
EPA/DHA Concentrate by Urea Complexation Decreases Hyperinsulinemia and Increases Plin5 in the Liver of Mice Fed a High-Fat Diet

 , and
, and
Abstract
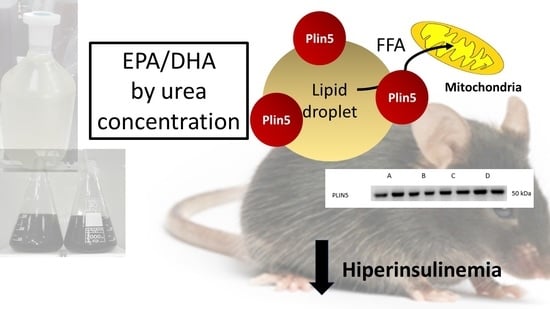
:
1. Introduction
2. Results
2.1. Fatty Acid Composition and Quantification of Refined Salmon Oil
2.2. Optimization of the Process for Obtaining CPUFA by Urea Complexation
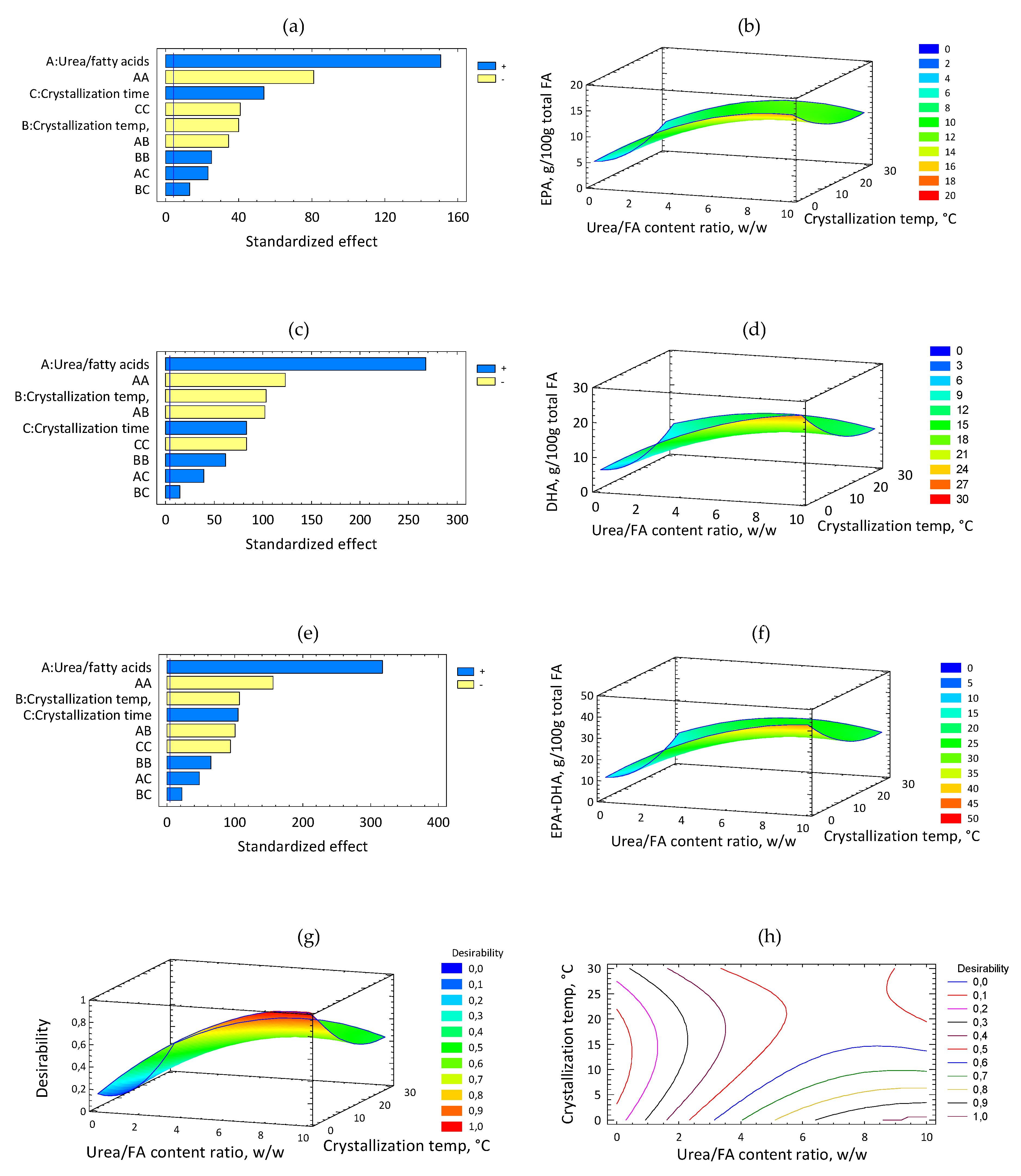
2.3. Effect of Urea Concentration Process Variables on EPA, DHA and EPA + DHA of CPUFA: Analysis by RSM
2.4. Optimization of the Process for Obtaining CPUFA and CDSO by Urea Complexation by RSM
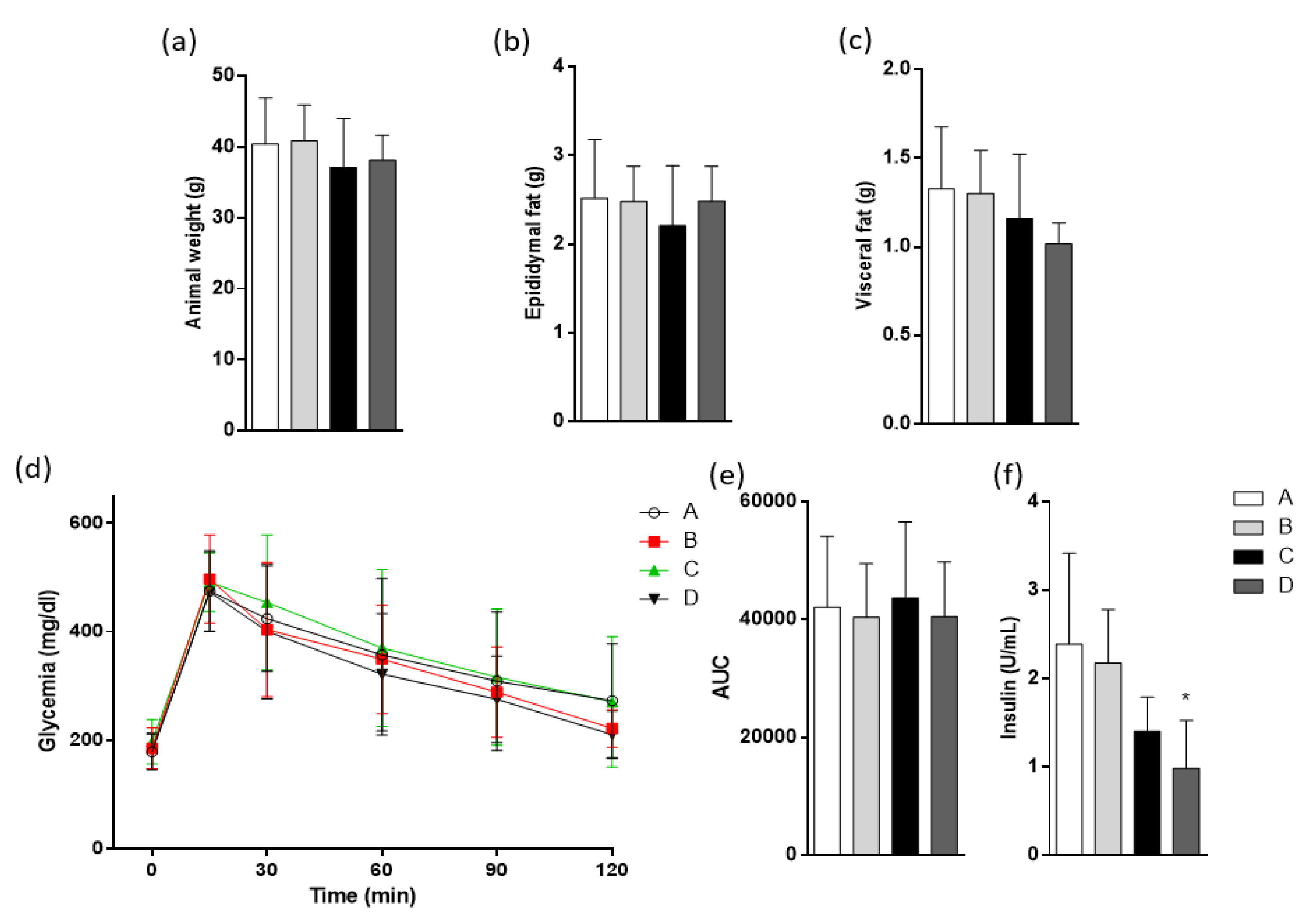
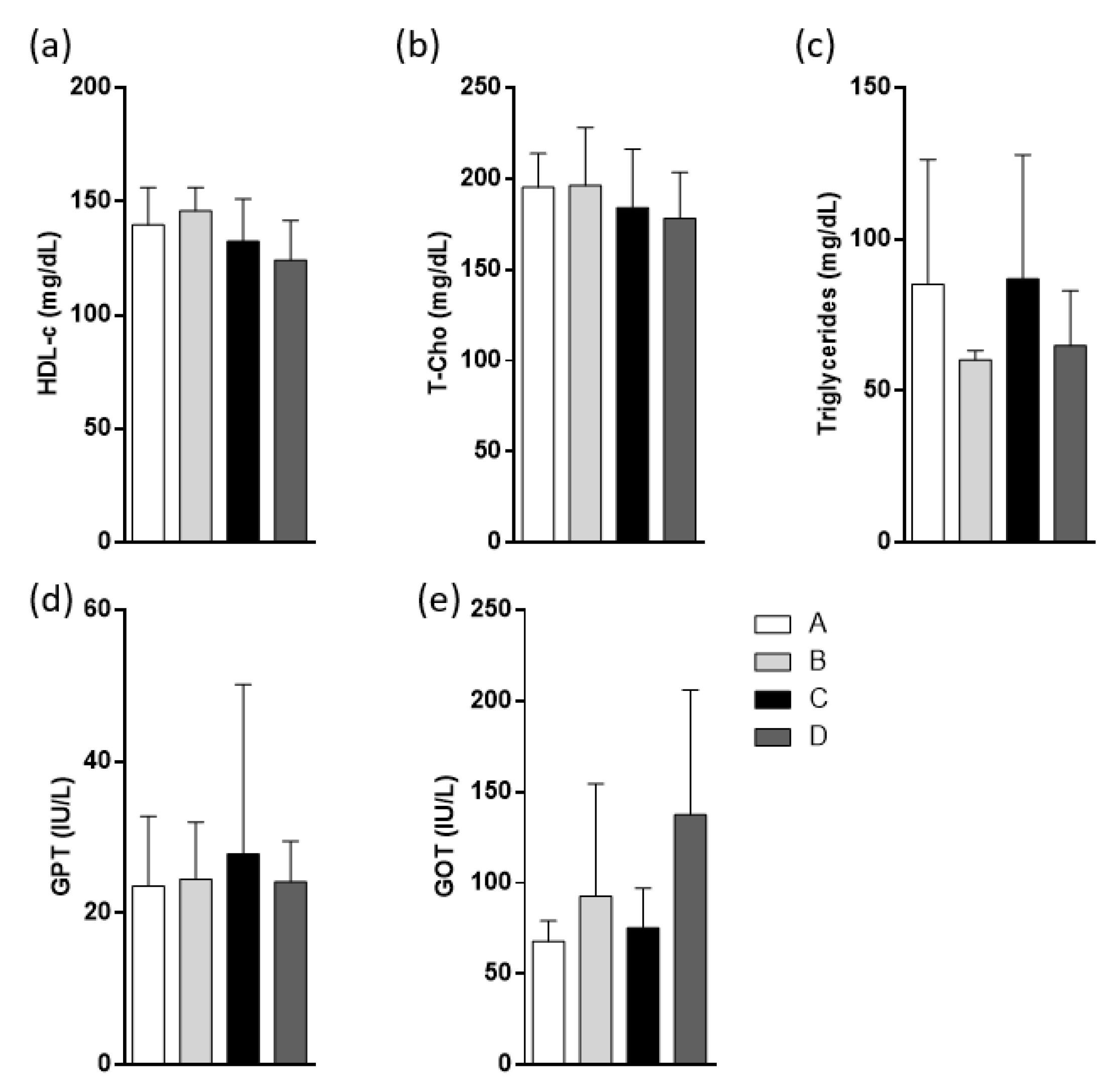
2.5. Weights and Metabolic Parameters in High-Fat Diet (HFD)-Fed Mice
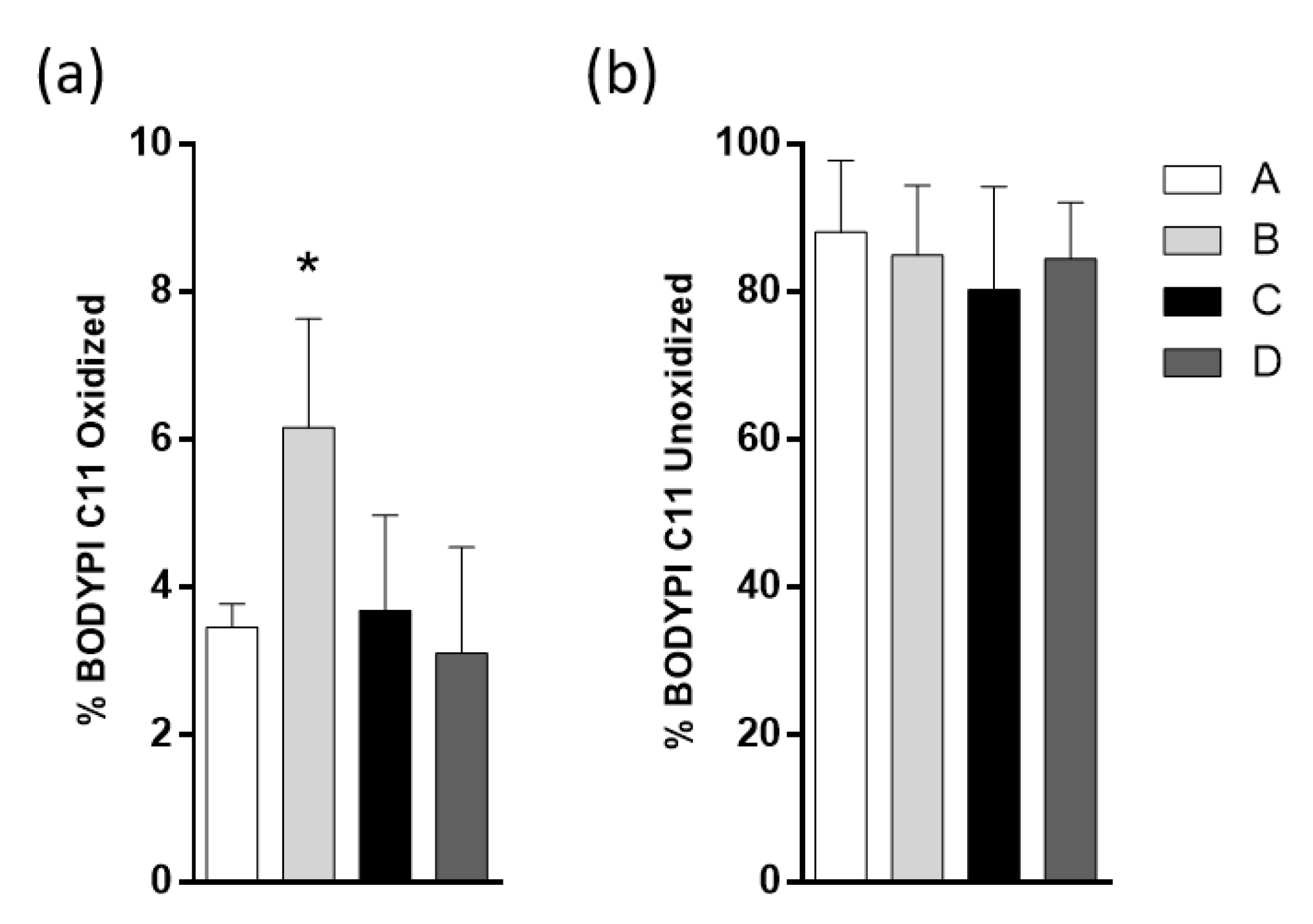
2.6. Fish Oil Products do Not Affect the Lipid Droplets (LD) Oxidation Induced by HFD
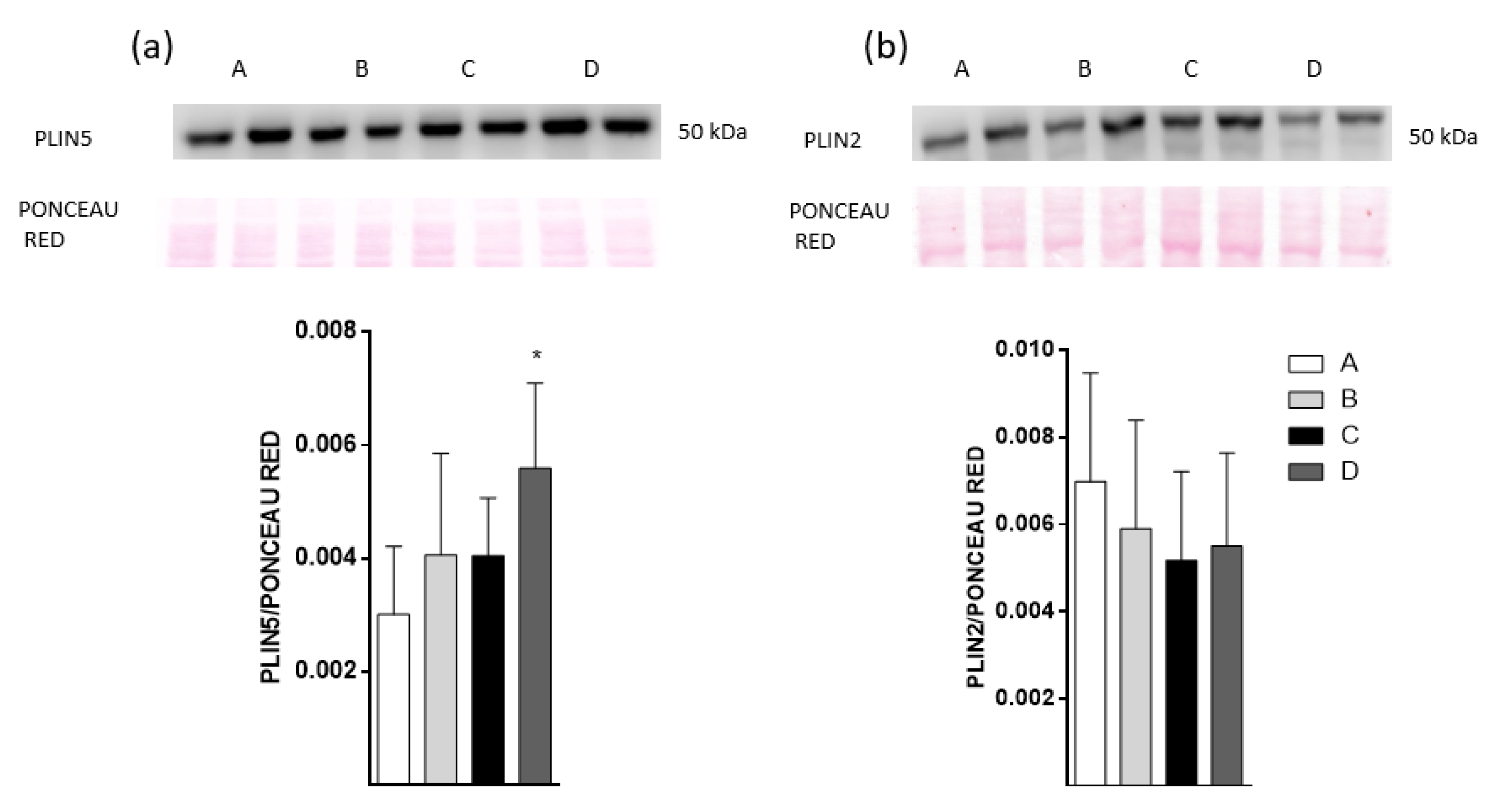
2.7. Plin5 Protein Level Increase with Dietary Fish Oil Products Intake in HFD-Fed Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. DSO Characterization
4.3. Fatty Acid Composition and EPA/DHA Quantification
4.4. EPA and DHA Concentrate from DSO by Urea Complexation Process (CPUFA)
4.5. CPUFA Concentrate
4.6. Optimization of CPUFA by Response Surface Methodology (RSM)
4.7. Animals
4.8. Biological Samples
