Changes in Cuticle Components and Morphology of ‘Satsuma’ Mandarin (Citrus unshiu) during Ambient Storage and Their Potential Role on Penicillium digitatum Infection

Abstract
:1. Introduction
2. Results and Discussion
2.1. Postharvest Decay and Weight Loss
2.2. Changes in Cuticular Fraction Contents of the Fruits during Storage
2.3. Changes in Epicuticular Wax Compositions during Storage
2.4. Changes in Intracuticular Wax Compositions during Storage
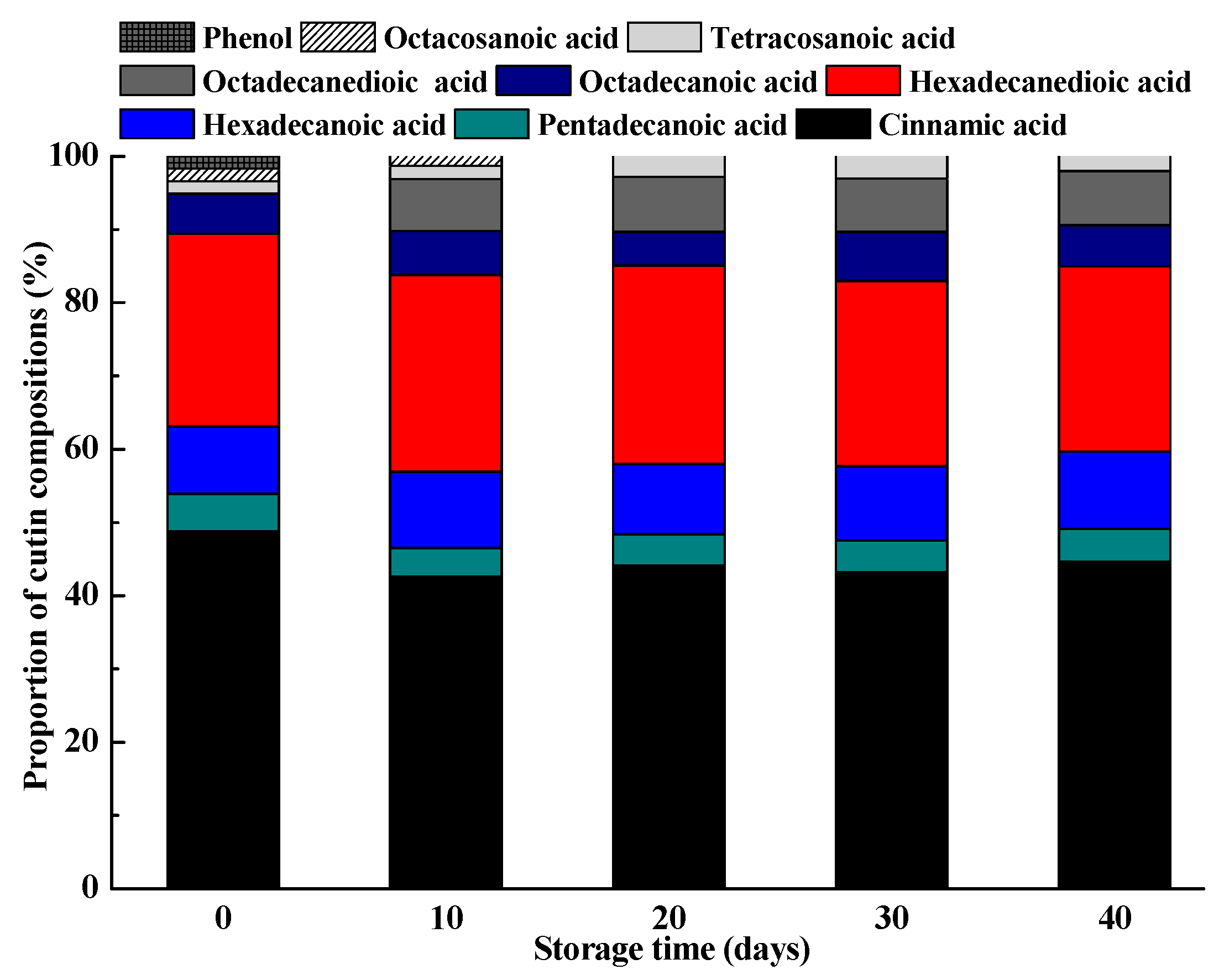
2.5. Changes in Cutin Monomers during Storage
2.6. Changes in Crystal Structure of Epicuticular Wax during Storage
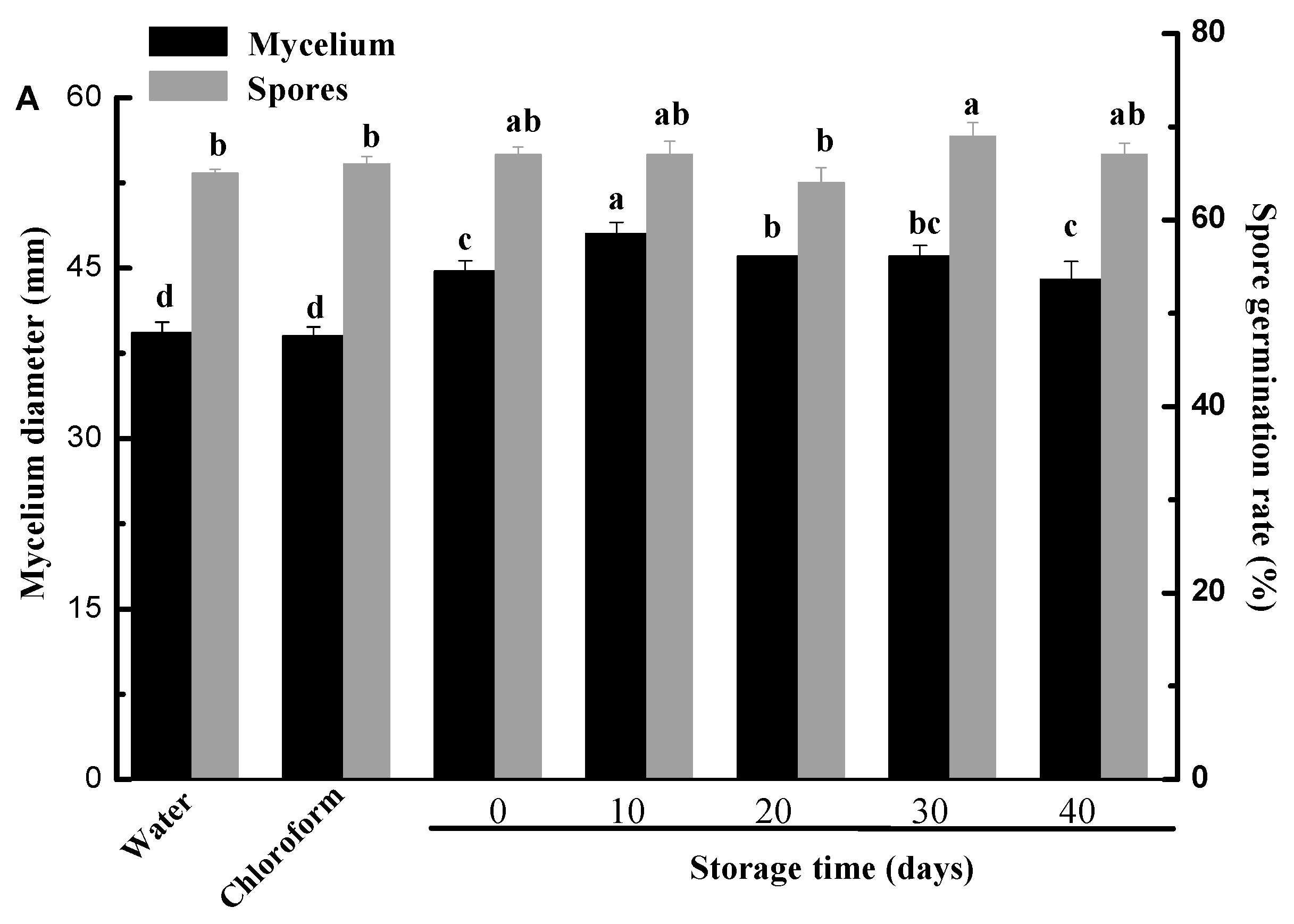
2.7. Effect of Different Cuticle Extracts on Mycelium Growth and Conidial Germination of Penicillium digitatum In Vitro
3. Materials and Methods
3.1. Experimental Materials and Pathogen
3.2. Determination of Pericarp Surface Area
3.3. Cuticle Extraction and Analysis
3.3.1. Epicuticular and Intracuticular Wax Extraction
3.3.2. Epicuticular and Intracuticular Wax Analysis
3.4. Cutin Isolation and Its Monomers Analysis
3.5. Scanning Electron Microscopy Observation
3.6. Effect of Different Cuticle Extracts on Mycelium Growth and Conidial Germination of Penicillium digitatum In Vitro
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1-MCP | 1-methylcyclopropene |
| BSTFA | bis-N,N-(trimethylsilyl)-trifluoroacetamide |
| EGCG | epigallocatechin-3-gallate |
| GC-MS | gas chromatography-mass spectrometry |
| ROS | reactive oxygen species |
References
- Belge, B.; Llovera, M.; Comabella, E.; Gatius, F.; Guillén, P.; Graell, J.; Lara, I. Characterization of cuticle composition after cold storage of “Celeste” and “Somerset” sweet cherry fruit. J. Agric. Food Chem. 2014, 62, 8722–8729. [Google Scholar] [CrossRef]
- Koch, K.; Ensikat, H.J. The hydrophobic coatings of plant surfaces: Epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly. Micron 2008, 39, 759–772. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Lara, I.; Belge, B.; Goulao, L.F. The fruit cuticle as a modulator of postharvest quality. Postharvest Biol. Technol. 2014, 87, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Sala, J.M.; Lafuente, T.; Cuñat, P. Content and chemical composition of epicuticular wax of ‘Navelina’ oranges and ‘Satsuma’ mandarins as related to rindstaining of fruit. J. Sci. Food Agric. 1992, 59, 489–495. [Google Scholar] [CrossRef]
- Wang, J.Q.; Hao, H.H.; Liu, R.S.; Ma, Q.L.; Xu, J.; Chen, F.; Cheng, Y.J.; Deng, X.X. Comparative analysis of surface wax in mature fruits between Satsuma mandarin (Citrus unshiu) and ‘Newhall’ navel orange (Citrus sinensis) from the perspective of crystal morphology, chemical composition and key gene expression. Food Chem. 2014, 153, 177–185. [Google Scholar] [CrossRef]
