
IgY Targeting Bacterial Quorum-Sensing Molecules in Implant-Associated Infections


Abstract
:1. Introduction
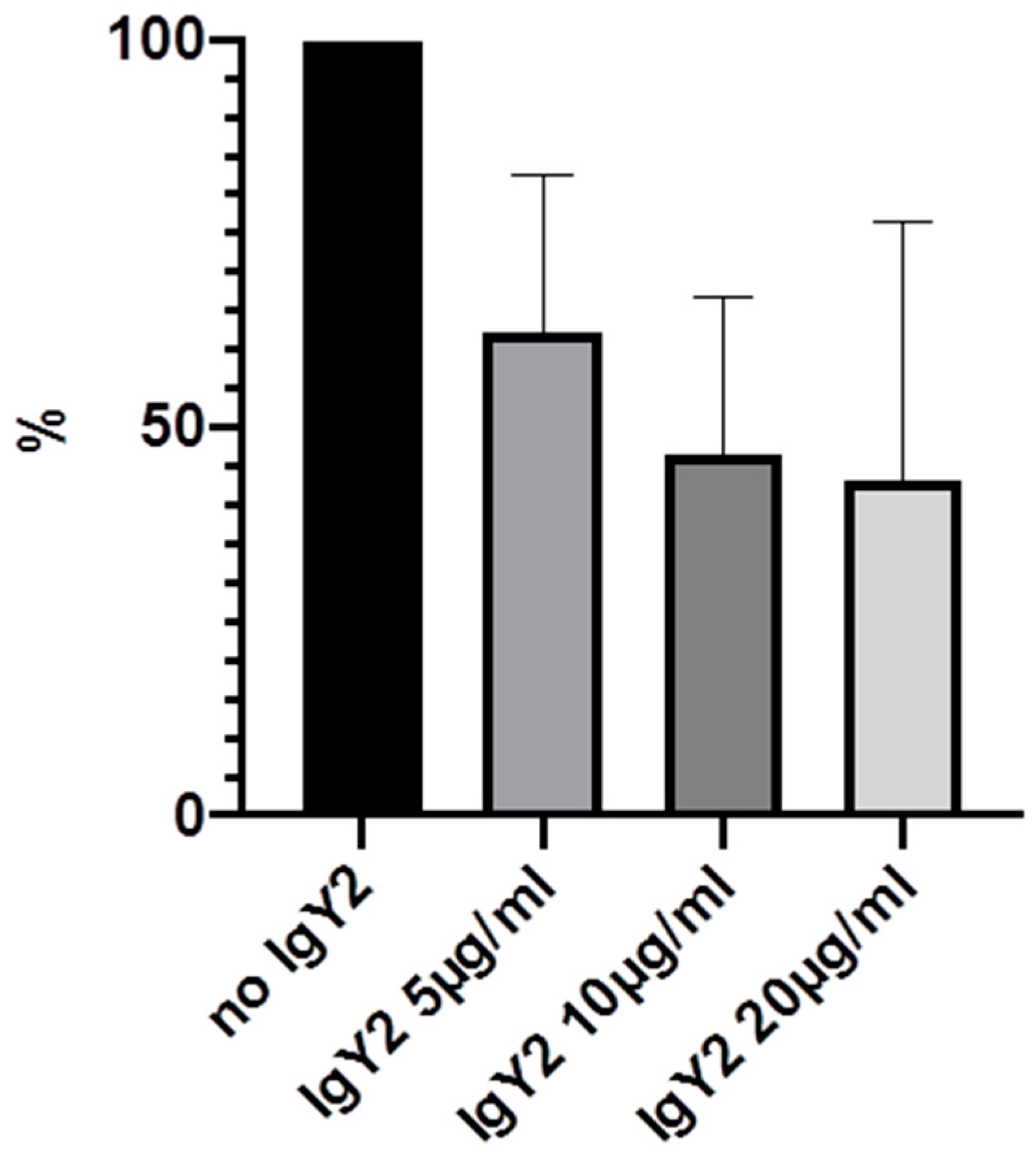
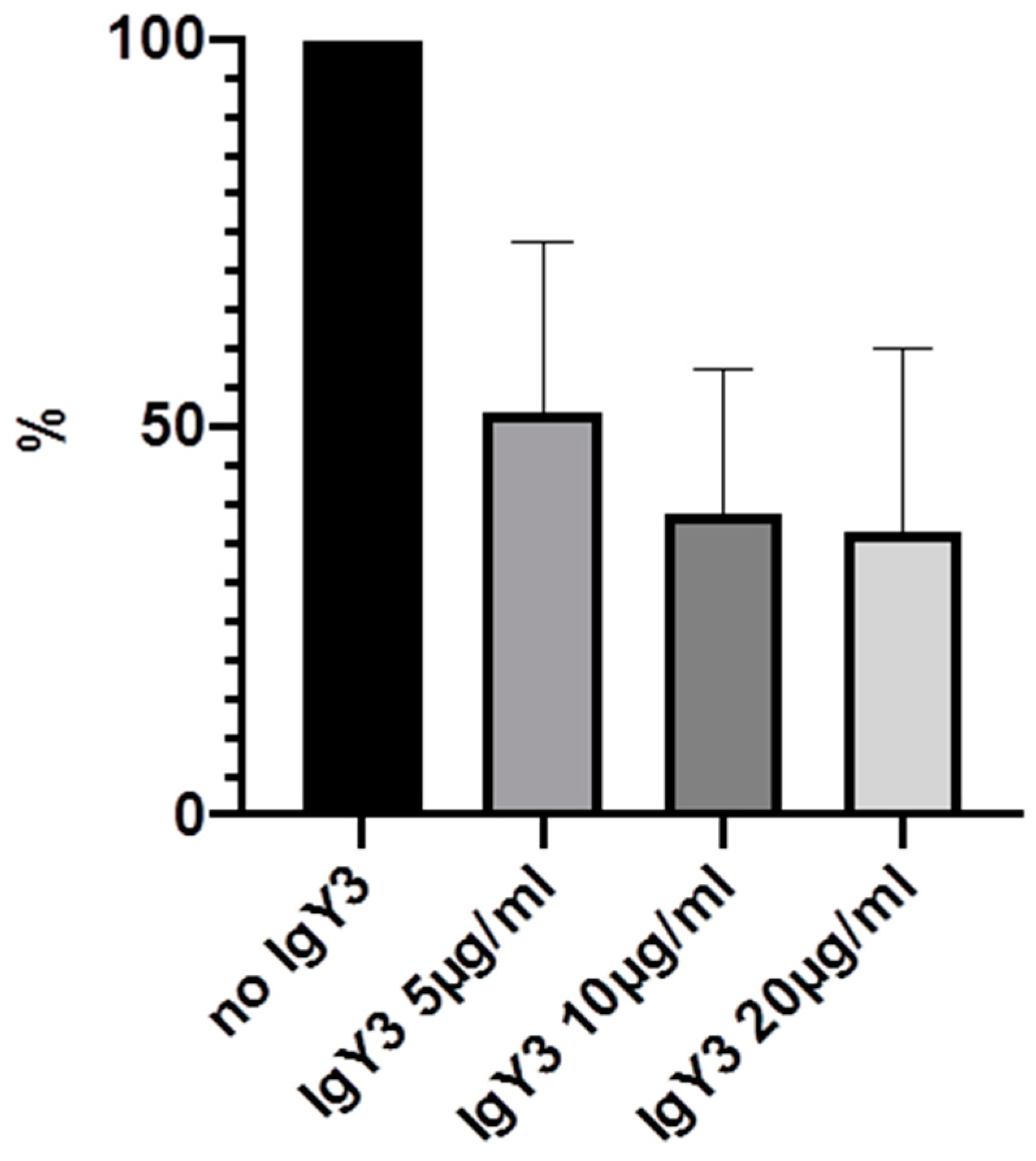
2. Results
3. Discussion
4. Materials and Methods
4.1. Generation of IgY
4.2. Formation of Biofilms and Effect of IgYs
4.3. Culture of Osteoblasts
4.4. Bicompatibility of IgY
4.5. ELISA
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Haenle, M.; Skripitz, C.; Mittelmeier, W.; Skripitz, R. Economic impact of infected total hip arthroplasty in the German diagnosis-related groups system. Orthopade 2012, 41, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Sambri, A.; Maso, A.; Storni, E.; Megaloikonomos, P.D.; Igoumenou, V.G.; Errani, C.; Mavrogenis, A.F.; Bianchi, G. Sonication Improves the Diagnosis of Megaprosthetic Infections. Orthopedics 2019, 42, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Sambri, A.; Maso, A.; Storni, E.; Donati, M.E.; Pederzoli, A.; Dallari, D.; Bianchi, G.; Donati, D.M. Is sonication of antibiotic-loaded cement spacers useful in two-stage revision of prosthetic joint infection? J. Microbiol. Methods 2019, 156, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Sambri, A.; Cadossi, M.; Giannini, S.; Pignatti, G.; Marcacci, M.; Neri, M.P.; Maso, A.; Storni, E.; Gamberini, S.; Naldi, S.; et al. Is Treatment with Dithiothreitol More Effective Than Sonication for the Diagnosis of Prosthetic Joint Infection? Clin. Orthop. Relat. Res. 2018, 476, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Flemming, H.C. Biofouling in water systems—Cases, causes and countermeasures. Appl. Microbiol. Biotechnol. 2002, 59, 629–640. [Google Scholar] [CrossRef]
- Dunne, W.M., Jr. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.R. Microcolony and biofilm formation as a survival strategy for bacteria. J. Theor. Biol. 2008, 251, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Fux, C.A.; Costerton, J.W.; Stewart, P.S.; Stoodley, P. Survival strategies of infectious biofilms. Trends Microbiol. 2005, 13, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Montanaro, L.; Arciola, C.R. Biofilm in implant infections: Its production and regulation. Int. J. Artif. Organs 2005, 28, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Karatan, E.; Watnick, P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310–347. [Google Scholar] [CrossRef] [Green Version]
