
Neuroprotective Activities of Boophone haemanthoides (Amaryllidaceae) Extract and Its Chemical Constituents



Abstract
:1. Introduction
2. Results
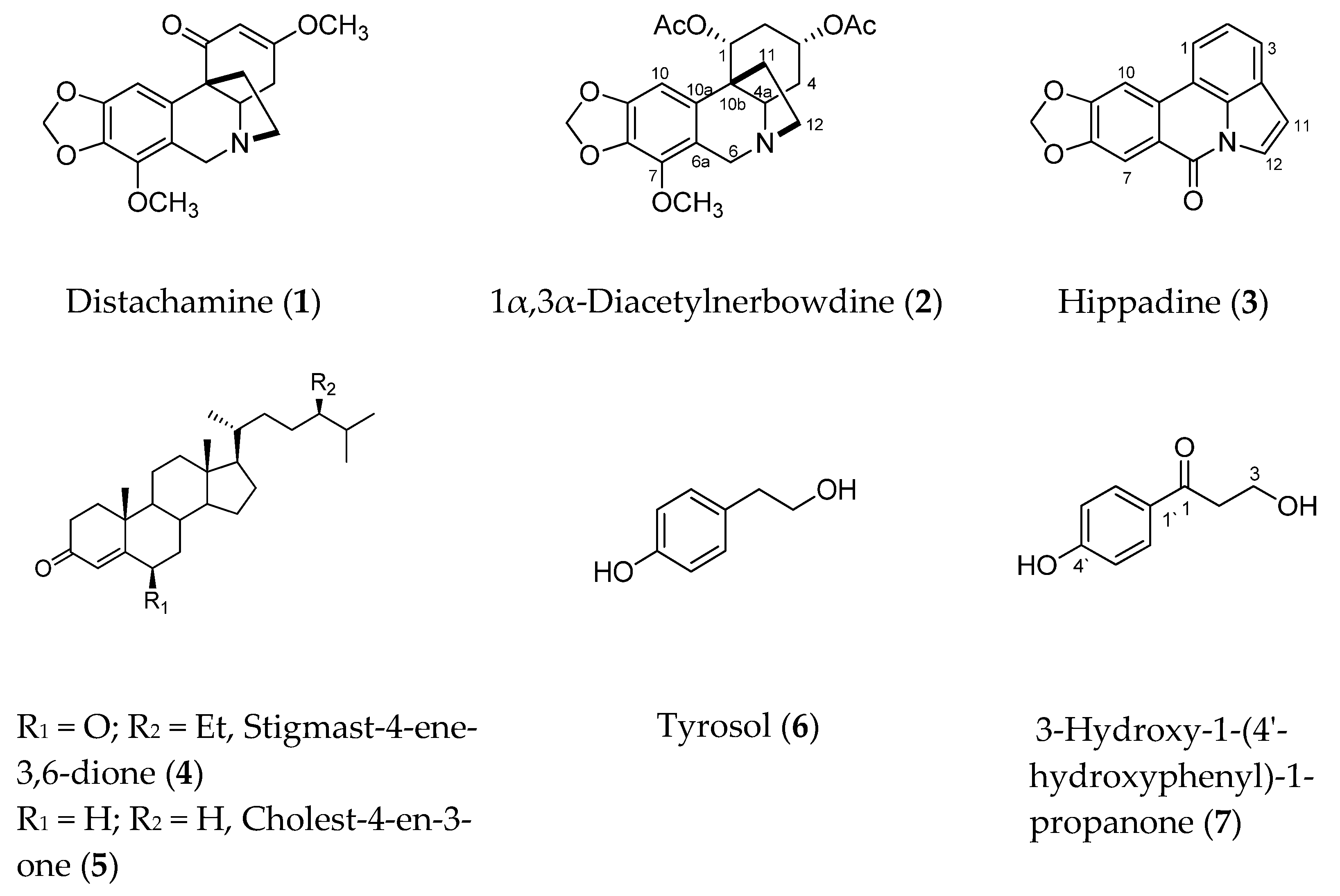
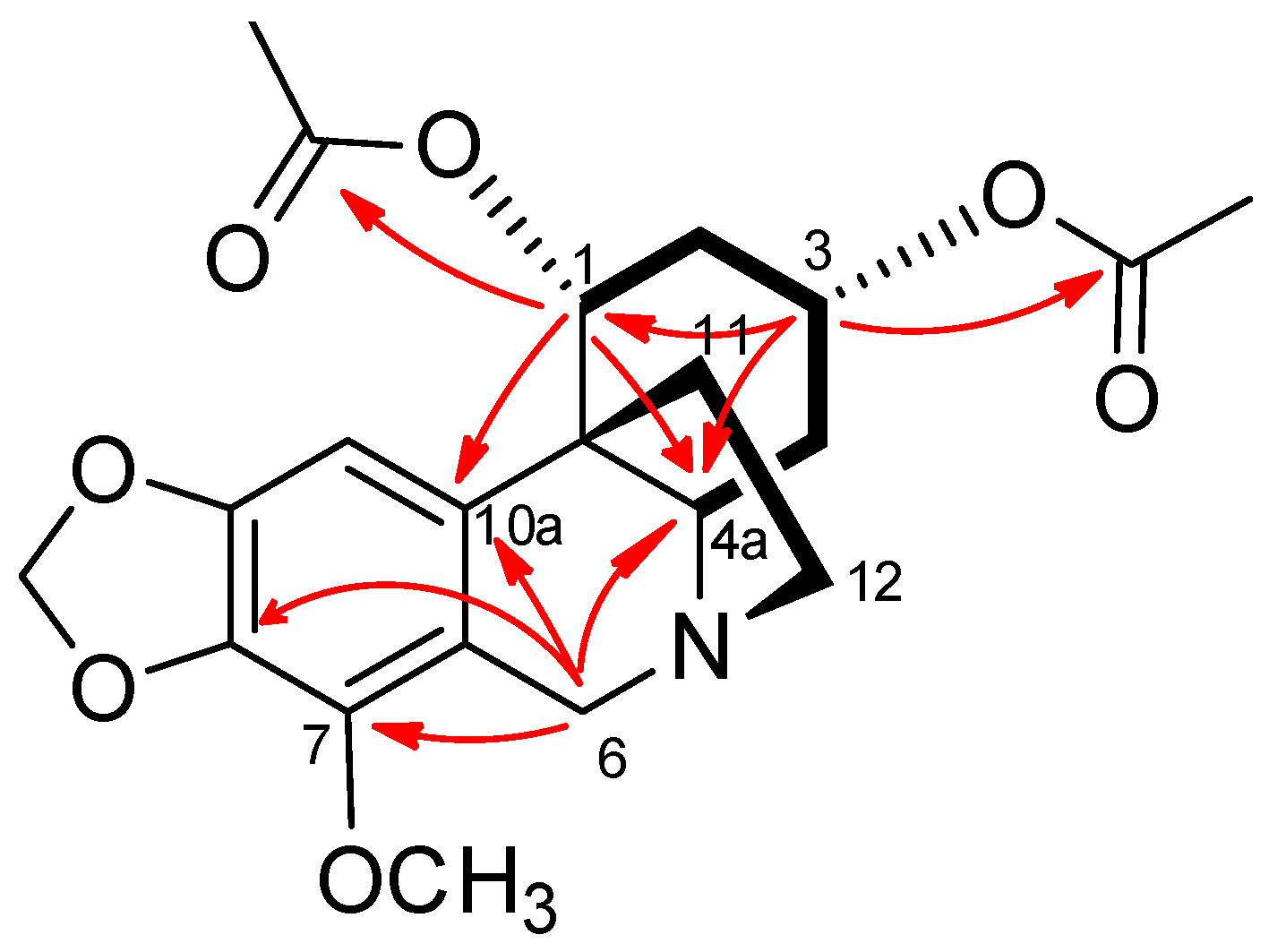
2.1. Isolation and Identification of the Chemical Constituents
2.2. Dose Response of BHE and Isolated Compounds
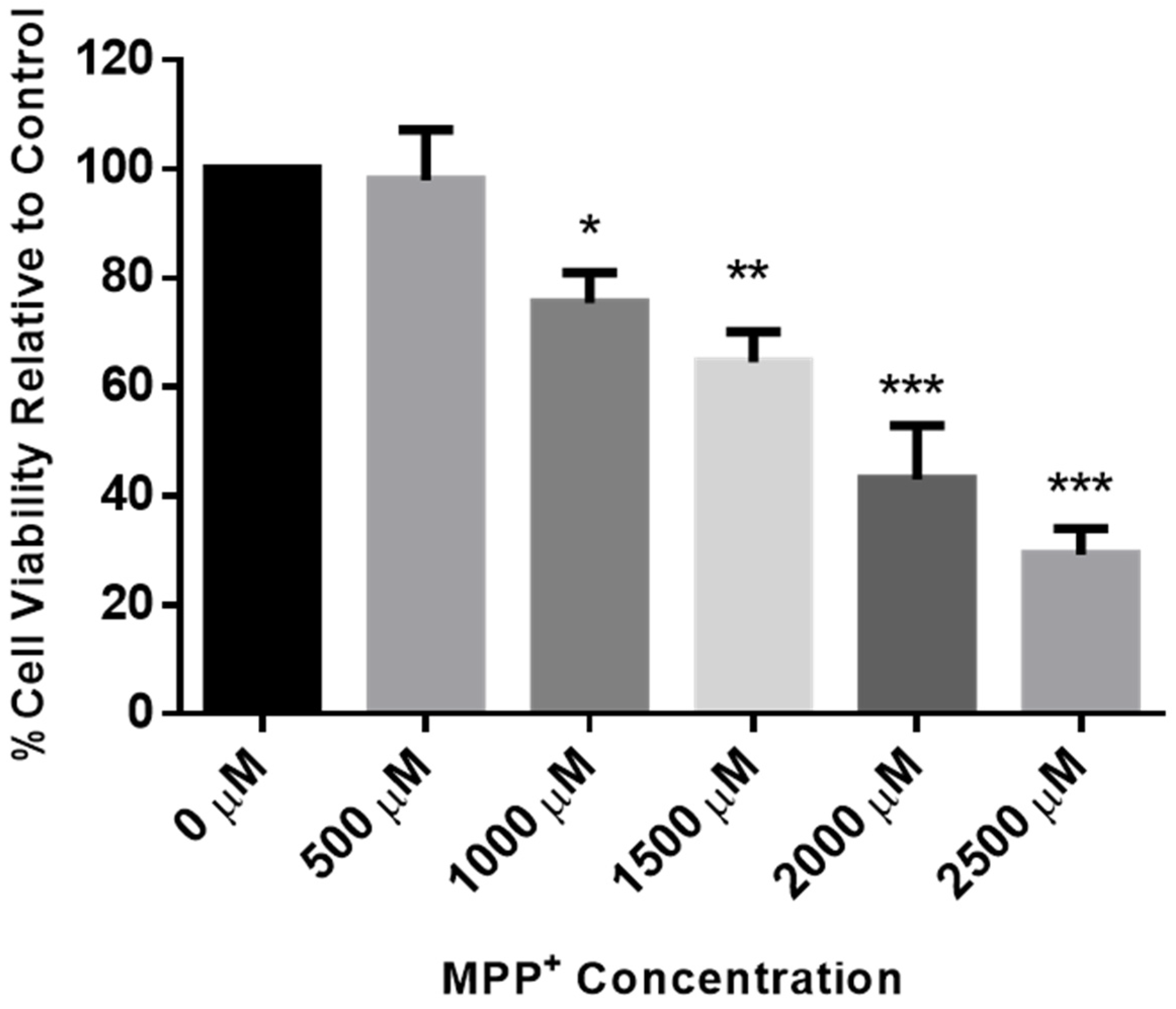
2.3. Dose Response of MPP+ in SH-SY5Y
2.4. BHE and Isolated Compounds Protect SH-SY5Y Cells from MPP+-Induced Toxicity
2.5. BHE Improves Cell Morphology in SH-SY5Y after MPP+ Insult
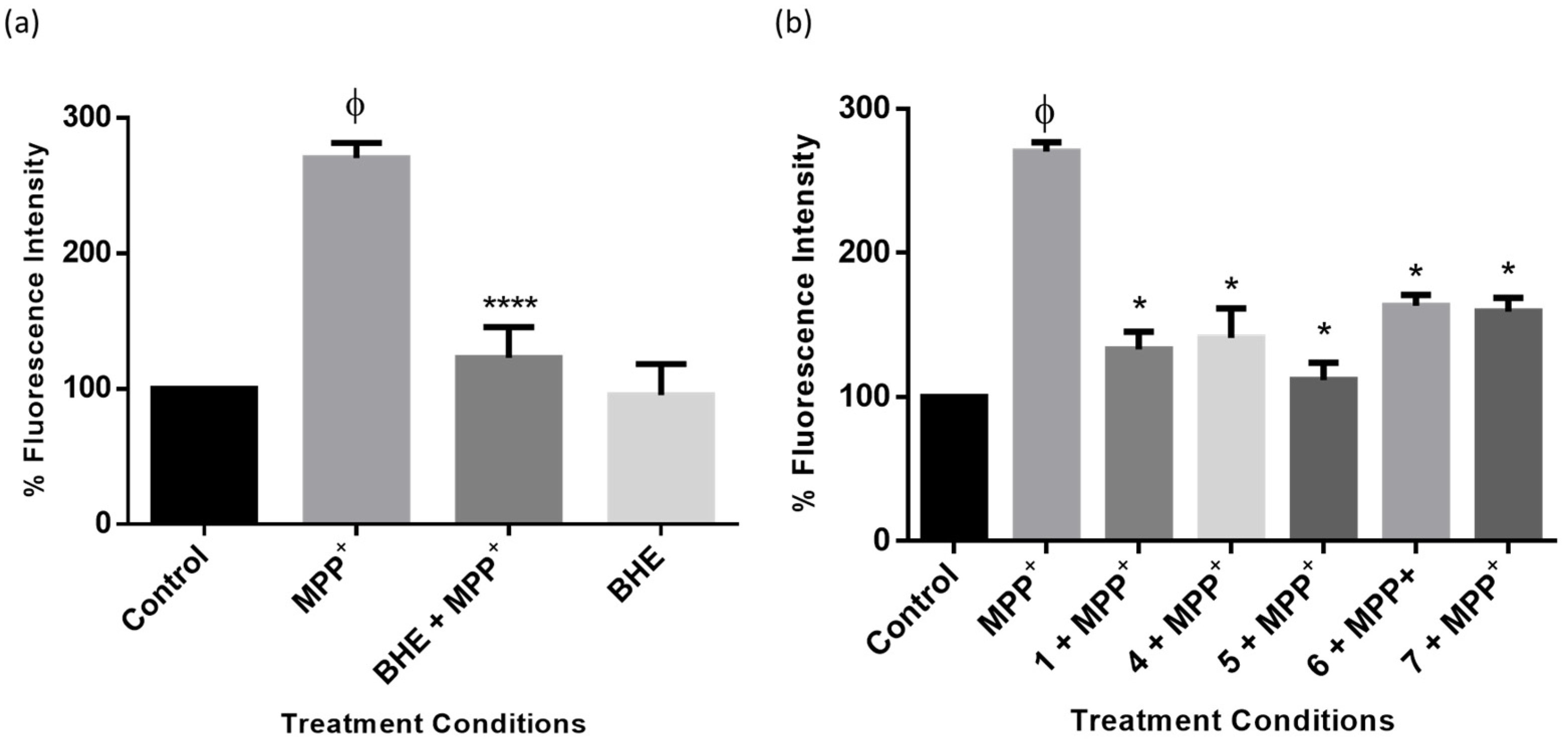
2.6. BHE and Isolated Compounds Inhibit MPP+-Induced Overproduction of ROS in SH-SY5Y Cells
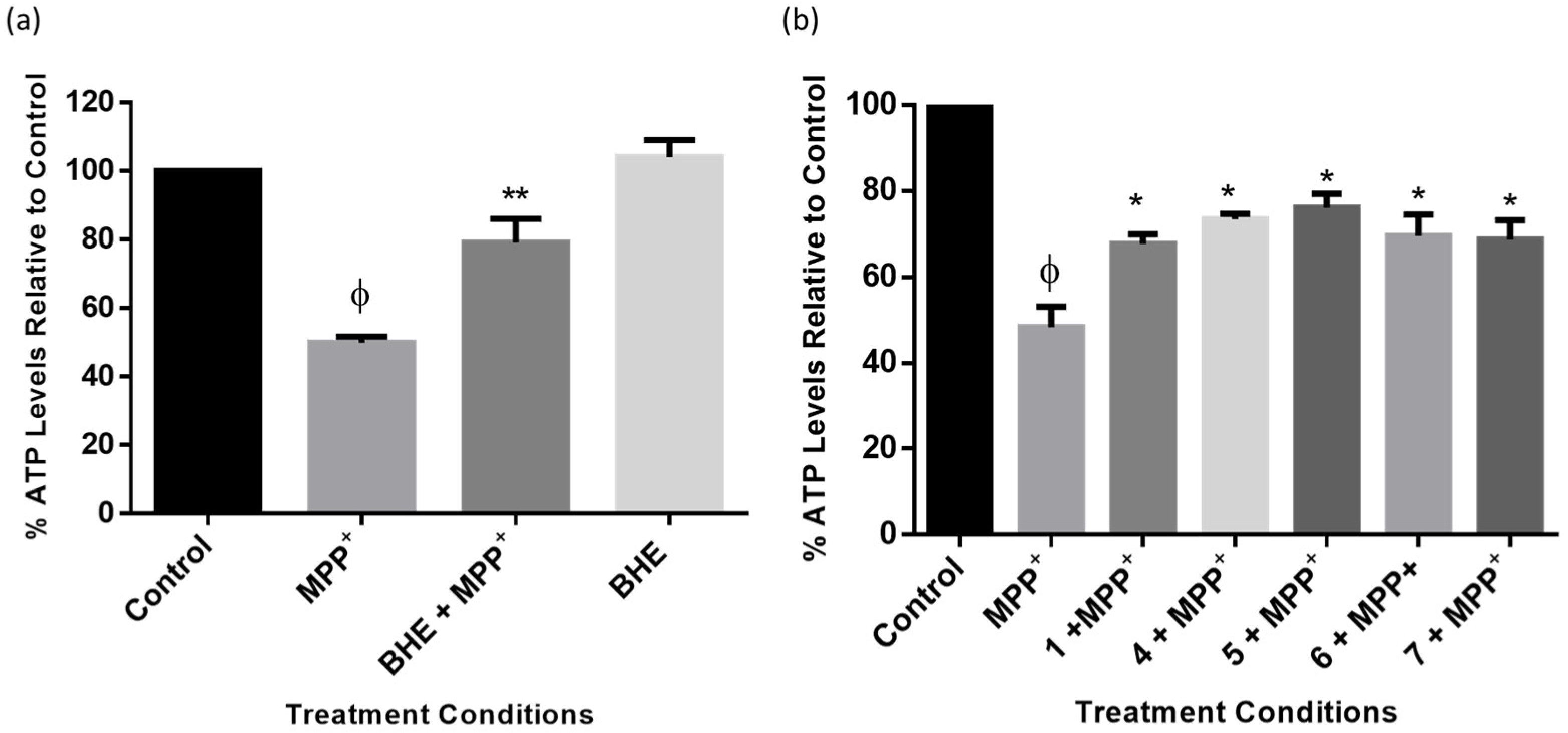
2.7. BHE and Isolated Compounds Mitigate MPP+-Induced ATP Depletion in SH-SY5Y Cells
2.8. BHE and Isolated Compounds Inhibit MPP+-Induced Apoptosis in SH-SY5Y Cells
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of Compounds
4.3. Physical and Spectroscopic Data of the Isolated Compounds
4.4. Cell Culture and Maintenance
4.5. Treatments
4.6. Cell Viability Assays
4.7. Cell Morphology
4.8. Adenosine Triphosphate Assay
4.9. Caspase 3/7 Apoptosis Assay
4.10. Determination of Intracellular ROS
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [Green Version]
- Lopes, F.M.; Schröder, R.; da Frota Júnior, M.L.C.; Zanotto-Filho, A.; Müller, C.B.; Pires, A.S.; Meurer, R.T.; Colpo, G.D.; Gelain, D.P.; Kapczinski, F. Comparison between proliferative and neuron-like SH-SY5Y cells as an in vitro model for Parkinson disease studies. Brain Res. 2010, 1337, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Wickremaratchi, M.M.; Perera, D.; O’Loghlen, C.; Sastry, D.; Morgan, E.; Jones, A.; Edwards, P.; Robertson, N.; Butler, C.; Morris, H.R. Prevalence and age of onset of Parkinson’s disease in Cardiff: A community based cross sectional study and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2009, 80, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Moriano, C.; Gonzalez-Burgos, E.; Gómez-Serranillos, M.P. Mitochondria-targeted protective compounds in Parkinson’s and Alzheimer’s diseases. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef]