- Liu, D.C.; Zeng, Q.; Ji, Q.X.; Liu, C.F.; Liu, S.B.; Liu, Y. A comparison of the ultrastructure and composition of fruits’ cuticular wax from the wild-type ‘Newhall’ navel orange (Citrus sinensis [L] Osbeck cv. Newhall) and its glossy mutant. Plant Cell Rep. 2012, 31, 2239–2246. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.C.; Li, Y.; Zheng, Q.; Wang, Y.C.; Wang, M.L.; Zhuang, X.; Wu, Q.; Liu, C.F.; Liu, S.H.; Liu, Y. Analysis of cuticular wax constituents and genes that contribute to the formation of ‘glossy Newhall’, a spontaneous bud mutant from the wild-type ‘Newhall’ navel orange. Plant Mol. Biol. 2015, 88, 573–590. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.C.; Yang, L.; Wang, Y.C.; Zhuang, X.; Liu, C.F.; Liu, S.B.; Liu, Y. Transcriptome sequencing identified wax-related genes controlling the glossy phenotype formation of “Ganqi 3,” a bud mutant derived from wild-type “Newhall” navel orange. Tree Genet. Genomes 2016, 12, 55. [Google Scholar] [CrossRef]
- Wang, J.Q.; Li, S.; Li, X.; He, Y.Z.; Luo, T.; Sheng, L.; Luo, Y.; Zeng, Y.L.; Xu, J.; Deng, X.X.; et al. Regulation of cuticle formation during fruit development and ripening in ‘Newhall’ navel orange (Citrus sinensis Osbeck) revealed by transcriptomic and metabolomic profiling. Plant Sci. 2016, 243, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Zhou, B.; Zhang, J.; Wang, C.; Liu, C.H.; Liu, Y.L.; Zhu, X.B.; Ren, X.L. Relationships between cuticular waxes and skin greasiness of apple during storage. Postharvest Biol. Technol. 2017, 131, 55–67. [Google Scholar] [CrossRef]
- Belge, B.; Llovera, M.; Comabella, E.; Graell, J.; Lara, I. Fruit cuticle composition of a melting and nonmelting peach cultivar. J. Agric. Food Chem. 2014, 62, 3488–3495. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yin, H.; Chen, Y.Y.; Li, L.; Wang, Y.Z.; Hao, P.P.; Cao, P.; Qi, K.J.; Zhang, S.L. Chemical composition, crystal morphology and key gene expression of cuticular waxes of Asian pears at harvest and after storage. Postharvest Biol. Technol. 2017, 132, 71–80. [Google Scholar] [CrossRef]
- Chu, W.J.; Gao, H.Y.; Chen, H.J.; Wu, W.J.; Fang, X.J. Changes in cuticular wax composition of two blueberry cultivars during fruit ripening and postharvest cold storage. J. Agric. Food Chem. 2018, 66, 2870–2876. [Google Scholar] [CrossRef]
- Li, F.J.; Min, D.D.; Ren, C.T.; Dong, L.L.; Shu, P.; Cui, X.X.; Zhang, X.H. Ethylene altered fruit cuticular wax, the expression of cuticular wax synthesis-related and fruit quality during cold storage of apple (Malus domestica Borkh. c.v. Starkrimson) fruit. Postharvest Biol. Technol. 2019, 149, 58–65. [Google Scholar] [CrossRef]