4.9. Measurements of Serum Parameters
4.10. Glucose Tolerance Test
4.11. Hepatocytes Isolation and Flow Cytometry
4.12. Western Blot Analysis of Perilipins
4.13. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Apovian, C. Obesity: Definition, Comorbidities, Causes, and Burden. Am. J. Manag. Care 2016, 22, s176–s185. [Google Scholar] [PubMed]
- De Luca, C.; Olefsky, J.M. Inflammation and Insulin Resistance. FEBS Lett. 2007, 582, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose Tissue in Obesity-Related Inflammation and Insulin Resistance: Cells, Cytokines, and Chemokines. ISRN Inflamm. 2013, 2013, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, E.D.; Lee, J.; Hansen, L.; Yuan, M.; Shoelson, S.E. Insulin Resistance Due to Phosphorylation of Insulin Receptor Substrate-1 at Serine 302. J. Boil. Chem. 2004, 279, 35298–35305. [Google Scholar] [CrossRef] [Green Version]
- Anusree, S.; Sindhu, G.; Rani, M.P.; Raghu, K. Insulin Resistance in 3T3-L1 Adipocytes by TNF-α is Improved by Punicic Acid Through Upregulation of Insulin Signalling Pathway and Endocrine Function, and Downregulation of Proinflammatory Cytokines. Biochimie 2018, 146, 79–86. [Google Scholar] [CrossRef]