- Onyango, I.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease. Neurochem. Res. 2008, 33, 589–597. [Google Scholar] [CrossRef]
- Requejo-Aguilar, R.; Bolaños, J.P. Mitochondrial control of cell bioenergetics in Parkinson’s disease. Free Radic. Biol. Med. 2016, 100, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Jenner, P.; Olanow, C.W. The pathogenesis of cell death in Parkinson’s disease. Neurology 2006, 66, S24–S36. [Google Scholar] [CrossRef]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Olanow, C.W.; Agid, Y.; Mizuno, Y.; Albanese, A.; Bonucelli, U.; Damier, P.; De Yebenes, J.; Gershanik, O.; Guttman, M.; Grandas, F. Levodopa in the treatment of Parkinson’s disease: Current controversies. Mov. Disord. 2004, 19, 997–1005. [Google Scholar] [CrossRef]
- Zhao, Y.; Luo, D.; Ning, Z.; Rong, J.; Lao, L. Electro-Acupuncture ameliorated MPTP-induced parkinsonism in mice via TrkB neurotrophic signaling. Front. Neurosci. 2019, 13, 496. [Google Scholar] [CrossRef]
- Ren, Z.-l.; Wang, C.-d.; Wang, T.; Ding, H.; Zhou, M.; Yang, N.; Liu, Y.-y.; Chan, P. Ganoderma lucidum extract ameliorates MPTP-induced parkinsonism and protects dopaminergic neurons from oxidative stress via regulating mitochondrial function, autophagy, and apoptosis. Acta Pharmacol. Sin. 2019, 40, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-Y.; Jeon, H.; Kim, H.; Koo, S.; Kim, S. Sophora flavescens Aiton Decreases MPP(+)-Induced Mitochondrial Dysfunction in SH-SY5Y Cells. Front. Aging Neurosci. 2018, 10, 119. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; He, X.; Wang, D. Lycopene protects against MPP+-induced cytotoxicity by maintaining mitochondrial function in SH-SY5Y cells. Neurochem. Res. 2013, 38, 1747–1757. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Jeon, H.; Ryu, S.; Koo, S.; Ha, K.-T.; Kim, S. Proteomic analysis of the effect of Korean red ginseng in the striatum of a Parkinson’s disease mouse model. PLoS ONE 2016, 11, e0164906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-C.; Liu, K.-F.; Teng, C.-J.; Lai, S.-C.; Yang, S.-E.; Ching, H.; Wu, C.-R. Sophora tomentosa extract prevents MPTP-induced parkinsonism in C57BL/6 mice via the inhibition of GSK-3β phosphorylation and oxidative stress. Nutrients 2019, 11, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrinkle, G. An introduction to the genus Boophane. Herbertia 1984, 40, 77–82. [Google Scholar]