- Yan, D.; Yang, Y.Q.; Wang, C.; Qi, Y.W.; Liu, C.H.; Zhou, B.; Ren, X.L. Effects of epigallocatechin-3-gallate (EGCG) on skin greasiness and related gene expression in ‘Jonagold’ apple fruit during ambient storage. Postharvest Biol. Technol. 2018, 143, 28–34. [Google Scholar] [CrossRef]
- Belge, B.; Goulao, L.F.; Comabella, E.; Graell, J.; Lara, I. Postharvest heat and CO2 shocks induce changes in cuticle composition and cuticle-related gene expression in ‘October Sun’ peach fruit. Postharvest Biol. Technol. 2019, 148, 200–207. [Google Scholar] [CrossRef]
- Silva-Moreno, E.; Brito-Echeverŕıa, J.; López, M.; Ríos, J.; Balic, I.; Campos-Vargas, R.; Polanco, R. Effect of cuticular waxes compounds from table grapes on growth, germination and gene expression in Botrytis cinerea. World J. Microb. Biot. 2016, 74, 2–6. [Google Scholar] [CrossRef]
- Feng, J.; Wang, F.; Liu, G.S.; Greenshields, D.; Shen, W.Y.; Kaminskyj, S.; Hughes, G.R.; Peng, Y.L.; Selvaraj, G.; Zou, J.T.; et al. Analysis of a Blumeria graminis-secreted lipase reveals the importance of host epicuticular wax components for fungal adhesion and development. Mol. Plant Microbe Interact. 2009, 22, 1601–1610. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.T.; Batt, R.F.; Burchill, R.T. Fungistatic properties of apple leaf wax. Nature 1957, 180, 796–797. [Google Scholar] [CrossRef]
- Yin, Y.; Bi, Y.; Chen, S.J.; Li, Y.C.; Wang, Y.; Ge, Y.H.; Ding, B.; Li, Y.C.H.; Zhang, Z. Chemical composition and antifungal activity of cuticular wax isolated from Asian pear fruit (cv. Pingguoli). Sci. Hortic. 2011, 129, 577–582. [Google Scholar] [CrossRef]
- Li, Y.C.; Yin, Y.; Chen, S.J.; Bi, Y.; Ge, Y.H. Chemical composition of cuticular waxes during fruit development of Pingguoli pear and their potential role on early events of Alternaria alternata infection. Funct. Plant Biol. 2014, 41, 313–320. [Google Scholar] [CrossRef]
- Zabka, V.; Stangl, M.; Bringmann, G.; Vogg, G.; Riederer, M.; Hildebrandt, U. Host surface properties affect prepenetration processes in the barley powdery mildew fungus. New Phytol. 2008, 177, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, Y.C.; Bi, Y.; Wang, Y. Role of pear fruit cuticular wax and surface hydrophobicity in regulating the prepenetration phase of Alternaria alternata infection. J. Phytopathol. 2017, 165, 313–322. [Google Scholar] [CrossRef]
- Hansjakob, A.; Bischof, S.; Bringmann, G.; Riederer, M.; Hildebrandt, U. Very-long-chain aldehydes promote in vitro prepenetration processes of Blumeria graminis in a dose- and chain length-dependent manner. New Phytol. 2010, 188, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Hansjakob, A.; Riederer, M.; Hildebrandt, U. Appressorium morphogenesis and cell cycle progression are linked in the grass powdery mildew fungus Blumeria graminis. Fungal Biol. 2012, 116, 890–901. [Google Scholar] [CrossRef]