- Smith, U.; Kahn, B.B. Adipose Tissue Regulates Insulin Sensitivity: Role of Adipogenesis, de Novolipogenesis and Novel Lipids. J. Intern. Med. 2016, 280, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Hodson, L.; Bhatia, L.; Scorletti, E.; E Smith, D.; Jackson, N.C.; Shojaee-Moradie, F.; Umpleby, M.; Calder, P.C.; Byrne, C.D. Docosahexaenoic Acid Enrichment in NAFLD is Associated with Improvements in Hepatic Metabolism and Hepatic Insulin Sensitivity: A Pilot Study. Eur. J. Clin. Nutr. 2017, 71, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Green, C.J.; Pramfalk, C.; A Charlton, C.; Gunn, P.J.; Cornfield, T.; Pavlides, M.; Karpe, F.; Hodson, L. Hepatic de Novo Lipogenesis is Suppressed and Fat Oxidation is Increased by Omega-3 fatty Acids at the Expense of Glucose Metabolism. BMJ Open Diabetes Res. Care 2020, 8, e000871. [Google Scholar] [CrossRef] [Green Version]
- Lalia, A.Z.; Johnson, M.L.; Jensen, M.D.; Hames, K.C.; Port, J.D.; Lanza, I.R. Effects of Dietary n-3 Fatty Acids on Hepatic and Peripheral Insulin Sensitivity in Insulin-Resistant Humans. Diabetes Care 2015, 38, 1228–1237. [Google Scholar] [CrossRef] [Green Version]
- Gnoni, A.; Giudetti, A.M. Dietary Long-Chain Unsaturated Fatty Acids Acutely and Differently Reduce the Activities of Lipogenic Enzymes and of Citrate Carrier in Rat Liver. J. Physiol. Biochem. 2016, 72, 485–494. [Google Scholar] [CrossRef]