- Meerow, A.; Snijman, D. Amaryllidaceae. In Flowering Plants·Monocotyledons; Springer: Berlin/Heidelberg, Germany, 1998; pp. 83–110. [Google Scholar]
- Meerow, A.W.; Snijman, D.A. Phylogeny of Amaryllidaceae tribe Amaryllideae based on nrDNA ITS sequences and morphology. Am. J. Bot. 2001, 88, 2321–2330. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.J.; Rárová, L.; Strnad, M.; Bastida, J.; Cheesman, L.; van Staden, J. Crinane alkaloids of the Amaryllidaceae with cytotoxic effects in human cervical adenocarcinoma (HeLa) cells. Nat. Prod. Commun. 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Neergaard, J.S.; Andersen, J.; Pedersen, M.E.; Stafford, G.I.; Staden, J.V.; Jäger, A.K. Alkaloids from Boophone disticha with affinity to the serotonin transporter. S. Afr. J. Bot. 2009, 75, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Cheesman, L.; Nair, J.J.; van Staden, J. Antibacterial activity of crinane alkaloids from Boophone disticha (Amaryllidaceae). J. Ethnopharmacol. 2012, 140, 405–408. [Google Scholar] [CrossRef]
- De Beer, J.J.J.; Van Wyk, B.E. An ethnobotanical survey of the Agter–Hantam, Northern Cape Province, South Africa. S. Afr. J. Bot. 2011, 77, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Hauth, H.; Stauffacher, D. The Alkaloids from Buphane disticha (L. f.) Herb. 2. Communication on amaryllidaceae alkaloids. Helv. Chim. Acta 1961, 44, 491–502. [Google Scholar] [CrossRef]
- Nair, J.; Manning, J.; Van Staden, J. Distichamine, a chemotaxonomic marker for the genus Boophone Herb (Amaryllidaceae). S. Afr. J. Bot. 2012, 83, 89–91. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.J.; Rárová, L.; Strnad, M.; Bastida, J.; van Staden, J. Alkaloids from Boophone haemanthoides (Amaryllidaceae). Nat. Prod. Commun. 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Lepule, K.H.; Cordier, W.; Steenkamp, P.; Nell, M.; Steenkamp, V. The ability of three African herbal remedies to offer protection against an in vitro model of Parkinson’s disease. S. Afr. J. Bot. 2019, 126, 121–131. [Google Scholar] [CrossRef]