- Henke, J.M.; Bassler, B.L. Bacterial social engagements. Trends Cell Biol. 2004, 14, 648–656. [Google Scholar] [CrossRef]
- Bassler, B.L. Small talk. Cell-to-cell communication in bacteria. Cell 2002, 109, 421–424. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, C.; Greenberg, E.P. Listening in on bacteria: Acyl-homoserine lactone signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 685–695. [Google Scholar] [CrossRef]
- Gaida, M.M.; Dapunt, U.; Hansch, G.M. Sensing developing biofilms: The bitter receptor T2R38 on myeloid cells. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [Green Version]
- Dapunt, U.; Radzuweit-Mihaljevic, S.; Lehner, B.; Haensch, G.M.; Ewerbeck, V. Bacterial Infection and Implant Loosening in Hip and Knee Arthroplasty: Evaluation of 209 Cases. Materials 2016, 9, 871. [Google Scholar] [CrossRef] [Green Version]
- Kong, K.F.; Vuong, C.; Otto, M. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol. 2006, 296, 133–139. [Google Scholar] [CrossRef]
- Biswas, R.; Voggu, L.; Simon, U.K.; Hentschel, P.; Thumm, G.; Gotz, F. Activity of the major staphylococcal autolysin Atl. FEMS Microbiol. Lett. 2006, 259, 260–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speziale, P.; Pietrocola, G.; Rindi, S.; Provenzano, M.; Provenza, G.; Di Poto, A.; Visai, L.; Arciola, C.R. Structural and functional role of Staphylococcus aureus surface omponents recognizing adhesive matrix molecules of the host. Future Microbiol. 2009, 4, 1337–1352. [Google Scholar] [CrossRef] [PubMed]
- O’Gara, J.P. Ica and beyond: Biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdbart, J.O.; Allignet, J.; Tung, H.S.; Ryden, C.; El Solh, N. Screening for Staphylococcus epidermidis markers discriminating between skin-flora strains and those responsible for infections of joint prostheses. J. Infect. Dis. 2000, 182, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Rupp, M.E.; Ulphani, J.S.; Fey, P.D.; Bartscht, K.; Mack, D. Characterization of the importance of polysaccharide intercellular adhesin/hemagglutinin of Staphylococcus epidermidis in the pathogenesis of biomaterial-based infection in a mouse foreign body infection model. Infect. Immun. 1999, 67, 2627–2632. [Google Scholar] [CrossRef] [Green Version]