- Valenzuela, R.; Espinosa, A.; Llanos, P.; Hernandez-Rodas, M.C.; Barrera, C.; Vergara, D.; Romero, N.; Pérez, F.; Ruz, M.; Videla, L. Anti-Steatotic Effects of an n-3 LCPUFA and Extra Virgin Olive Oil Mixture in the Liver of Mice Subjected to High-Fat Diet. Food Funct. 2016, 7, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Soto-Alarcón, S.A.; Ortiz, M.; Orellana, P.; Echeverría, F.; Bustamante, A.; Espinosa, A.; Illesca, P.; Gonzalez-Mañán, D.; Valenzuela, R.; Videla, L.A. Docosahexaenoic Acid and Hydroxytyrosol Co-Administration Fully Prevents Liver Steatosis and Related Parameters in Mice Subjected to High-Fat Diet: A Molecular Approach. BioFactors 2019, 45, 930–943. [Google Scholar] [CrossRef] [PubMed]
- Keenan, S.N.; Meex, R.C.; Lo, J.C.Y.; Ryan, A.; Nie, S.; Montgomery, M.K.; Watt, M.J. Perilipin 5 Deletion in Hepatocytes Remodels Lipid Metabolism and Causes Hepatic Insulin Resistance in Mice. Diabetes 2019, 68, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Morales, P.E.; Bucarey, J.L.; Espinosa, A. Muscle Lipid Metabolism: Role of Lipid Droplets and Perilipins. J. Diabetes Res. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Ritter, J.C.S.; Budge, S.M.; Jovica, F.; Reid, A.-J.M. Oxidation Rates of Triacylglycerol and Ethyl Ester Fish Oils. J. Am. Oil Chem. Soc. 2015, 92, 561–569. [Google Scholar] [CrossRef]