- Prusky, D.; McEvoy, J.L.; Saftner, R.; Conway, W.S.; Jones, R. Relationship between host acidification and virulence of Penicillium spp. on apple and citrus fruit. Phytopathology 2004, 94, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Macarisin, D.; Cohen, L.; Eick, A.; Rafael, G.; Belausov, E.; Wisniewski, M.; Droby, S. Penicillium digitatum suppresses production of hydrogen peroxide in host tissue during infection of citrus fruit. Phytopathology 2007, 97, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- González-Candelas, L.; Alamar, S.; Sánchez-Torres, P.; Zacarías, L.; Marcos, J.F. A transcriptomic approach highlights induction of secondary metabolism in citrus fruit in response to Penicillium digitatum infection. BMC Plant Biol. 2010, 10, 194–195. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.H.; Zhang, J.; Wang, R.R.; Ou, S.Y.; Shan, Y. Changes in cuticle compositions and crystal structure of ‘Bingtang’ sweet orange fruits (Citrus sinensis) during storage. Int. J. Food Prop. 2018, 21, 2411–2427. [Google Scholar] [CrossRef] [Green Version]
- Li, F.J.; Min, D.D.; Song, B.C.; Shao, S.J.; Zhang, X.H. Ethylene effects on apple fruit cuticular wax composition and content during cold storage. Postharvest Biol. Technol. 2017, 134, 98–105. [Google Scholar] [CrossRef]
- Domínguez, E.; López-Casado, G.; Guartero, J.; Heredia, A. Development of fruit cuticle in cherry tomato (Solanum lycopersicum). Func. Plant Biol. 2008, 35, 403–411. [Google Scholar] [CrossRef]
- Huang, H.; Burghardt, M.; Schuster, A.C.; Leide, J.; Lara, I. Chemical composition and water permeability of fruit and leaf cuticles of Olea europaea L. J. Agric. Food Chem. 2017, 65, 8790–8797. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, S.; Ozaki, Y.; Yonemori, K.; Azuma, J. Mechanical properties of fruit-cuticular membranes isolated from 27 cultivars of Diospyros kaki Thunb. Food Chem. 2012, 132, 2135–2139. [Google Scholar] [CrossRef]
- Isaacaon, T.; Kosma, D.K.; Matas, A.J. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 609, 363–377. [Google Scholar] [CrossRef]
- L’Haridon, F.; Besson-Bard, B.; Binda, M.; Serrano, M.; Abou-Mansour, E.; Balet, F.; Schoonbeek, H.; Hess, S.; Mir, R.; Léon, J.; et al. A permeable cuticle is associated with the release of reactive oxygen species and induction of innate immunity. PLoS Pathog. 2011, 7, e1002148. [Google Scholar] [CrossRef]
- Dong, X.Q.; Rao, J.P.; Huber, D.J.; Chang, X.X.; Xin, F.C. Wax composition of ‘Red Fuji’ apple fruit during development and storage after 1-methylcyclopropene treatment. Hortic. Environ. Biotechnol. 2012, 53, 288–297. [Google Scholar] [CrossRef]
