
Synthesis and Matrix Properties of α-Cyano-5-phenyl-2,4-pentadienic Acid (CPPA) for Intact Proteins Analysis by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry



Abstract
:1. Introduction
2. Results and Discussion
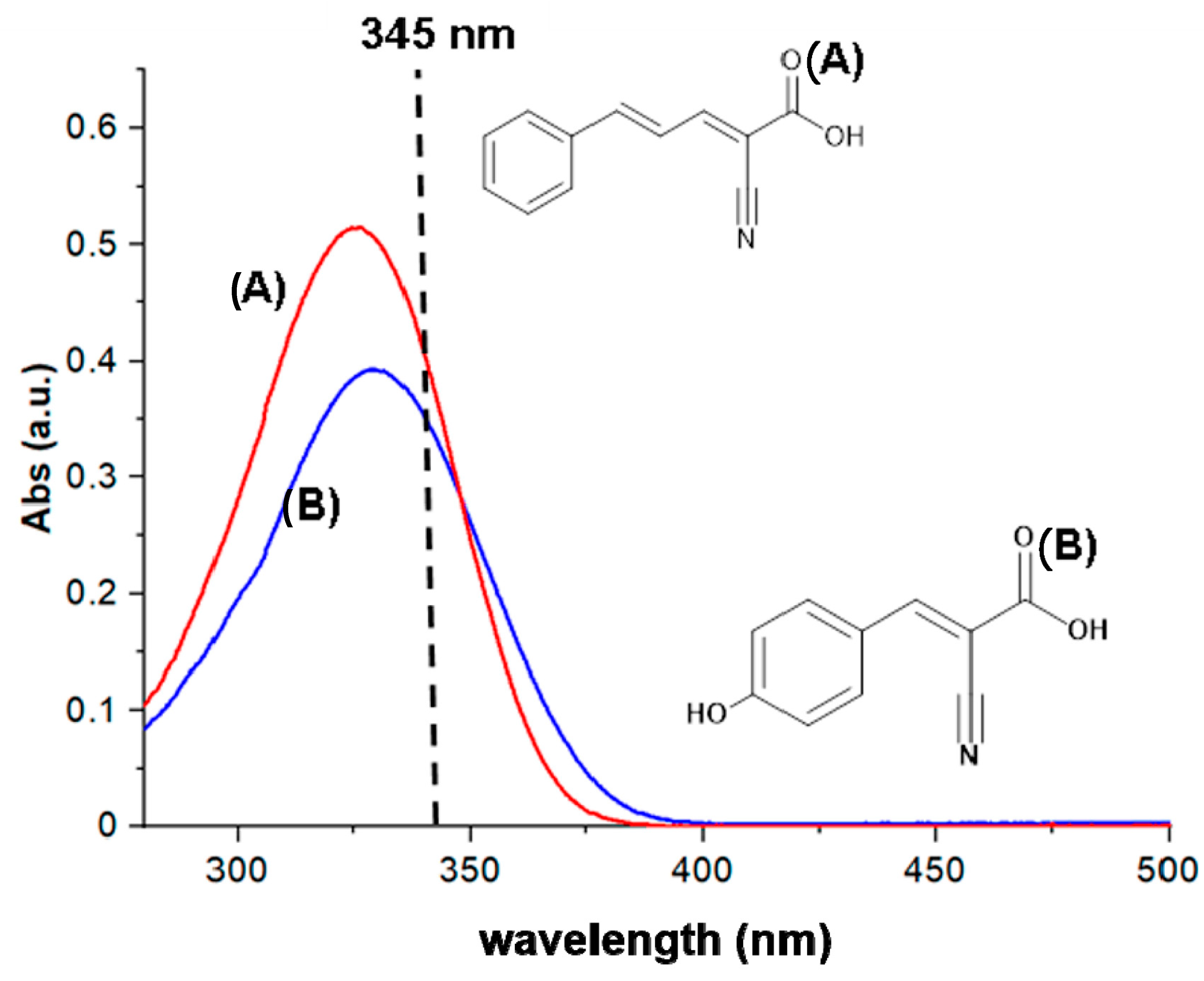
2.1. Synthesis and Characterisation of CPPA
2.2. CPPA as a MALDI Matrix of Standard Proteins
2.3. CPPA as a Matrix for Complex Samples by MALDI-TOF MS
2.4. CPPA as a Matrix for Intact Bacterial Cells in MALDI-TOF MS
3. Materials and Methods
3.1. Materials
3.2. Chemical Synthesis of CPPA
3.3. Matrix Characterization
3.4. Sample Preparations
3.4.1. Standard Proteins
3.4.2. Milk Proteins
3.4.3. Hazelnut Proteins
3.4.4. Bacterial Proteins
3.5. Instrumentations
3.6. Detailed Data Description and Computation Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaufmann, R. Matrix-assisted laser desorption ionization (MALDI) mass spectrometry: A novel analytical tool in molecular biology and biotechnology. J. Biotechnol. 1995, 41, 155–175. [Google Scholar] [CrossRef]
- Webster, J.; Oxley, D. Protein identification by MALDI-TOF mass spectrometry. Methods Mol. Biol. 2012, 800, 227–240. [Google Scholar]
- Calvano, C.D.; Aresta, A.; Iacovone, M.; De Benedetto, G.E.; Zambonin, C.G.; Battaglia, M.; Ditonno, P.; Rutigliano, M.; Bettocchi, C. Optimization of analytical and pre-analytical conditions for MALDI-TOF-MS human urine protein profiles. J. Pharm. Biomed. Anal. 2010, 51, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Hajduk, J.; Matysiak, J.; Kokot, Z.J. Challenges in biomarker discovery with MALDI-TOF MS. Clin. Chim. Acta 2016, 458, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; De Ceglie, C.; Monopoli, A.; Zambonin, C.G. Detection of sheep and goat milk adulterations by direct MALDI-TOF MS analysis of milk tryptic digests. J. Mass Spectrom. 2012, 47, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, R.A.; d’Esposito, D.; Mazzeo, M.F. Food authentication by MALDI MS: MALDI-TOF MS analysis of fish species. In Advances in MALDI and Laser-Induced Soft Ionization Mass Spectrometry; Springer International Publishing: Cham, Switzerland, 2015; pp. 263–277. ISBN 9783319048192. [Google Scholar]
- Camarasa, C.G.; Cobo, F. Application of MALDI-TOF Mass Spectrometry in Clinical Virology. In The Use of Mass Spectrometry Technology (MALDI-TOF) in Clinical Microbiology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 167–180. ISBN 9780128144527. [Google Scholar]
- Mortelmaier, C.; Panda, S.; Robertson, I.; Krell, M.; Christodoulou, M.; Reichardt, N.; Murder, I. Identification performance of MALDI-ToF-MS upon mono- and bi-microbial cultures is cell number and culture proportion dependent. Anal. Bioanal. Chem. 2019, 411, 7027–7038. [Google Scholar] [CrossRef] [Green Version]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [Green Version]