- Berríos, M.M.; Rodriguez, A.; Rivera, M.; Pando, M.E.; Valenzuela, M.A.; Aubourg, S.P. Optimisation of Rancidity Stability in Long-Chain PUFA Concentrates Obtained from a Rainbow Trout (Oncorhynchus Mykiss) by-Product. Int. J. Food Sci. Technol. 2017, 52, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Ma, E.P.; Bravo, B.; Berrios, M.; Galdames, A.; Rojas, C.; Romero, N.; Camilo, C.; Encina, C.; Rivera, M.; Rodríguez, A.; et al. Concentrating n-3 Fatty Acids from Crude and Refined Commercial Salmon Oil. Czech J. Food Sci. 2014, 32, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Pando, M.E.; Rodríguez, A.; Galdames, A.; Berríos, M.M.; Rivera, M.; Romero, N.; Valenzuela, M.A.; Ortiz, J.; Aubourg, S.P. Maximization of the Docosahexaenoic and Eicosapentaenoic Acids Content in Concentrates Obtained from a by-Product of Rainbow Trout (Oncorhynchus mykiss) processing. Eur. Food Res. Technol. 2017, 244, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Méndez, C.; Masson, L.; Jiménez, P. Estabilización de Aceite de Pescado por Medio de Antioxidantes Naturales. Aceites Grasas 2010, 80, 492–500. [Google Scholar]
- Global Organization for EPA and DHA Omega-3. Voluntary Monograph. 2016. Available online: https://www.goedomega3.com/storage/app/media/annual%20reports/GOED%20Annual%20Report%20-%202016.pdf (accessed on 1 February 2020).