- Baranova, I.N.; Vishnyakova, T.G.; Bocharov, A.V.; Leelahavanichkul, A.; Kurlander, R.; Chen, Z.; Souza, A.C.; Yuen, P.S.; Star, R.A.; Csako, G.; et al. Class B scavenger receptor types I and II and CD36 mediate bacterial recognition and proinflammatory signaling induced by Escherichia coli, lipopolysaccharide, and cytosolic chaperonin 60. J. Immunol. 2012, 188, 1371–1380. [Google Scholar] [CrossRef] [Green Version]
- Young, D.B.; Ivanyi, J.; Cox, J.H.; Lamb, J.R. The 65kDa antigen of mycobacteria-a common bacterial protein? Immunol. Today 1987, 8, 215–219. [Google Scholar] [CrossRef]
- Dapunt, U.; Gaida, M.M.; Meyle, E.; Prior, B.; Hansch, G.M. Activation of phagocytic cells by Staphylococcus epidermidis biofilms: Effects of extracellular matrix proteins and the bacterial stress protein GroEL on netosis and MRP-14 release. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [Green Version]
- Maurer, S.F.P.; Meyle, E.; Prior, B.; Hänsch, G.M.; Dapunt, U. Activation of Neutrophils by the Extracellular Polymeric Substance of S.Epidermidis Biofilms is Mediated by The Bacterial Heat Shock Protein Groel. J. Biotechnol. Biomater. 2015, 5, 176. [Google Scholar]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Zimmerli, W.; Sendi, P. Pathogenesis of implant-associated infection: The role of the host. Semin. Immunopathol. 2011, 33, 295–306. [Google Scholar] [CrossRef]
- Zimmerli, W.; Trampuz, A.; Ochsner, P.E. Prosthetic-joint infections. N. Engl. J. Med. 2004, 351, 1645–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoodley, P.; Ehrlich, G.D.; Sedghizadeh, P.P.; Hall-Stoodley, L.; Baratz, M.E.; Altman, D.T.; Sotereanos, N.G.; Costerton, J.W.; Demeo, P. Orthopaedic biofilm infections. Curr. Orthop. Pract. 2011, 22, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Otto, M. Quorum-sensing control in Staphylococci—A target for antimicrobial drug therapy? FEMS Microbiol. Lett. 2004, 241, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef] [PubMed]
