
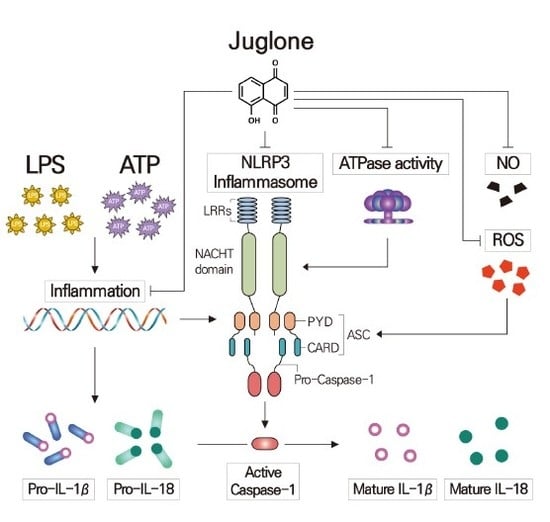
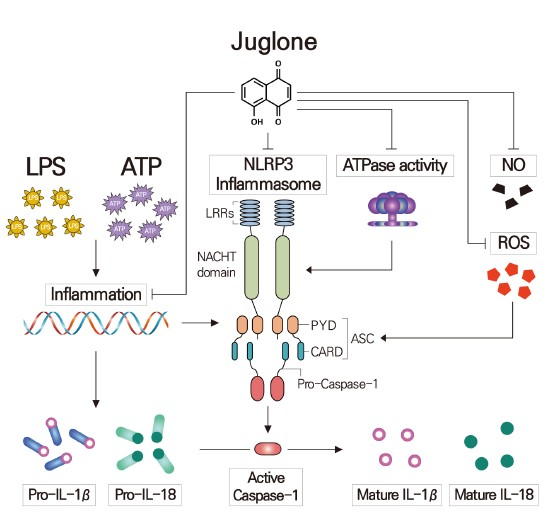
Juglone Suppresses LPS-induced Inflammatory Responses and NLRP3 Activation in Macrophages

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Effects of Juglone on Cell Viability