- Aresta, A.; Calvano, C.D.; Palmisano, F.; Zambonin, C.G.; Monaco, A.; Tommasi, S.; Pilato, B.; Paradiso, A. Impact of sample preparation in peptide/protein profiling in human serum by MALDI-TOF mass spectrometry. J. Pharm. Biomed. Anal. 2008, 46, 157–164. [Google Scholar] [CrossRef]
- Mehta, A.; Silva, L.P. MALDI-TOF MS profiling approach: How much can we get from it? Front. Plant Sci. 2015, 6, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Böhme, K.; Calo-Mata, P.; Barros-Velázquez, J.; Ortea, I. Review of Recent DNA-Based Methods for Main Food-Authentication Topics. J. Agric. Food Chem. 2019, 67, 3854–3864. [Google Scholar] [CrossRef]
- Kohl, C.; Brinkmann, A.; Dabrowski, P.W.; Radonić, A.; Nitsche, A.; Kurth, A. Protocol for metagenomic virus detection in clinical specimens. Emerg. Infect. Dis. 2015, 21, 48–57. [Google Scholar] [CrossRef]
- Tang, W.; Ranganathan, N.; Shahrezaei, V.; Larrouy-Maumus, G. MALDI-TOF mass spectrometry on intact bacteria combined with a refined analysis framework allows accurate classification of MSSA and MRSA. PLoS ONE 2019, 14, e0218951. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Arcangeletti, M.C.; Rodighiero, I.; Buttrini, M.; Gorrini, C.; Motta, F.; Germini, D.; Medici, M.C.; Chezzi, C.; De Conto, F. Matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry applied to virus identification. Sci. Rep. 2014, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez, I.; Frenich, A.G.; Romero-González, R. Mass spectrometry approaches to ensure food safety. Anal. Methods 2020, 12, 1148–1162. [Google Scholar] [CrossRef]
- Shao, M.; Bi, H. Direct identification of fish species by surface molecular transferring. Analyst 2020, 145, 4148–4155. [Google Scholar] [CrossRef] [PubMed]
- De Ceglie, C.; Calvano, C.D.; Zambonin, C.G. Determination of Hidden Hazelnut Oil Proteins in Extra Virgin Olive Oil by Cold Acetone Precipitation Followed by In-Solution Tryptic Digestion and MALDI-TOF-MS Analysis. J. Agric. Food Chem. 2014, 62, 9401–9409. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Ceglie, C.D.; D’Accolti, L.; Zambonin, C. MALDI-TOF mass spectrometry detection of extra-virgin olive oil adulteration with hazelnut oil by analysis of phospholipids using an ionic liquid as matrix and extraction solvent. Food Chem. 2012, 134, 1192–1198. [Google Scholar] [CrossRef]
- Liébana-Martos, C. Indications, Interpretation of Results, Advantages, Disadvantages, and Limitations of MALDI-TOF. In The Use of Mass Spectrometry Technology (MALDI-TOF) in Clinical Microbiology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 75–86. ISBN 9780128144527. [Google Scholar]
- Tholey, A.; Heinzle, E. Ionic (liquid) matrices for matrix-assisted laser desorption/ionization mass spectrometry-applications and perspectives. Anal. Bioanal. Chem. 2006, 386, 24–37. [Google Scholar] [CrossRef]
- Monopoli, A.; Cotugno, P.; Cortese, M.; Calvano, C.D.; Ciminale, F.; Nacci, A. Selective N-alkylation of arylamines with alkyl chloride in ionic liquids: Scope and applications. Eur. J. Org. Chem. 2012, 16, 3105–3111. [Google Scholar] [CrossRef]