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Ibrakaw, A.; Boatwright, J.; Lesch, T.; Cupido, C.; Hussein, A. Triterpenes and other minor chemical constituents of Boophone haemanthoides FM Leight (Amaryllidaceae). S. Afr. J. Bot. 2020, in press. [Google Scholar] [CrossRef]
- Robbins, D.W.; Boebel, T.A.; Hartwig, J.F. Iridium-catalyzed, silyl-directed borylation of nitrogen-containing heterocycles. J. Am. Chem. Soc. 2010, 132, 4068–4069. [Google Scholar] [CrossRef]
- Hauth, H.; Stauffacher, D. 89. Die Struktur von Acetylnerbowdin: 4. Mitteilung über Amaryllidaceen-Alkaloide. Helv. Chim. Acta 1963, 46, 810–812. [Google Scholar] [CrossRef]
- Cho, N.; Du, Y.; Valenciano, A.L.; Fernández-Murga, M.L.; Goetz, M.; Clement, J.; Cassera, M.B.; Kingston, D.G. Antiplasmodial alkaloids from bulbs of Amaryllis belladonna Steud. Bioorgan. Med. Chem. Lett. 2018, 28, 40–42. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of Amaryllidaceae alkaloids. In The Alkaloids: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 83, pp. 113–185. [Google Scholar]
- Enogieru, A.B.; Omoruyi, S.I.; Ekpo, O.E. Aqueous leaf extract of Sutherlandia frutescens attenuates ROS-induced apoptosis and loss of mitochondrial membrane potential in MPP+-treated SH-SY5Y cells. Trop. J. Pharm. Res. 2020, 19, 549–555. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Mitochondrial oxidative stress: Implications for cell death. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 143–183. [Google Scholar] [CrossRef] [PubMed]
- Limboonreung, T.; Tuchinda, P.; Chongthammakun, S. Chrysoeriol mediates mitochondrial protection via PI3K/Akt pathway in MPP+ treated SH-SY5Y cells. Neurosci. Lett. 2020, 714, 134545. [Google Scholar] [CrossRef] [PubMed]
- Höglinger, G.U.; Carrard, G.; Michel, P.P.; Medja, F.; Lombès, A.; Ruberg, M.; Friguet, B.; Hirsch, E.C. Dysfunction of mitochondrial complex I and the proteasome: Interactions between two biochemical deficits in a cellular model of Parkinson’s disease. J. Neurochem. 2003, 86, 1297–1307. [Google Scholar] [CrossRef]
- Stennicke, H.R.; Renatus, M.; Meldal, M.; Salvesen, G.S. Internally quenched fluorescent peptide substrates disclose the subsite preferences of human caspases 1, 3, 6, 7 and 8. Biochem. J. 2000, 350, 563–568. [Google Scholar] [CrossRef]