- Rampioni, G.; Leoni, L.; Williams, P. The art of antibacterial warfare: Deception through interference with quorum sensing-mediated communication. Bioorg. Chem. 2014, 55, 60–68. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, F.; Baldelli, V.; Halliday, N.; Pantalone, P.; Polticelli, F.; Fiscarelli, E.; Williams, P.; Visca, P.; Leoni, L.; Rampioni, G. Identification of FDA-Approved Drugs as Antivirulence Agents Targeting the pqs Quorum-Sensing System of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e01296-e18. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, R.R.; Jin, L.Z.; Kim, J.W.; Fang, L.; Frohlich, A.A.; Baidoo, S.K. Passive protective effect of egg-yolk antibodies against enterotoxigenic Escherichia coli K88+ infection in neonatal and early-weaned piglets. FEMS Immunol. Med. Microbiol. 1999, 23, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.N.; Sunwoo, H.H.; Menninen, K.; Sim, J.S. In vitro studies of chicken egg yolk antibody (IgY) against Salmonella enteritidis and Salmonella typhimurium. Poult. Sci. 2002, 81, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Tsubokura, K.; Berndtson, E.; Bogstedt, A.; Kaijser, B.; Kim, M.; Ozeki, M.; Hammarstrom, L. Oral administration of antibodies as prophylaxis and therapy in Campylobacter jejuni-infected chickens. Clin. Exp. Immunol. 1997, 108, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.A.; Casswall, T.H.; Juneja, L.R.; Hoq, E.; Hossain, I.; Fuchs, G.J.; Hammarstrom, L. Randomized, placebo-controlled, clinical trial of hyperimmunized chicken egg yolk immunoglobulin in children with rotavirus diarrhea. J. Pediatr. Gastroenterol. Nutr. 2001, 32, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Warr, G.W.; Magor, K.E.; Higgins, D.A. IgY: Clues to the origins of modern antibodies. Immunol. Today 1995, 16, 392–398. [Google Scholar] [CrossRef]
- Carlander, D.; Kollberg, H.; Wejaker, P.E.; Larsson, A. Peroral immunotherapy with yolk antibodies for the prevention and treatment of enteric infections. Immunol. Res. 2000, 21, 1–6. [Google Scholar] [CrossRef]
- Tini, M.; Jewell, U.R.; Camenisch, G.; Chilov, D.; Gassmann, M. Generation and application of chicken egg-yolk antibodies. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 131, 569–574. [Google Scholar] [CrossRef]