- Buschhaus, C.; Herz, H.; Jetter, R. Chemical composition of the epicuticular and intracuticular wax layers on the adaxial side of Ligustrum vulgare leaves. New Phytol. 2007, 176, 311–316. [Google Scholar] [CrossRef]
- Vogg, G.; Fischer, S.; Leide, J.; Emmanuel, E.; Jetter, R.; Levy, A.A.; Riederer, M. Tomato fruit cuticular waxes and their effects on transpiration barrier properties: Functional characterization of a mutant deficient in a very-long-chain fatty acid β-ketoacyl-CoA synthase. J. Exp. Bot. 2004, 55, 1401–1410. [Google Scholar] [CrossRef] [Green Version]
- Buschhaus, C.; Jetter, R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Leide, J.; Hildebrandt, U.; Reussing, K.; Riederer, M.; Vogg, G. The developmental pattern of tomato fruit wax accumulation and its impact on cuticular transpiration barrier properties: Effects of a deficiency in a β-ketoacy-coenzyme A synthase (LeCER6). Plant Physiol. 2007, 144, 1667–1679. [Google Scholar] [CrossRef] [Green Version]
- Szakiel, A.; Pączkowski, C.; Pensec, F.; Bertsch, C. Fruit cuticular waxes as a source of biologically active tripenoids. Phytochem. Rev. 2012, 11, 263–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, E.A.; Procopiou, J. The cuticles of Citrus species. Compostion of the intracuticular lipids of leaves and fruits. J. Sci. Food Agric. 1975, 26, 1347–1352. [Google Scholar] [CrossRef]
- Ding, Y.; Chang, J.; Ma, Q.; Chen, L.; Liu, S.; Jin, S.; Han, J.; Xu, R.; Zhu, A.; Guo, J. Network analysis of postharvest senescence process in citrus fruits revealed by transcriptomic and metabolomic profiling. Plant Physiol. 2015, 168, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Pararsons, E.P.; Popopvsky, S.; Lohrey, G.T.; Alkalai-tuvia, S.; Perzelan, Y.; Bosland, P.; Bebeli, P.; Paran, I.; Fallik, E.; Jenks, M.A. Fruit cuticle lipid composition and water loss in a diverse collection of pepper (Capsicum). Physiol. Plant. 2013, 149, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Marga, F.; Pesacreta, T.C.; Hasenstein, K.G. Biochemical analysis of elastic and rigid cuticles of Cirsium horridulu. Planta 2001, 213, 841–848. [Google Scholar] [CrossRef]
- Cajuste, J.F.; González-Candelas, L.; Veyrat, A.; García-Breijo, F.J.; Reig-Armiñana, J.; Lafuente, M.T. Epicuticular wax content and morphology as related to ethylene and storage performance of ‘Navelate’ orange fruit. Postharvest Biol. Technol. 2010, 55, 29–35. [Google Scholar] [CrossRef] [Green Version]
- López-Castaňeda, J.; Corrales-García, J.; Terrazas-Salgado, T.; Colinas-León, T. Effect of vapor heat treatments on weight loss reduction and epicuticular changes in six varieties of cactus pear fruit. J. PACD 2010, 12, 37–47. [Google Scholar]