- Denault, J.-B.; Salvesen, G.S. Caspases: Keys in the ignition of cell death. Chem. Rev. 2002, 102, 4489–4500. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Rano, T.A.; Peterson, E.P.; Rasper, D.M.; Timkey, T.; Garcia-Calvo, M.; Houtzager, V.M.; Nordstrom, P.A.; Roy, S.; Vaillancourt, J.P. A combinatorial approach defines specificities of members of the caspase family and granzyme B Functional relationships established for key mediators of apoptosis. J. Biol. Chem. 1997, 272, 17907–17911. [Google Scholar] [CrossRef] [Green Version]
- Bressenot, A.; Marchal, S.; Bezdetnaya, L.; Garrier, J.; Guillemin, F.; Plénat, F. Assessment of apoptosis by immunohistochemistry to active caspase-3, active caspase-7, or cleaved PARP in monolayer cells and spheroid and subcutaneous xenografts of human carcinoma. J. Histochem. Cytochem. 2009, 57, 289–300. [Google Scholar] [CrossRef]
- Omoruyi, S.; Enogieru, A.; Hussein, A.; Ekpo, O. Preliminary investigation of the neuroprotective potentials of Crossyne guttata in MPP+-induced toxicity in SH-SY5Y. J. Afr. Assoc. Physiol. Sci. 2019, 7, 138–144. [Google Scholar]
- Omoruyi, S.I.; Delport, J.; Kangwa, T.S.; Ibrakaw, A.S.; Cupido, C.N.; Ekpo, O.E.; Hussein, A.A. In vitro neuroprotective potential of Clivia miniata and Nerine humilis (Amaryllidaceae) in MPP+-induced neuronal toxicity in SH-SY5Y neuroblastoma cells. S. Afr. J. Bot. 2020, in press. [Google Scholar] [CrossRef]
- Seoposengwe, K.; Van Tonder, J.J.; Steenkamp, V. In vitro neuroprotective potential of four medicinal plants against rotenone-induced toxicity in SH-SY5Y neuroblastoma cells. BMC Complement. Altern. Med. 2013, 13, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ago, Y.; Koda, K.; Takuma, K.; Matsuda, T. Pharmacological aspects of the acetylcholinesterase inhibitor galantamine. J. Pharmacol. Sci. 2011, 116, 6–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, I.H.; Park, Y.H.; Lee, S.I.; Yang, H.D.; Moon, H.-I. Neuroprotective activity of triterpenoid saponins from Platycodi radix against glutamate-induced toxicity in primary cultured rat cortical cells. Molecules 2007, 12, 1147–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, J.; Wang, X.; Song, C. Neuroprotective and antioxidant lanostanoid triterpenes from the fruiting bodies of Ganoderma atrum. Fitoterapia 2016, 109, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.; Cooper, J.; Dexter, D.; Clark, J.; Jenner, P.; Marsden, C. Mitochondrial complex I deficiency in Parkinson’s disease. J. Neurochem. 1990, 54, 823–827. [Google Scholar] [CrossRef]