- Calvano, C.D.; Monopoli, A.; Cataldi, T.R.I.; Palmisano, F. MALDI matrices for low molecular weight compounds: An endless story? Anal. Bioanal. Chem. 2018, 410, 4015–4038. [Google Scholar] [CrossRef]
- Monopoli, A.; Calvano, C.D.; Nacci, A.; Palmisano, F. Boronic acid chemistry in MALDI MS: A step forward in designing a reactive matrix with molecular recognition capabilities. Chem. Commun. 2014, 50, 4322–4324. [Google Scholar] [CrossRef]
- Osaka, I.; Sakai, M.; Takayama, M. 5-Amino-1-naphthol, a novel 1,5-naphthalene derivative matrix suitable for matrixassisted laser desorption/ionization insource decay of phosphorylated peptides. Rapid Commun. Mass Spectrom. 2013, 27, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, L.; Wang, J.; Hou, J.; He, Q.; Liu, J.; Wang, J.; Xiong, S.; Yang, G.; Nie, Z. 2,3,4,5-Tetrakis(3′,4′-dihydroxylphenyl)thiophene: A new matrix for the selective analysis of low molecular weight amines and direct determination of creatinine in urine by MALDI-TOF MS. Anal. Chem. 2012, 84, 10291–10297. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Cataldi, T.R.I.; Kögel, J.F.; Monopoli, A.; Palmisano, F.; Sundermeyer, J. Structural Characterization of Neutral Saccharides by Negative Ion MALDI Mass Spectrometry Using a Superbasic Proton Sponge as Deprotonating Matrix. J. Am. Soc. Mass Spectrom. 2017, 28, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Capozzi, M.A.M.; Punzi, A.; Farinola, G.M.; Cataldi, T.R.I.; Palmisano, F. 1,5-Diaminonaphtalene is a Highly Performing Electron-Transfer Secondary-Reaction Matrix for Laser Desorption Ionization Mass Spectrometry of Indolenine-Based Croconaines. ACS Omega 2018, 3, 17821–17827. [Google Scholar] [CrossRef]
- Shroff, R.; Svatoš, A. Proton sponge: A novel and versatile MALDI matrix for the analysis of metabolites using mass spectrometry. Anal. Chem. 2009, 81, 7954–7959. [Google Scholar] [CrossRef]
- Calvano, C.D.; Cataldi, T.R.I.; Kögel, J.F.; Monopoli, A.; Palmisano, F.; Sundermeyer, J.; Kögel, J.F. Superbasic alkyl-substituted bisphosphazene proton sponges: A new class of deprotonating matrices for negative ion matrix-assisted ionization/laser desorption mass spectrometry of low molecular weight hardly ionizable analytes. Rapid Commun. Mass Spectrom. 2017, 30, 1680–1686. [Google Scholar] [CrossRef]
- Williams, T.L.; Andrzejewski, D.; Lay, J.O.; Musser, S.M. Experimental factors affecting the quality and reproducibility of MALDI TOF mass spectra obtained from whole bacteria cells. J. Am. Soc. Mass Spectrom. 2003, 14, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Jaskolla, T.W.; Lehmann, W.D.; Karas, M. 4-Chloro-α-cyanocinnamic acid is an advanced, rationally designed MALDI matrix. Proc. Natl. Acad. Sci. USA 2008, 105, 12200–12205. [Google Scholar] [CrossRef] [Green Version]
- Jones, G. The Knoevenagel Condensation. In Organic Reactions; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 204–599. [Google Scholar]