- Gassmann, M.; Thommes, P.; Weiser, T.; Hubscher, U. Efficient production of chicken egg yolk antibodies against a conserved mammalian protein. FASEB J. 1990, 4, 2528–2532. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.C.; Guo, J.; Chen, S.H.; Tian, L.G.; Steinmann, P.; Chen, M.X.; Li, H.; Ai, L.; Chen, J.X. Chicken egg yolk antibodies (IgY) for detecting circulating antigens of Schistosoma japonicum. Parasitol. Int. 2012, 61, 385–390. [Google Scholar] [CrossRef]
- Rahman, S.; Van Nguyen, S.; Icatlo, F.C., Jr.; Umeda, K.; Kodama, Y. Oral passive IgY-based immunotherapeutics: A novel solution for prevention and treatment of alimentary tract diseases. Hum. Vaccines Immunother. 2013, 9, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- LeClaire, R.D.; Hunt, R.E.; Bavari, S. Protection against bacterial superantigen staphylococcal enterotoxin B by passive vaccination. Infect. Immun. 2002, 70, 2278–2281. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, G.; Akova, M.; Rello, J.; Poulakou, G. Understanding resistance in Pseudomonas. Intensiv. Care Med. 2020, 46, 350–352. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No IgY | 5 µg/mL (SD) | 10 µg/mL (SD) | 20 µg/mL (SD) | |
|---|---|---|---|---|
| IgY1 (AtlE) | 100% | 72% (15.4) | 50.2% (21.7) | 44.4% (22.7) |
| IgY2 (GroEL) | 100% | 62.3% (20.3) | 46.8% (19.9) | 43.3% (33.3) |
| IgY3 (PIA) | 100% | 52% (21.8) | 38.8% (18.5) | 36.6% (23.4) |
| IgYcontrol | 100% | 58.4% (15.8) | 49.6% (16.2) | 31.8% (14.4) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dapunt, U.; Prior, B.; Oelkrug, C.; Kretzer, J.P. IgY Targeting Bacterial Quorum-Sensing Molecules in Implant-Associated Infections. Molecules 2020, 25, 4027. https://doi.org/10.3390/molecules25174027
Dapunt U, Prior B, Oelkrug C, Kretzer JP. IgY Targeting Bacterial Quorum-Sensing Molecules in Implant-Associated Infections. Molecules. 2020; 25(17):4027. https://doi.org/10.3390/molecules25174027
Chicago/Turabian StyleDapunt, Ulrike, Birgit Prior, Christopher Oelkrug, and Jan Philippe Kretzer. 2020. "IgY Targeting Bacterial Quorum-Sensing Molecules in Implant-Associated Infections" Molecules 25, no. 17: 4027. https://doi.org/10.3390/molecules25174027