- Dovale-Rosabal, G.; Rodríguez, A.; Contreras, E.; Ortiz, J.; Muñoz, M.; Trigo, M.; Aubourg, S.P.; Espinosa, A. Concentration of EPA and DHA from Refined Salmon Oil by Optimizing the Urea-Fatty Acid Adduction Reaction Conditions Using Response Surface Methodology. Molecules 2019, 24, 1642. [Google Scholar] [CrossRef] [Green Version]
- Thammapat, P.; Siriamornpun, S.; Raviyan, P. Concentration of Eicosapentaenoic Acid (EPA) and Docosahexaenoic acid (DHA) of Asian Catfish Oil by Urea Complexation: Optimization of Reaction Conditions. Songklanakarin J. Sci. Technol. (SJST) 2016, 38. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Belarbi, E.-H. Purification Process for Cod Liver Oil Polyunsaturated Fatty Acids. J. Am. Oil Chem. Soc. 2001, 78, 477–484. [Google Scholar] [CrossRef]
- Zuta, C.P.; Simpson, B.; Chan, H.M.; Phillips, L. Concentrating PUFA from Mackerel Processing Waste. J. Am. Oil Chem. Soc. 2003, 80, 933–936. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, C.; Hong, P.; Ji, H. Concentration of Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) of Tuna Oil by urea Complexation: Optimization of Process Parameters. J. Food Eng. 2006, 73, 203–209. [Google Scholar] [CrossRef]
- Vázquez, L.; Prados, I.M.; Reglero, G.; Torres, C. Identification and Quantification of ethyl Carbamate Occurring in Urea Complexation Processes Commonly Utilized for Polyunsaturated Fatty Acid Concentration. Food Chem. 2017, 229, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Magallanes, L.; Tarditto, L.V.; Grosso, N.R.; Pramparo, M.C.; Gayol, M.F. Highly Concentrated Omega-3 Fatty Acid Ethyl Esters by Urea Complexation and Molecular Distillation. J. Sci. Food Agric. 2018, 99, 877–884. [Google Scholar] [CrossRef]
- Haagsma, N.; Van Gent, C.M.; Luten, J.B.; De Jong, R.W.; Van Doorn, E. Preparation of an ω3 Fatty Acid Concentrate from Cod Liver Oil. J. Am. Oil Chem. Soc. 1982, 59, 117–118. [Google Scholar] [CrossRef]
- Ratnayake, W.M.N.; Olsson, B.; Matthews, D.; Ackman, R.G. Preparation of Omega-3 PUFA Concentrates from Fish Oils via Urea Complexation. Fette Seifen Anstrichm. 1988, 90, 381–386. [Google Scholar] [CrossRef]
- Wanasundara, U.N.; Shahidi, F. Concentration of Omega 3-Polyunsaturated Fatty Acids of Seal Blubber Oil by Urea Complexation: Optimization of Reaction Conditions. Food Chem. 1999, 65, 41–49. [Google Scholar] [CrossRef]
- Jacobsen, C. Food Enrichment with Omega-3 Fatty Acids; Publishing Series in Food Science, Technology and Nutrition; Woodhead Publ: Oxford, UK, 2013; ISBN 978-0-85709-886-3. [Google Scholar]
- Horn, S.S.; Sonesson, A.K.; Krasnov, A.; Moghadam, H.; Hillestad, B.; Meuwissen, T.H.E.; Ruyter, B. Individual Differences in EPA and DHA Content of Atlantic Salmon are Associated with Gene Expression of key Metabolic Processes. Sci. Rep. 2019, 9, 3889. [Google Scholar] [CrossRef]
- Liu, B.; Fang, Y.; Yi, R.; Zhao, X. Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice. Foods 2019, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- La Fuente, F.P.-D.; Nocetti, D.; Sacristán, C.; Ruiz, P.; Guerrero, J.; Jorquera, G.; Uribe, E.; Bucarey, J.L.; Espinosa, A.; Puente, L. Physalis peruviana L. Pulp Prevents Liver Inflammation and Insulin Resistance in Skeletal Muscles of Diet-Induced Obese Mice. Nutrients 2020, 12, 700. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, A.; Campos, C.; Díaz-Vegas, A.; Galgani, J.E.; Juretić, N.; Osorio-Fuentealba, C.; Bucarey, J.L.; Tapia, G.; Valenzuela, R.; Contreras-Ferrat, A.; et al. Insulin-Dependent H2O2 Production Is Higher in Muscle Fibers of Mice Fed with a High-Fat Diet. Int. J. Mol. Sci. 2013, 14, 15740–15754. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.A.; Minze, L.J.; Reyes, M.; Ren, Y.; Wang, X.; Brunner, G.; Ghosn, M.; Cordero-Reyes, A.M.; Ding, K.; Praticò, M.; et al. High-Fat Feeding-Induced Hyperinsulinemia Increases Cardiac Glucose Uptake and Mitochondrial Function Despite Peripheral Insulin Resistance. Endocrinology 2013, 154, 2650–2662. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Corral, C.; Alderete, T.L.; Richey, J.; Sequeira, P.; Goran, M.I.; Weigensberg, M.J. Fasting, Post-OGTT Challenge, and Nocturnal Free Fatty Acids in Prediabetic Versus Normal Glucose Tolerant Overweight and Obese Latino Adolescents. Acta Diabetol. 2014, 52, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Tsuboyama-Kasaoka, N.; Nakatani, T.; Ishii, M.; Tsutsumi, S.; Aburatani, H.; Ezaki, O. Fish Oil Feeding Alters Liver Gene Expressions to Defend Against PPARα Activation and ROS Production. Am. J. Physiol. Liver Physiol. 2002, 282, G338–G348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polus, A.; Zapala, B.; Raźny, U.; Gielicz, A.; Kiec-Wilk, B.; Malczewska-Malec, M.; Sanak, M.; Childs, C.E.; Calder, P.C.; Dembinska-Kiec, A. Omega-3 Fatty Acid Supplementation Influences the Whole Blood Transcriptome in Women with Obesity, Associated with Pro-Resolving Lipid Mediator Production. Biochim. Biophys. Acta (BBA)-Mol. Cell Boil. Lipids 2016, 1861, 1746–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhard, A.; Berntssen, M.H.; Lundebye, A.-K.; Alvheim, A.R.; Myrmel, L.S.; Fjære, E.; Torstensen, B.E.; Kristiansen, K.; Madsen, L.; Brattelid, T.; et al. Marine Fatty Acids Aggravate Hepatotoxicity of α-HBCD in Juvenile Female BALB/c mice. Food Chem. Toxicol. 2016, 97, 411–423. [Google Scholar] [CrossRef]
- De Meijer, V.E.; Kalish, B.T.; Meisel, J.A.; Le, H.D.; Puder, M. Dietary Fish Oil Aggravates Paracetamol-Induced Liver Injury in Mice. J. Parenter. Enter. Nutr. 2012, 37, 268–273. [Google Scholar] [CrossRef]
- Trevino, M.B.; Mazur-Hart, D.; Machida, Y.; King, T.; Nadler, J.; Galkina, E.V.; Poddar, A.; Dutta, S.; Imai, Y. Liver Perilipin 5 Expression Worsens Hepatosteatosis But Not Insulin Resistance in High Fat-Fed Mice. Mol. Endocrinol. 2015, 29, 1414–1425. [Google Scholar] [CrossRef] [PubMed]
- Hill, W.J.; Hunter, W.G. A Review of Response Surface Methodology: A Literature Survey. Technometrics 1966, 8, 571. [Google Scholar] [CrossRef]
- Gunst, R.F.; Myers, R.H.; Montgomery, D.C. Response Surface Methodology: Process and Product Optimization Using Designed Experiments. Technometrics 1996, 38, 285. [Google Scholar] [CrossRef]
- European Pharmacopoeia. Composition of Fatty Acids in Oils Rich in Omega-3-Acids, 5th ed.; Main Volume 5.0; 01/2005:20429; Council of Europe: Strasbourg, France, 2005. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systematic Name | Abbreviated Name | DSO (g/100 Total FA) |
|---|---|---|
| Myristic acid | C14:0 | 2.17 ± 0.07 |
| Palmitic acid | C16:0 | 11.40 ± 0.23 |
| Palmitoleic acid | C16:1 n-7 | 3.29 ± 0.06 |
| Hexadecadienoic acid | C16:2 n-4 | 0.29 ± 0.01 |
| Hexadecatrienoic acid | C16:3 n-4 | 0.26 ± 0.01 |
| Hexadecatetraenoic acid | C16:4 n-1 | 0.42 ± 0.01 |
| Heptadecanoic | C17:0 | 0.03 ± 0.01 |
| 10c-heptadecenoic | C17:1 | 0.08 ± 0.05 |
| Stearic acid | C18:0 | 3.19 ± 0.05 |
| Oleic acid | C18:1 n-9 | 38.90 ± 0.02 |
| cis-vaccenic acid | C18:1 n-7 | 3.23 ± 0.03 |
| Linoleic acid | C18:2 n-6 | 14.68 ± 0.16 |
| Linolenic acid (alpha) | C18:3 n-3 | 4.11 ± 0.02 |
| Linolenic acid (Gamma) | C18:3 n-6 | 0.18 ± 0.01 |
| Stearidonic acid | C18:4 n-3 | 0.72 ± 0.01 |
| Arachidic acid | C20:0 | 0.29 ± 0.01 |
| Gadoleic acid | C20:1 n-11 | 0.19 ± 0.01 |
| Eicosenoic acid | C20:1 n-9 | 1.99 ± 0.02 |
| Paullinic acid | C20:1 n-7 | 0.11 ± 0.01 |
| Eicosadienoic acid | C20:2 | 0.09 ± 0.01 |
| Eicosatrienoic acid | C20:3 n-3 | 0.27 ± 0.01 |
| Dihomo-gamma-linolenic acid | C20:3 n-6 | 0.86 ± 0.02 |
| Eicosatetraenoic acid | C20:4 n-3 | 0.59 ± 0.02 |
| Arachidonic acid | C20:4 n-6 | 0.29 ± 0.01 |
| Eicosapentaenoic Acid (EPA) | C20:5 n-3 | 3.50 ± 0.06 |
| Heneicosapentaenoic acid | C21:5 n-3 | 0.23 ± 0.02 |
| Erucic acid | C22:1 n-9 | 0.34 ± 0.02 |
| Docosapentaenoic acid | C22:5 n-3 | 1.53 ± 0.03 |
| Docosapentaenoic acid | C22:5 n-6 | 0.15 ± 0.04 |