- Singh, N.; Sandhu, J.S. Studies in Conjugated Imines: Addition of Active Methylene Compounds. J. Indian Chem. Soc. 1969, 46, 751–753. [Google Scholar]
- Bahr, U.; Jaskolla, T.W. Employing ‘Second Generation’ Matrices. In Advances in MALDI and Laser-Induced Soft Ionization Mass Spectrometry; Cramer, R., Ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Liu, X.; Cole, J.M.; Low, K.S. Solvent Effects on the UV–vis Absorption and Emission of Optoelectronic Coumarins: A Comparison of Three Empirical Solvatochromic Models. J. Phys. Chem. C 2013, 117, 14731–14741. [Google Scholar] [CrossRef]
- Williams, J.B.; Gusev, A.I.; Hercules, D.M. Use of liquid matrices for matrix-assisted laser desorption ionization of polyglycols and poly(dimethylsiloxanes). Macromolecules 1996, 29, 8144–8150. [Google Scholar] [CrossRef]
- Knochenmuss, R.; Dubois, F.; Dale, M.J.; Zenobi, R. The Matrix Suppression Effect and Ionization Mechanisms in Matrix-assisted Laser Desorptiod Ionization. Rapid Commun. Mass Spectrom. 1996, 10, 871–877. [Google Scholar] [CrossRef]
- Guo, Z.; He, L. A binary matrix for background suppression in MALDI-MS of small molecules. Anal. Bioanal. Chem. 2007, 387, 1939–1944. [Google Scholar] [CrossRef]
- Soltwisch, J.; Jaskolla, T.W.; Hillenkamp, F.; Karas, M.; Dreisewerd, K. Ion Yields in UV-MALDI Mass Spectrometry as a Function of Excitation Laser Wavelength and Optical and Physico-Chemical Properties of Classical and Halogen-Substituted MALDI Matrixes. Anal. Chem. 2012, 84, 6567–6576. [Google Scholar] [CrossRef]
- Smirnov, I.P.; Zhu, X.; Taylor, T.; Huang, Y.; Ross, P.; Papayanopoulos, I.A.; Martin, S.A.; Pappin, D.J. Suppression of α-Cyano-4-hydroxycinnamic Acid Matrix Clusters and Reduction of Chemical Noise in MALDI-TOF Mass Spectrometry. Anal. Chem. 2004, 76, 2958–2965. [Google Scholar] [CrossRef]
- Keller, B.O.; Li, L. Discerning matrix-cluster peaks in matrix-assisted laser desorption/ionization time-of-flight mass spectra of dilute peptide mixtures. J. Am. Soc. Mass Spectrom. 2000, 11, 88–93. [Google Scholar] [CrossRef]
- Mirabelli, M.F.; Zenobi, R. Observing Proton Transfer Reactions Inside the MALDI Plume: Experimental and Theoretical Insight into MALDI Gas-Phase Reactions. J. Am. Soc. Mass Spectrom. 2017, 28, 1676–1686. [Google Scholar] [CrossRef]
- Knochenmuss, R. Ion formation mechanisms in UV-MALDI. Analyst 2006, 131, 966–986. [Google Scholar] [CrossRef]
- Molin, L.; Seraglia, R.; Czarnocki, Z.; Maurin, J.K.; Plucinski, F.A.; Traldi, P. On the Primary Ionization Mechanism(s) in Matrix-Assisted Laser Desorption Ionization. J. Anal. Methods Chem. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Karas, M.; Bachmann, D.; Hillekamp, F. Influence of the wavelength in high-irradiance ultraviolet laser desorption mass spectrometry of organic molecules. Anal Chem. 1985, 57, 2935–2939. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H. Application of proteomics to the areas of milk production, processing and quality control—A review. Int. J. Dairy Technol. 2014, 67, 153–166. [Google Scholar] [CrossRef]