- Keeney, P.M.; Xie, J.; Capaldi, R.A.; Bennett, J.P. Parkinson’s disease brain mitochondrial complex I has oxidatively damaged subunits and is functionally impaired and misassembled. J. Neurosci. 2006, 26, 5256–5264. [Google Scholar] [CrossRef]
- Henchcliffe, C.; Beal, M.F. Mitochondrial biology and oxidative stress in Parkinson disease pathogenesis. Nat. Clin. Pract. Neurol. 2008, 4, 600–609. [Google Scholar] [CrossRef]
- Su, Y.H.; Lee, Y.L.; Chen, S.F.; Lee, Y.P.; Hsieh, Y.H.; Tsai, J.H.; Hsu, J.L.; Tian, W.T.; Huang, W. Essential role of β-human 8-oxoguanine DNA glycosylase 1 in mitochondrial oxidative DNA repair. Environ. Mol. Mutagenesis 2013, 54, 54–64. [Google Scholar] [CrossRef]
- Shults, C.W.; Oakes, D.; Kieburtz, K.; Beal, M.F.; Haas, R.; Plumb, S.; Juncos, J.L.; Nutt, J.; Shoulson, I.; Carter, J. Effects of coenzyme Q10 in early Parkinson disease: Evidence of slowing of the functional decline. Arch. Neurol. 2002, 59, 1541–1550. [Google Scholar] [CrossRef]
- Paraskevas, G.P.; Kapaki, E.; Petropoulou, O.; Anagnostouli, M.; Vagenas, V.; Papageorgiou, C. Plasma levels of antioxidant vitamins C and E are decreased in vascular parkinsonism. J. Neurol. Sci. 2003, 215, 51–55. [Google Scholar] [CrossRef]
- Kieburtz, K. A randomized clinical trial of coenzyme Q10 and GPI-1485 in early Parkinson’s disease: P574. Mov. Disord. 2006, 21, 20–28. [Google Scholar]
- Baunthiyal, M.; Singh, V.; Dwivedi, S. Insights of antioxidants as molecules for drug discovery. Int. J. Pharmacol. 2017, 13, 874–889. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Ding, J.-H.; Zhou, F.; Wang, Z.-Y.; Zhou, X.-Q.; Hu, G. Iptakalim ameliorates MPP+-induced astrocyte mitochondrial dysfunction by increasing mitochondrial complex activity besides opening mitoKATP channels. J. Neurosci. Res. 2009, 87, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.W.-L.; Liu, H.-F.; Ho, J.W.-M.; Zhang, W.-Y.; Chu, A.C.-Y.; Kwok, K.H.-H.; Ge, X.; Chan, K.-H.; Ramsden, D.B.; Ho, S.