| Docosahexaenoic Acid (DHA) | C22:6 n-3 | 3.57 ± 0.12 |
| Nervonic acid | C24:1 n-9 | 0.20± 0.01 |
| Total Fatty acid Omega-3 | 14.5 ± 0.25 | |
| Total identified | 97.94 ± 0.02 | |
| Total unidentified | 2.06 ± 0.02 | |
| Peroxide value (PV), (meq active oxygen kg−1 oil) | 0.29 ± 0.01 | |
| p-Anisidine Value (AV) Totox Value | 3.70 ± 0.33 3.99 ± 0.34 | |
| Independent and Response Variables of CPUFA | ||||||
|---|---|---|---|---|---|---|
| Run | Urea/FA Content Ratio | Crystallization Temperature (°C) | Crystallization Time (h) | EPA (g/100 g Total FA) | DHA (g/100 g Total FA) | EPA + DHA (g/100 g Total FA) |
| 1 | 0 | 4.0 | 12 | 3.12 | 3.41 | 6.53 |
| 2 | 10 | 4.0 | 12 | 16.38 | 25.08 | 41.46 |
| 3 | 0 | 25.0 | 12 | 3.14 | 3.40 | 6.54 |
| 4 | 10 | 25.0 | 12 | 11.24 | 13.12 | 24.36 |
| 5 | 0 | 14.5 | 0 | 3.29 | 3.57 | 6.86 |
| 6 | 10 | 14.5 | 0 | 6.87 | 7.76 | 14.63 |
| 7 | 0 | 14.5 | 24 | 3.16 | 3.61 | 6.77 |
| 8 | 10 | 14.5 | 24 | 10.23 | 12.4 | 22.63 |
| 9 | 5 | 4.0 | 0 | 9.32 | 11.94 | 21.26 |
| 10 | 5 | 25.0 | 0 | 6.67 | 8.46 | 15.13 |
| 11 | 5 | 4.0 | 24 | 12.42 | 15.63 | 28.05 |
| 12 | 5 | 25.0 | 24 | 11.74 | 13.89 | 25.63 |
| 13 | 5 | 14.5 | 12 | 10.73 | 13.09 | 23.82 |
| 14 | 5 | 14.5 | 12 | 10.65 | 13.20 | 23.85 |
| 15 | 5 | 14.5 | 12 | 10.58 | 13.11 | 23.69 |
| Independent Variables | Sum of Squares | Medium Square | F | p-Value |
|---|---|---|---|---|
| A: Urea/FA content ratio | 729.238 | 729.238 | 100,816.32 | 0.0000 |
| B: Crystallization temp. | 82.1762 | 82.1762 | 11,360.76 | 0.0001 |
| C: Crystallization time | 79.38 | 79.38 | 10,974.19 | 0.0001 |
| AA | 177.301 | 177.301 | 24,511.71 | 0.0000 |
| AB | 73.188 | 73.188 | 10,118.16 | 0.0001 |
| AC | 16.362 | 16.362 | 2262.03 | 0.0004 |
| BB | 30.3161 | 30.3161 | 4191.17 | 0.0002 |
| BC | 3.44102 | 3.44102 | 475.72 | 0.0021 |
| CC | 63.1192 | 63.1192 | 8726.15 | 0.0001 |
| Lack of fit | 126.111 | 42.0371 | 5811.58 | 0.0002 |
| Pure error | 0.0144667 | 0.00723333 | ||
| Total (corr.) | 1385.28 |
| A. Optimal Values for Maximum CPUFA and Optimal Values for CDSO | ||
| CPUFA: Maximize EPA + DHA Optimum Value = 38.35 (g/100 g total FA) | CDSO: EPA + DHA Similar Content to DSO Optimum value = 7.10 (g/100 g total FA) | |
| Independent Variables | Process Optimum Variables | Process Optimum Variables |
| Urea/FA content ratio | 9.997 | 0.047 |
| Crystallization temp, °C | 4.000 | 13.400 |
| Crystallization time, h | 18.120 | 17.380 |
| B. Validation of CPUFA and CDSO by Fatty Acid Composition | ||
| FA | CPUFA (g/100 g total FA) | CDSO (g/100 g total FA) |
| C14:0 | 0.03 | 2.17 |
| C16:0 | 0.25 | 11.25 |
| C18:0 | 0.14 | 4.11 |
| C18:1 | 1.70 | 40.76 |
| C18:2 | 19.08 | 14.7 |
| C18:3 | 8.58 | 4.43 |
| EPA | 15.80 | 3.34 |
| DHA | 22.55 | 3.76 |
| EPA + DHA | 38.35 | 7.10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinosa, A.; Ross, A.; Dovale-Rosabal, G.; Pino-de la Fuente, F.; Uribe-Oporto, E.; Sacristán, C.; Ruiz, P.; Valenzuela, R.; Romero, N.; Aubourg, S.P.; et al. EPA/DHA Concentrate by Urea Complexation Decreases Hyperinsulinemia and Increases Plin5 in the Liver of Mice Fed a High-Fat Diet. Molecules 2020, 25, 3289. https://doi.org/10.3390/molecules25143289
Espinosa A, Ross A, Dovale-Rosabal G, Pino-de la Fuente F, Uribe-Oporto E, Sacristán C, Ruiz P, Valenzuela R, Romero N, Aubourg SP, et al. EPA/DHA Concentrate by Urea Complexation Decreases Hyperinsulinemia and Increases Plin5 in the Liver of Mice Fed a High-Fat Diet. Molecules. 2020; 25(14):3289. https://doi.org/10.3390/molecules25143289
Chicago/Turabian StyleEspinosa, Alejandra, Andrés Ross, Gretel Dovale-Rosabal, Francisco Pino-de la Fuente, Ernesto Uribe-Oporto, Camila Sacristán, Paulina Ruiz, Rodrigo Valenzuela, Nalda Romero, Santiago P. Aubourg, and et al. 2020. "EPA/DHA Concentrate by Urea Complexation Decreases Hyperinsulinemia and Increases Plin5 in the Liver of Mice Fed a High-Fat Diet" Molecules 25, no. 14: 3289. https://doi.org/10.3390/molecules25143289