- Vincent, D.; Elkins, A.; Condina, M.R.; Ezernieks, V.; Rochfort, S. Quantitation and Identification of Intact Major Milk Proteins for High-Throughput LC-ESI-Q-TOF MS Analyses. PLoS ONE 2016, 11, e0163471. [Google Scholar] [CrossRef] [PubMed]
- Fuquay, J.W. Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1–4170. ISBN 978-0-12-374407-4. [Google Scholar]
- Olumee-Shabon, Z.; Boehmer, J.L. Proteomic Analysis of Goat Milk. In Goat Science; InTech: Rijeka, Croatia, 2018. [Google Scholar] [CrossRef] [Green Version]
- Flinterman, A.E.; Akkerdaas, J.H.; Knlsta, A.C.; Van Ree, R.; Pasmans, S.G. Hazelnut allergy: From pollen-associated mild allergy to severe anaphylactic reactions. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 261–265. [Google Scholar] [CrossRef]
- Lauer, I.; Foetisch, K.; Kolarich, D.; Ballmer-Weber, B.K.; Conti, A.; Altmann, F.; Vieths, S.; Scheurer, S. Hazelnut (Corylus avellana) vicilin Cor a 11: Molecular characterization of a glycoprotein and its allergenic activity. Biochem. J. 2004, 383, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Dooper, M.M.B.W.; Plassen, C.; Holden, L.; Moen, L.H.; Namork, E.; Egaas, E. Antibody binding to hazelnut (Corylus avellana) proteins: The effects of extraction procedure and hazelnut source. Food Agric. Immunol. 2008, 19, 229–240. [Google Scholar] [CrossRef]
- Ribeiro, M.; Costa, J.; Mafra, I.; Cabo, S.; Silva, A.P.; Gonçalves, B.; Hillion, M.; Hébraud, M.; Igrejas, G. Natural Variation of Hazelnut Allergenicity: Is There Any Potential for Selecting Hypoallergenic Varieties? Nutrients 2020, 12, 2100. [Google Scholar] [CrossRef]
- Costa, J.; Mafra, I.; Carrapatoso, I.; Oliveira, M.B.P.P. Hazelnut allergens: Molecular characterization, detection, and clinical relevance. Crit. Rev. Food Sci. Nutr. 2016, 56, 2579–2605. [Google Scholar] [CrossRef] [Green Version]
- Fenselau, C.; Demirev, F.A. Characterization of intact microorganisms by MALDI mass spectrometry. Mass Spectrom. Rev. 2001, 20, 157–171. [Google Scholar] [CrossRef]
- Ryzhov, V.; Fenselau, C. Characterization of the protein subset desorbed by MALDI from whole bacterial cells. Anal. Chem. 2001, 73, 746–750. [Google Scholar] [CrossRef]
- Momo, R.A.; Povey, J.F.; Smales, C.M.; O’Malley, C.J.; Montague, G.A.; Martin, E.B. MALDI-ToF mass spectrometry coupled with multivariate pattern recognition analysis for the rapid biomarker profiling of Escherichia coli in different growth phases. Anal. Bioanal. Chem. 2013, 405, 8251–8265. [Google Scholar] [CrossRef] [PubMed]
- Krause, E.; Wenschuh, H.; Jungblut, P.R. The dominance of arginine-containing peptides in MALDI-derived tryptic mass fingerprints of proteins. Anal. Chem. 1999, 71, 4160–4165. [Google Scholar] [CrossRef] [PubMed]
