1. Isolation of Lipofuscin Granules From Human RPE Cells
4.2. Dry Weight Measurement
4.3. Morphological Analysis
4.4. Synthesis of A2E and A2P
4.5. Quantification of A2E in RPE Granules
4.6. Fluorescence Spectroscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brunk, U.T.; Terman, A. Lipofuscin: Mechanisms of age-related accumulation and influence on cell function. Free Radic. Biol. Med. 2002, 33, 611–619. [Google Scholar] [CrossRef]
- Kaur, J.; Debnath, J. Autophagy at the crossroads of catabolism and anabolism. Nat. Rev. Mol. Cell Biol. 2015, 16, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Sulzer, D.; Mosharov, E.; Talloczy, Z.; Zucca, F.A.; Simon, J.D.; Zecca, L. Neuronal pigmented autophagic vacuoles: Lipofuscin, neuromelanin, and ceroid as macroautophagic responses during aging and disease. J. Neurochem. 2008, 106, 24–36. [Google Scholar] [CrossRef]
- Katz, M.L.; Robison, W.G., Jr. What is lipofuscin? Defining characteristics and differentiation from other autofluorescent lysosomal storage bodies. Arch. Gerontol. Geriatr. 2002, 34, 169–184. [Google Scholar] [CrossRef]
- Moreno-Garcia, A.; Kun, A.; Calero, O.; Medina, M.; Calero, M. An Overview of the Role of Lipofuscin in Age-Related Neurodegeneration. Front. Neurosci. 2018, 12, 464. [Google Scholar] [CrossRef]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [Green Version]
- Strauss, O. The retinal pigment epithelium. In Webvision The Orignaization of the Retina and Visual System; Kolb, H., Nelson, R., Fernandez, E., Jones, B., Eds.; John Moran Eye Center, University of Utah: Salt Lake City, UT, USA, 2012; Available online: https://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the-retina/the-retinal-pigment-epithelium/ (accessed on 18 February 2020).
- Young, R.W. The renewal of photoreceptor cell outer segments. J. Cell Biol. 1967, 33, 61–72. [Google Scholar] [CrossRef]
- Young, R.W.; Bok, D. Participation of the retinal pigment epithelium in the rod outer segment renewal process. J. Cell Biol. 1969, 42, 392–403. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Boulton, M. RPE lipofuscin and its role in retinal pathobiology. Exp. Eye Res. 2005, 80, 595–606. [Google Scholar] [CrossRef]
- Eldred, G.E.; Lasky, M.R. Retinal age pigments generated by self-assembling lysosomotropic detergents. Nature 1993, 361, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Decatur, J.; Nakanishi, K.; Eldred, G.E. Ocular age pigment ‘‘A2-E’’: An unprecedented pyridinium bisretinoid. J. Am. Chem. Soc. 1996, 118, 1559–1560. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Fishkin, N.; Zhou, J.; Cai, B.; Jang, Y.P.; Krane, S.; Itagaki, Y.; Nakanishi, K. A2E, a byproduct of the visual cycle. Vision Res. 2003, 43, 2983–2990. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shabat, S.; Parish, C.A.; Vollmer, H.R.; Itagaki, Y.; Fishkin, N.; Nakanishi, K.; Sparrow, J.R. Biosynthetic studies of A2E, a major fluorophore of retinal pigment epithelial lipofuscin. J. Biol. Chem. 2002, 277, 7183–7190. [Google Scholar] [CrossRef] [Green Version]
- Petrukhin, K. Pharmacological inhibition of lipofuscin accumulation in the retina as a therapeutic strategy for dry AMD treatment. Drug. Discov. Today Ther. Strateg. 2013, 10, e11–e20. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, Y.; Ma, L.; Washington, I. Deuterium enrichment of vitamin A at the C20 position slows the formation of detrimental vitamin A dimers in wild-type rodents. J. Biol. Chem. 2011, 286, 7958–7965. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Kaufman, Y.; Zhang, J.; Washington, I. C20-D3-vitamin A slows lipofuscin accumulation and electrophysiological retinal degeneration in a mouse model of Stargardt disease. J. Biol. Chem. 2011, 286, 7966–7974. [Google Scholar] [CrossRef] [Green Version]
- Charbel Issa, P.; Barnard, A.R.; Herrmann, P.; Washington, I.; MacLaren, R.E. Rescue of the Stargardt phenotype in Abca4 knockout mice through inhibition of vitamin A dimerization. Proc. Natl. Acad. Sci. USA 2015, 112, 8415–8420. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov. Phase 2 Tolerability and Effects of ALK-001 on Stargardt Disease (TEASE). Available online: https://clinicaltrials.gov/ct2/show/NCT02402660 (accessed on 18 February 2020).