-L. Mitochondrial uncoupling protein-2 (UCP2) mediates leptin protection against MPP+ toxicity in neuronal cells. Neurotox. Res. 2010, 17, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Imamura, H.; Sasaoka, N.; Yamamoto, M.; Uemura, N.; Shudo, T.; Fuchigami, T.; Takahashi, R.; Kakizuka, A. ATP maintenance via two types of ATP regulators mitigates pathological phenotypes in mouse models of Parkinson’s disease. EBioMedicine 2017, 22, 225–241. [Google Scholar] [CrossRef] [Green Version]
- Di Monte, D.; Sandy, M.S.; Ekström, G.; Smith, M.T. Comparative studies on the mechanisms of paraquat and 1-methyl-4-phenylpyridine (MPP+) cytotoxicity. Biochem. Biophys. Res. Commun. 1986, 137, 303–309. [Google Scholar] [CrossRef]
- Kehrer, J.P. Free radicals in biology: Sources, reactivities, and roles in the etiology of human diseases. Nat. Antioxid. Hum. Health Dis. 1994, 30, 25–62. [Google Scholar]
- Lee, D.-H.; Kim, C.-S.; Lee, Y.J. Astaxanthin protects against MPTP/MPP+-induced mitochondrial dysfunction and ROS production in vivo and in vitro. Food Chem. Toxicol. 2011, 49, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Nicotera, P.; Leist, M.; Fava, E.; Berliocchi, L.; Volbracht, C. Energy requirement for caspase activation and neuronal cell death. Brain Pathol. 2000, 10, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, R.-Y.; Zhao, J.; Dong, Z.; Feng, D.-Y.; Wu, R.; Shi, M.; Zhao, G. Ginsenoside Rd protects SH-SY5Y cells against 1-methyl-4-phenylpyridinium induced injury. Int. J. Mol. Sci. 2015, 16, 14395–14408. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Eguchi, Y.; Imagawa, Y.; Akai, S.; Mochizuki, H.; Tsujimoto, Y. MPP+ induces necrostatin-1- and ferrostatin-1-sensitive necrotic death of neuronal SH-SY5Y cells. Cell Death Discov. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds 1, 2, 3, 4, 5, 6 and 7 as well as the plant extracts and fractions are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 1 | Compound 2 | ||||
|---|---|---|---|---|---|
| δC | δH (JHz) | δC | δH (JHz) | HMBC (H→C) | |
| 1 | 198.7 s | 68.3 d | 5.71 br t (2.6) | C2, C10b, C4a, C3, CO | |
| 2 | 102.0 d | 5.37 d (1.1) | 28.6 t | 1.95 * (2α) | |
| 2.29 dq (2.6, 16.2) (2β) | C4, C10b, C4a, C1, C3 | ||||
| 3 | 173.2 s | 68.0 d | 5.15 br quint (2.6) | C2, C4, C4a, C1, C3 | |
| 4-α | 30.3 t | 2.49 dd (17.3, 6.7) | 30.5 t | 2.08 * | |