- Naziri, E.; Mantzouridou, F.; Tsimidou, M.Z. Enhanced squalene production by wild-type Saccharomyces cerevisiae strains using safe chemical means. J. Agric. Food Chem. 2011, 59, 9980–9989. [Google Scholar] [CrossRef]
- Liu, P.; Luo, L.; Guo, J.H.; Liu, H.M.; Wang, B.Q.; Deng, B.X.; Long, C.A.; Cheng, Y.J. Farnesol induces apoptosis and oxidative stress in the fungal pathogen Penicillium expansum. Mycologia 2010, 102, 311–318. [Google Scholar] [CrossRef]
- Łaźniewska, J.; Macioszek, V.K.; Kononowicz, A.K. Plant-fungus interface: The role of surface structures in plant resistance and susceptibility to pathogenic fungi. Physiol. Mol. Plant Pathol. 2012, 78, 24–30. [Google Scholar] [CrossRef]
- Hegde, Y.; Kolattukudy, P.E. Cuticular waxes relieve self-inhibition of germination and appressorium formation by the conidia of Magnaporthe grisea. Physiol. Mol. Plant Pathol. 1997, 51, 75–84. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cuticle Fractions | Cuticular Fraction Contents (μg cm−2) | ||||
|---|---|---|---|---|---|
| Harvest | 10 days | 20 days | 30 days | 40 days | |
| Epicuticular wax | 1.11 ± 0.18 c | 3.22 ± 0.25 b | 4.21 ± 0.39 a | 3.52 ± 0.63 b | 1.35 ± 0.02 d |
| Intracuticular wax | 4.78 ± 0.19 b | 3.70 ± 0.14 c | 6.09 ± 0.30 a | 5.00 ± 0.26 b | 3.66 ± 0.55 c |
| Total wax | 5.89 ± 0.21 d | 6.92 ± 0.28 c | 10.30 ± 0.40 a | 8.52 ± 0.74 b | 5.01 ± 0.59 d |
| Cutin | 52.50 ± 1.79 a | 38.10 ± 0.78 b | 39.00 ± 1.16 b | 27.80 ± 2.54 c | 28.21 ± 5.25 c |
| Compositions | Contents of Epicuticular Wax Constituents (μg cm−2) | ||||
|---|---|---|---|---|---|
| Harvest | 10 days | 20 days | 30 days | 40 days | |
| Acids | 0.73 ± 0.01 c | 2.25 ± 0.26 b | 3.03 ± 0.11 a | 2.63 ± 0.18 b | 0.70 ± 0.03 c |
| Dodecanoic acid | 0.03 ± 0.01 b | 0.04 ± 0.00 b | 0.07 ± 0.01 a | 0.11 ± 0.04 a | - |
| Tetradecanoic acid | 0.05 ± 0.01 b | 0.10 ± 0.01 a | 0.10 ± 0.02 a | - | - |
| Pentadecanoic acid | 0.02 ± 0.01 | - | - | - | - |
| cis-9-Hexadecenoic acid | 0.02 ± 0.01 b | 0.07 ± 0.03 a | 0.11 ± 0.05 a | 0.11 ± 0.01 a | - |
| Hexadecanoic acid | 0.40 ± 0.02 c | 0.80 ± 0.06 b | 1.02 ± 0.13 a | 0.80 ± 0.07 b | 0.31 ± 0.03 c |
| 9,12-Octadecanoic acid | 0.02 ± 0.01 b | 0.07 ± 0.03 a | 0.11 ± 0.03 a | 0.08 ± 0.02 a | - |
| Oleic acid | 0.03 ± 0.00 b | 0.16 ± 0.01 a | 0.16 ± 0.07 a | 0.12 ± 0.04 a | - |
| Octadecanoic acid | 0.12 ± 0.01 e | 0.35 ± 0.07 c | 0.72 ± 0.08 a | 0.49 ± 0.03 b | 0.18 ± 0.01 d |
| Eicosanoic acid | 0.04 ± 0.02 d | 0.66 ± 0.11 b | 0.74 ± 0.13 a,b | 0.92 ± 0.19 a | 0.21 ± 0.03 c |
| Alkanes | 0.12 ± 0.01 d | 0.45 ± 0.05 b | 0.68 ± 0.03 a | 0.54 ± 0.06 b | 0.35 ± 0.02 c |
| Heptadecane | 0.04 ± 0.02 | - | - | - | - |
| Heptacosane | 0.02 ± 0.01 c | 0.18 ± 0.09 a | 0.23 ± 0.05 a | 0.22 ± 0.04 a | 0.10 ± 0.02 b |
| Nonacosane | 0.02 ± 0.01 d | 0.27 ± 0.10 b,c | 0.45 ± 0.04 a | 0.32 ± 0.04 b | 0.25 ± 0.01 c |