- Boulton, M.; Docchio, F.; Dayhaw-Barker, P.; Ramponi, R.; Cubeddu, R. Age-related changes in the morphology, absorption and fluorescence of melanosomes and lipofuscin granules of the retinal pigment epithelium. Vision Res. 1990, 30, 1291–1303. [Google Scholar] [CrossRef]
- Boulton, M.; Marshall, J. Repigmentation of human retinal pigment epithelial cells in vitro. Exp. Eye Res. 1985, 41, 209–218. [Google Scholar] [CrossRef]
- Bazan, H.E.; Bazan, N.G.; Feeney-Burns, L.; Berman, E.R. Lipids in human lipofuscin-enriched subcellular fractions of two age populations. Comparison with rod outer segments and neural retina. Investig. Ophthalmol. Visual Sci. 1990, 31, 1433–1443. [Google Scholar]
- Hong, L.; Garguilo, J.; Anzaldi, L.; Edwards, G.S.; Nemanich, R.J.; Simon, J.D. Age-dependent photoionization thresholds of melanosomes and lipofuscin isolated from human retinal pigment epithelium cells. Photochem. Photobiol. 2006, 82, 1475–1481. [Google Scholar] [CrossRef]
- Schutt, F.; Ueberle, B.; Schnolzer, M.; Holz, F.G.; Kopitz, J. Proteome analysis of lipofuscin in human retinal pigment epithelial cells. FEBS Lett. 2002, 528, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Taubitz, T.; Fang, Y.; Biesemeier, A.; Julien-Schraermeyer, S.; Schraermeyer, U. Age, lipofuscin and melanin oxidation affect fundus near-infrared autofluorescence. EBioMedicine 2019, 48, 592–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parish, C.A.; Hashimoto, M.; Nakanishi, K.; Dillon, J.; Sparrow, J. Isolation and one-step preparation of A2E and iso-A2E, fluorophores from human retinal pigment epithelium. Proc. Natl. Acad. Sci. USA 1998, 95, 14609–14613. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.; Elliott, M.H.; Floor, E.; Truscott, T.G.; Zareba, M.; Sarna, T.; Shamsi, F.A.; Boulton, M.E. Photocytotoxicity of lipofuscin in human retinal pigment epithelial cells. Free Radic. Biol. Med. 2001, 31, 256–265. [Google Scholar] [CrossRef]
- Panda-Jonas, S.; Jonas, J.B.; Jakobczyk-Zmija, M. Retinal pigment epithelial cell count, distribution, and correlations in normal human eyes. Am. J. Ophthalmol. 1996, 121, 181–189. [Google Scholar] [CrossRef]
- Pollreisz, A.; Messinger, J.D.; Sloan, K.R.; Mittermueller, T.J.; Weinhandl, A.S.; Benson, E.K.; Kidd, G.J.; Schmidt-Erfurth, U.; Curcio, C.A. Visualizing melanosomes, lipofuscin, and melanolipofuscin in human retinal pigment epithelium using serial block face scanning electron microscopy. Exp. Eye Res. 2018, 166, 131–139. [Google Scholar] [CrossRef]
- Delori, F.C.; Dorey, C.K.; Staurenghi, G.; Arend, O.; Goger, D.G.; Weiter, J.J. In vivo fluorescence of the ocular fundus exhibits retinal pigment epithelium lipofuscin characteristics. Investig. Ophthalmol. Visual Sci. 1995, 36, 718–729. [Google Scholar]