| 4-β | 2.38 ddd (17.3, 11.5, 1.1) | 1.46 ddd (3.3, 12.2, 15.5) | C10b, C4a | ||
| 4a | 66.1 d | 3.51 dd (10.8, 6.8) | 59.9 d | 3.48 dd (5.5, 12.2) | C4, C11, C12, C6, C10a |
| 6-α | 57.7 t | 4.12 d (17.4) | 57.5 t | 4.13 d (17.3) | C11, C12, C10, C6a, C8, C10a, C7, C9 |
| 6-β | 3.73 d (17.4) | 3.76 d (17.3) | C12, C4a, C6a, C8, C10a, C7, C9 | ||
| 6a | 116.6 s | 117.2 s | |||
| 7 | 139.8 s | 140.4 s | |||
| 8 | 135.3 s | 133.2 s | |||
| 9 | 147.5 s | 148.4 s | |||
| 10 | 100.2 d | 7.71 s | 97.1 d | 6.11 s | C10a, C6a, C10b, C7, C9 |
| 10a | 133.4 s | 137.2 s | |||
| 10b | 49.8 s | 46.8 s | |||
| 11 exo | 41.4 t | 2.17 ddd (13.0, 10.4, 6.7) | 38.2 t | 1.99 * | |
| 11 endo | 2.30 ddd (12.8, 8.5, 3.6) | 1.88 ** | |||
| 12 exo | 52.5 t | 3.38 ddd (13.0, 10.4, 3.6) | 51.0 t | 3.30 ddd (13.6, 10, 3.2) | C6 |
| 12 endo | 2.86 ddd (15.0, 13.4, 6.7) | 2.76 ddd (13.6, 9.0, 6.3) | C4a, C11, C6 | ||
| 1-COCH3 | 170.5 s | ||||
| 1-COCH3 | 21.2 q | 1.90 ** s | CO(C1) | ||
| 3-COCH3 | 170.1 s | ||||
| 3-COCH3 | 21.3 q | 1.97 * s | CO(C3) | ||
| 3-OCH3 | 55.6 q | 3.74 (s) | |||
| 7-OCH3 | 58.8 q | 3.95 (s) | 59.1 q | 3.95 s | C7 |
| OCH2O | 100.3 t | 5.84/5.85, d/each (1.5) | 100.5 t | 5.83/;5.79 d/each (1.3) | C8, C9 |
| Compound | Name |
|---|---|
| 1 | Distachamine |
| 2 | 1α,3α-Diacetylnerbowdine |
| 3 | Hippadine |
| 4 | Stigmast-4-ene-3,6-dione |
| 5 | Cholest-4-en-3-one |
| 6 | Tyrosol |
| 7 | 3-Hydroxy-1-(4′-hydroxyphenyl)-1-propanone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrakaw, A.S.; Omoruyi, S.I.; Ekpo, O.E.; Hussein, A.A. Neuroprotective Activities of Boophone haemanthoides (Amaryllidaceae) Extract and Its Chemical Constituents. Molecules 2020, 25, 5376. https://doi.org/10.3390/molecules25225376
Ibrakaw AS, Omoruyi SI, Ekpo OE, Hussein AA. Neuroprotective Activities of Boophone haemanthoides (Amaryllidaceae) Extract and Its Chemical Constituents. Molecules. 2020; 25(22):5376. https://doi.org/10.3390/molecules25225376
Chicago/Turabian StyleIbrakaw, Abobaker S., Sylvester I. Omoruyi, Okobi E. Ekpo, and Ahmed A. Hussein. 2020. "Neuroprotective Activities of Boophone haemanthoides (Amaryllidaceae) Extract and Its Chemical Constituents" Molecules 25, no. 22: 5376. https://doi.org/10.3390/molecules25225376