| Hentriacontane | 0.04 ± 0.01 | - | - | - | - |
| Terpenoids | 0.17 ± 0.01 a | 0.12 ± 0.06 a,b | 0.20 ± 0.07 a | 0.15 ± 0.01 a | 0.10 ± 0.01 b |
| Squalene | 0.02 ± 0.00 | - | - | - | - |
| Friedelin | 0.15 ± 0.02 a | 0.12 ± 0.06 a,b | 0.20 ± 0.07 a | 0.15 ± 0.01 a | 0.10 ± 0.01 b |
| Compositions | Contents of Intracuticular Wax Constituents (μg cm−2) | ||||
|---|---|---|---|---|---|
| Harvest | 10 days | 20 days | 30 days | 40 days | |
| Acids | 1.71 ± 0.02 a | 0.72 ± 0.05 d | 1.15 ± 0.07 b | 0.83 ± 0.03 c | 0.69 ± 0.02 d |
| Dodecanoic acid | 0.03 ± 0.00 | - | - | - | - |
| Tetradecanoic acid | 0.13 ± 0.01 a | 0.02 ± 0.01 b,c | 0.01 ± 0.01 c | 0.04 ± 0.01 b | 0.03 ± 0.01 b |
| Pentadecanoic acid | 0.07 ± 0.01 | - | - | - | - |
| cis-9-Hexadecenoic acid | 0.15 ± 0.01 a | 0.01 ± 0.00 c | 0.02 ± 0.01 b,c | 0.03 ± 0.01 b | 0.02 ± 0.01 b,c |
| Hexadecanoic acid | 0.80 ± 0.05 a | 0.32 ± 0.02 c,d | 0.57 ± 0.03 b | 0.38 ± 0.02 c | 0.27 ± 0.03 d |
| Heptadecanoic acid | 0.02 ± 0.00 | - | - | - | - |
| 9,12-Octadecanoic acid | 0.06 ± 0.01 b | 0.07 ± 0.02 b | 0.11 ± 0.01 a | 0.06 ± 0.01 b | 0.07 ± 0.02 b |
| Oleic acid | 0.15 ± 0.03 a,b | 0.11 ± 0.01 b | 0.20 ± 0.03 a | 0.07 ± 0.01 c | 0.08 ± 0.01 c |
| Octadecanoic acid | 0.25 ± 0.02 a | 0.14 ± 0.03 b | 0.15 ± 0.04 b | 0.19 ± 0.04 a,b | 0.13 ± 0.04 b |
| Tetracosanoic acid | 0.05 ± 0.01 c | 0.05 ± 0.01 c | 0.09 ± 0.01 a | 0.06 ± 0.01 b,c | 0.09 ± 0.02 a,b |
| Alkanes | 0.45 ± 0.01e | 0.70 ± 0.06 d | 1.27 ± 0.04 a | 0.90 ± 0.06 c | 1.07 ± 0.06 b |
| Heptadecane | 0.02 ± 0.00 b | 0.06 ± 0.01 a | 0.03 ± 0.01 b | 0.02 ± 0.01 b,c | 0.01 ± 0.00 c |
| Pentacosane | 0.04 ± 0.01 a | 0.01 ± 0.00 b | 0.06 ± 0.01 a | 0.05 ± 0.01 a | 0.05 ± 0.01 a |
| Hexacosane | 0.03 ± 0.01 a | 0.01 ± 0.00 b | 0.02 ± 0.01 a,b | 0.02 ± 0.01 a,b | 0.02 ± 0.01 a,b |
| Heptacosane | 0.14 ± 0.01 b | 0.03 ± 0.01 b | 0.08 ± 0.02 a | 0.04 ± 0.01 b | 0.04 a ± 0.00 |
| Nonacosane | 0.16 ± 0.02 c | 0.17 ± 0.05 b,c | 0.34 ± 0.02 a | 0.22 ± 0.02 b | 0.30 ± 0.06 a,b |
| Triacontane | 0.02 ± 0.00 c | 0.39 ± 0.09 b | 0.68 ± 0.04 a | 0.50 ± 0.15 a,b | 0.60 ± 0.10 a |
| Hentriacontane | 0.04 ± 0.01 b | 0.03 ± 0.01 b | 0.06 ± 0.01 a | 0.05 ± 0.01 a,b | 0.05 ± 0.01 a,b |
| Alcohols | 0.27 ± 0.04 b | 0.27 ± 0.09 b | 0.53 ± 0.05 a | 0.52 ± 0.10 a | 0.58 ± 0.07 a |
| 13-Docosen-1-ol | 0.25 ± 0.05 a | 0.07 ± 0.01 c | 0.14 ± 0.05 b | 0.17 ± 0.07 a,b | 0.15 ± 0.04 b |
| Tetracosan-1-ol | 0.02 ± 0.01 b | 0.20 ± 0.09 b | 0.39 ± 0.09 a | 0.35 ± 0.10 a,b | 0.43 ± 0.08 a |
| Terpenoids | 2.54 ± 0.11 b | 2.06 ± 0.10 c | 4.35 ± 0.18 a | 2.76 ± 0.15 b | 2.27 ± 0.12 c |
| Farnesol | - | 0.01 ± 0.00 b | 0.03 ± 0.01 a | - | - |
| Squalene | 0.29 ± 0.09 b | 0.12 ± 0.01 c | 0.58 ± 0.10 a | 0.13 ± 0.09 c | - |