- Delori, F.; Keilhauer, C.; Sparrow, J.R.; Staurenghi, G.; Holz, F.; Schmitz-Valckenberg, S.; Spaide, R.; Bird, A. Origin of fundus autofluorescence. In Atlas of Fundus Autofluorescence Imaging; Springer: Berlin/Heidelberg, Germany, 2007; pp. 17–29. [Google Scholar]
- Lu, J.; Luo, L.; Huang, D.; Liu, X.; Xia, X.; Wang, Z.; Lam, B.L.; Yi, J.; Wen, R.; Li, Y. Photoreceptor Protection by Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF). eNeuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Penn, J.; Mihai, D.M.; Washington, I. Morphological and physiological retinal degeneration induced by intravenous delivery of vitamin A dimers in rabbits. Dis. Model. Mech. 2015, 8, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, R.; Lam, B.L.; Guan, Z. Aberrant dolichol chain lengths as biomarkers for retinitis pigmentosa caused by impaired dolichol biosynthesis. J. Lipid Res. 2013, 54, 3516–3522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples are available from the authors upon request. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Dry Weight (mg/100µL) (Mean ± SD) | Total Volume (µL) | Total Weight (mg) | Weight Distribution |
|---|---|---|---|---|
| F1 | 5.0 ± 0.0 (n = 3) | 845 | 42.3 | 50.2% |
| F2 | 4.0 ± 0.0 (n = 3) | 285 | 11.4 | 13.6% |
| F3 | 2.0 ± 0.0 (n = 3) | 245 | 4.9 | 5.8% |
| F4 | 2.5 ± 0.0 (n = 3) | 465 | 11.6 | 13.8% |
| F5 | 4.0 ± 0.1 (n = 3) | 350 | 14.0 | 16.6% |
| Total | 2190 | 84.2 | 100% |
| Fraction | A2E Level (µg/mg, Mean ± SD) | Total A2E | A2E Distribution |
|---|---|---|---|
| F1 | 8.3 ± 0.4 (n = 3) | 348.6 | 66.8% |
| F2 | 3.7 ± 0.6 (n = 3) | 42.2 | 8.1% |
| F3 | 6.7 ± 0.3 (n = 3) | 32.9 | 6.3% |
| F4 | 4.2 ± 0.3 (n = 3) | 48.8 | 9.4% |
| F5 | 3.5 ± 0.1 (n = 3) | 49.3 | 9.4% |
| Total | 521.7 | 100% |
| Fraction | Peak Intensity (at 600 nm, 1 mg in H2O) (Mean ± SD) | Projected Intensity | Projected Intensity Distribution |
|---|---|---|---|
| F1 | 2247 ± 126 (n = 5) | 95,048 | 75.3% |
| F2 | 1983 ± 159 (n = 5) | 22,606 | 17.9% |
| F3 | 1081 ± 86 (n = 5) | 5297 | 4.2% |
| F4 | 273 ± 13 (n = 5) | 3202 | 2.5% |
| F5 | 0 ± 0 (n = 5) | 0 | 0% |
| Total | 126,153 | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, Z.; Li, Y.; Jiao, S.; Yeasmin, N.; Rosenfeld, P.J.; Dubovy, S.R.; Lam, B.L.; Wen, R. A2E Distribution in RPE Granules in Human Eyes. Molecules 2020, 25, 1413. https://doi.org/10.3390/molecules25061413
Guan Z, Li Y, Jiao S, Yeasmin N, Rosenfeld PJ, Dubovy SR, Lam BL, Wen R. A2E Distribution in RPE Granules in Human Eyes. Molecules. 2020; 25(6):1413. https://doi.org/10.3390/molecules25061413
Chicago/Turabian StyleGuan, Ziqiang, Yiwen Li, Shuliang Jiao, Nusrat Yeasmin, Philip J. Rosenfeld, Sander R. Dubovy, Byron L. Lam, and Rong Wen. 2020. "A2E Distribution in RPE Granules in Human Eyes" Molecules 25, no. 6: 1413. https://doi.org/10.3390/molecules25061413