- Suh, M.J.; Limbach, P.A. Investigation of methods suitable for the matrix-assisted laser desorption/ionization mass spectrometric analysis of proteins from ribonucleoprotein complexes. Eur. J. Mass Spectrom. 2004, 10, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Lavollay, M.; Rostane, H.; Compain, F.; Carbonnelle, E. Identification Methods: Identification of Clinical Microorganisms with MALDI-TOF-MS in a Microbiology Laboratory. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; ISBN 9780123847331. [Google Scholar]
- Vargha, M.; Takáts, Z.; Konopka, A.; Nakatsu, C.H. Optimization of MALDI-TOF MS for strain level differentiation of Arthrobacter isolates. J. Microbiol. Methods 2006, 66, 399–409. [Google Scholar] [CrossRef]
- Božik, M.; Cejnar, P.; Šašková, M.; Nový, P.; Maršík, P.; Klouček, P. Stress response of Escherichia coli to essential oil components-insights on low-molecular-weight proteins from MALDI-TOF. Sci. Rep. 2018, 8, 13042–13050. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Picca, R.A.; Bonerba, E.; Tantillo, G.; Cioffi, N.; Palmisano, F. MALDI-TOF mass spectrometry analysis of proteins and lipids in Escherichia coli exposed to copper ions and nanoparticles. J. Mass Spectrom. 2016, 51, 828–840. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Literature m/z Value | Experimental m/z Value | Identified Protein |
|---|---|---|
| 4309.3 | 4310 | 50S ribosomal protein L36 |
| 4580.04 | 4580 | Osmotically inducible lipoprotein B |
| - | 4775 ([M + 2H]2+) | 30S ribosomal protein S16 |
| 5095.33 | 5090 | Protein S22 |
| 5380.55 | 5382 | 50S ribosomal protein L34 |
| 6314.92 | 6320 | 50S ribosomal protein L32 |
| 6855.88 | 6850 | Carbon storage regulator |
| 7332.3 | 7330 | Cold shock-like protein CspE |
| 7872.22 | 7880 | 50S ribosomal protein L31 |
| - | 8590 | 50S ribosomal protein L2 |
| 9190.21 | 9190 | 30S ribosomal protein S16 |
| 9553.44 | 9550 | 30S ribosomal protein S20 |
| 10,298.28 | 10,300 | 50S ribosomal protein L25 |
| 11,199.26 | 11,200 | 50S ribosomal protein L23 |
| 1227.40 | 12,225 | 50S ribosomal protein L22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monopoli, A.; Nacci, A.; Cataldi, T.R.I.; Calvano, C.D. Synthesis and Matrix Properties of α-Cyano-5-phenyl-2,4-pentadienic Acid (CPPA) for Intact Proteins Analysis by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Molecules 2020, 25, 6054. https://doi.org/10.3390/molecules25246054
Monopoli A, Nacci A, Cataldi TRI, Calvano CD. Synthesis and Matrix Properties of α-Cyano-5-phenyl-2,4-pentadienic Acid (CPPA) for Intact Proteins Analysis by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Molecules. 2020; 25(24):6054. https://doi.org/10.3390/molecules25246054
Chicago/Turabian StyleMonopoli, Antonio, Angelo Nacci, Tommaso R. I. Cataldi, and Cosima D. Calvano. 2020. "Synthesis and Matrix Properties of α-Cyano-5-phenyl-2,4-pentadienic Acid (CPPA) for Intact Proteins Analysis by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry" Molecules 25, no. 24: 6054. https://doi.org/10.3390/molecules25246054