| Campesterol | 0.05 ± 0.01 b | 0.05 ± 0.01 b | 0.10 ± 0.02 a | 0.05 ± 0.01 b | 0.06 ± 0.01 b |
| Stigmasterol | 0.04 ± 0.01 a,b | 0.02 ± 0.01 b | 0.05 ± 0.01 a | 0.03 ± 0.01 a,b | 0.04 ± 0.01 a,b |
| Lup-20(29)-en-3-one | 0.30 ± 0.07 c | 0.35 ± 0.04 c | 0.70 ± 0.03 a | 0.50 ± 0.10 b | - |
| Lanosterol | - | 0.04 ± 0.01 b | 0.10 ± 0.04 a | 0.06 ± 0.05 a,b | 0.08 ± 0.02 a |
| Urs-12-en-24-oic acid | - | 0.03 ± 0.01 b | 0.06 ± 0.01 a | 0.04 ± 0.01 a,b | 0.05 ± 0.01 a,b |
| β-Amyrin | 0.36 ± 0.06 c | 0.35 ± 0.05 c | 0.71 ± 0.08 a | 0.48 ± 0.11 b,c | 0.52 ± 0.08 b |
| Friedelin | 1.50 ± 0.12 b | 1.09 ± 0.09 c | 2.02 ± 0.16 a | 1.47 ± 0.14 b | 1.52 ± 0.13 b |
| Compositions | Contents of Cutin Monomer Compositions (μg cm−2) | ||||
|---|---|---|---|---|---|
| Harvest | 10 days | 20 days | 30 days | 40 days | |
| Cinnamic acid | 13.91 ± 0.82 a | 10.29 ± 0.73 b | 8.58 ± 0.32 c | 6.67 ± 0.38 d | 6.26 ± 0.21 d |
| Pentadecanoic acid | 2.21 ± 0.07 a | 1.64 ± 0.12 b | 1.76 ± 0.09 b | 1.08 ± 0.10 c | 1.04 ± 0.05 c |
| Hexadecanoic acid | 4.10 ± 0.13 a | 3.31 ± 0.28 c | 3.71 ± 0.17 b | 3.92 ± 0.09 a,b | 3.95 ± 0.23 a,b |
| Hexadecanedioic acid | 2.20 ± 0.06 b | 1.91 ± 0.05 c | 2.73 ± 0.07 a | 1.81 ± 0.05 c | 1.69 ± 0.02 d |
| Octadecanoic acid | 17.06 ± 0.52 a | 12.95 ± 0.24 c | 14.04 ± 0.34 b | 9.17 ± 0.12 d | 9.33 ± 0.29 d |
| Octadecanedioic acid | 2.89 ± 0.10 a | 1.90 ± 0.08 c | 2.73 ± 0.06 a | 2.09 ± 0.03 b | 1.41 ± 0.01 d |
| Tetracosanoic acid | 6.72 ± 0.32 a | 2.67 ± 0.12 d | 3.90 ± 0.21 b | 2.50 ± 0.13 d | 3.10 ± 0.09 c |
| Octacosanoic acid | 1.31 ± 0.02 a | 1.14 ± 0.07 b | 0.78 ± 0.03 c | - | - |
| Phenol | 2.10 ± 0.16 a | 2.29 ± 0.13 a | 0.78 ± 0.06 c | 0.56 ± 0.02 c | 1.41 ± 0.05 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, S.; Zhang, J.; Yang, L.; Wang, X.; Fu, F.; Wang, R.; Zhang, Q.; Shan, Y. Changes in Cuticle Components and Morphology of ‘Satsuma’ Mandarin (Citrus unshiu) during Ambient Storage and Their Potential Role on Penicillium digitatum Infection. Molecules 2020, 25, 412. https://doi.org/10.3390/molecules25020412
Ding S, Zhang J, Yang L, Wang X, Fu F, Wang R, Zhang Q, Shan Y. Changes in Cuticle Components and Morphology of ‘Satsuma’ Mandarin (Citrus unshiu) during Ambient Storage and Their Potential Role on Penicillium digitatum Infection. Molecules. 2020; 25(2):412. https://doi.org/10.3390/molecules25020412
Chicago/Turabian StyleDing, Shenghua, Jing Zhang, Lvzhu Yang, Xinyu Wang, Fuhua Fu, Rongrong Wang, Qun Zhang, and Yang Shan. 2020. "Changes in Cuticle Components and Morphology of ‘Satsuma’ Mandarin (Citrus unshiu) during Ambient Storage and Their Potential Role on Penicillium digitatum Infection" Molecules 25, no. 2: 412. https://doi.org/10.3390/molecules25020412