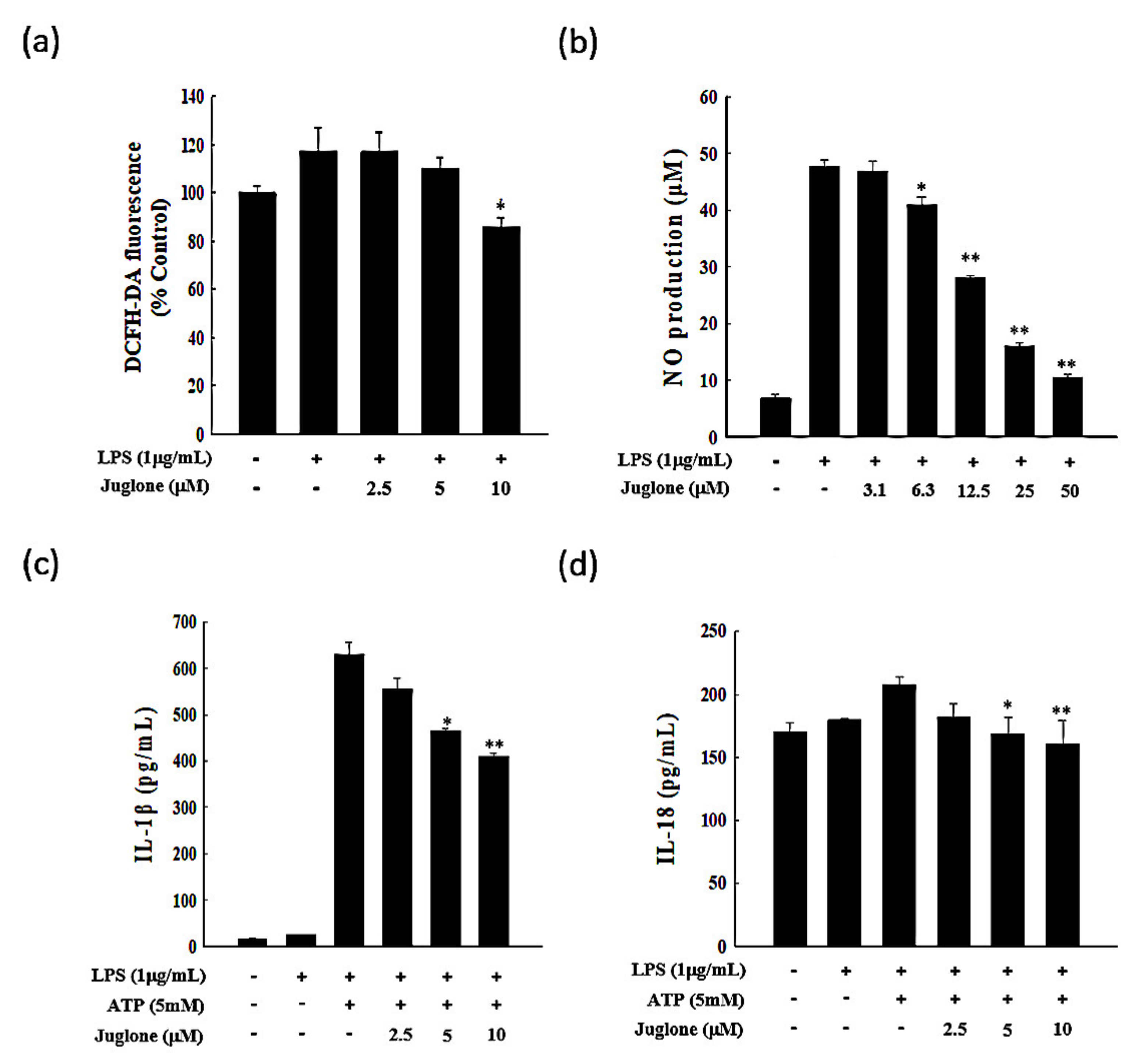
2.2. Effects of Juglone on Reactive Oxygen Species (ROS) Production
2.3. Effects of Juglone on Nitric Oxide (NO) Production
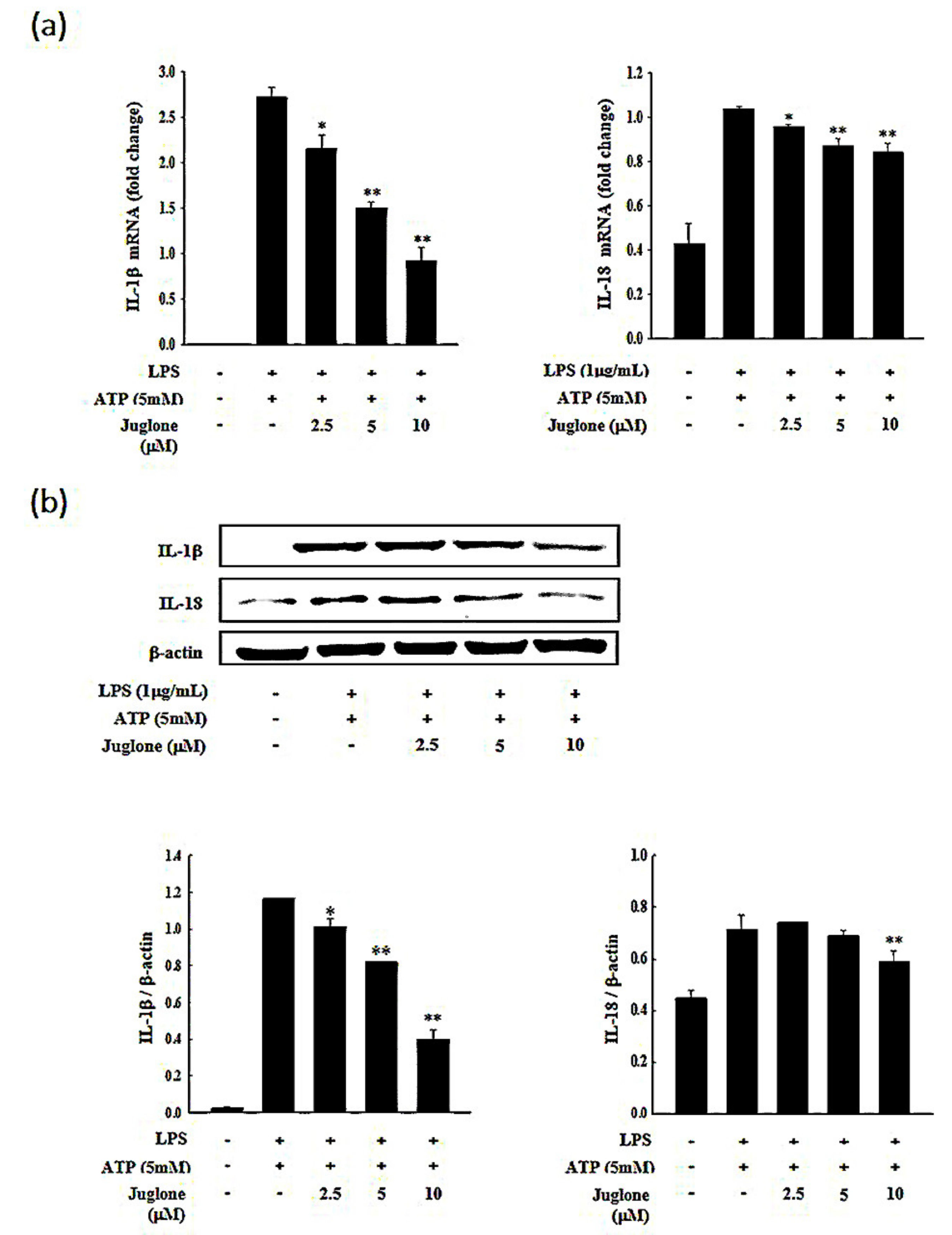
2.4. Effects of Juglone on the Secretion of the Pro-Inflammatory Cytokines IL-1β and IL-18
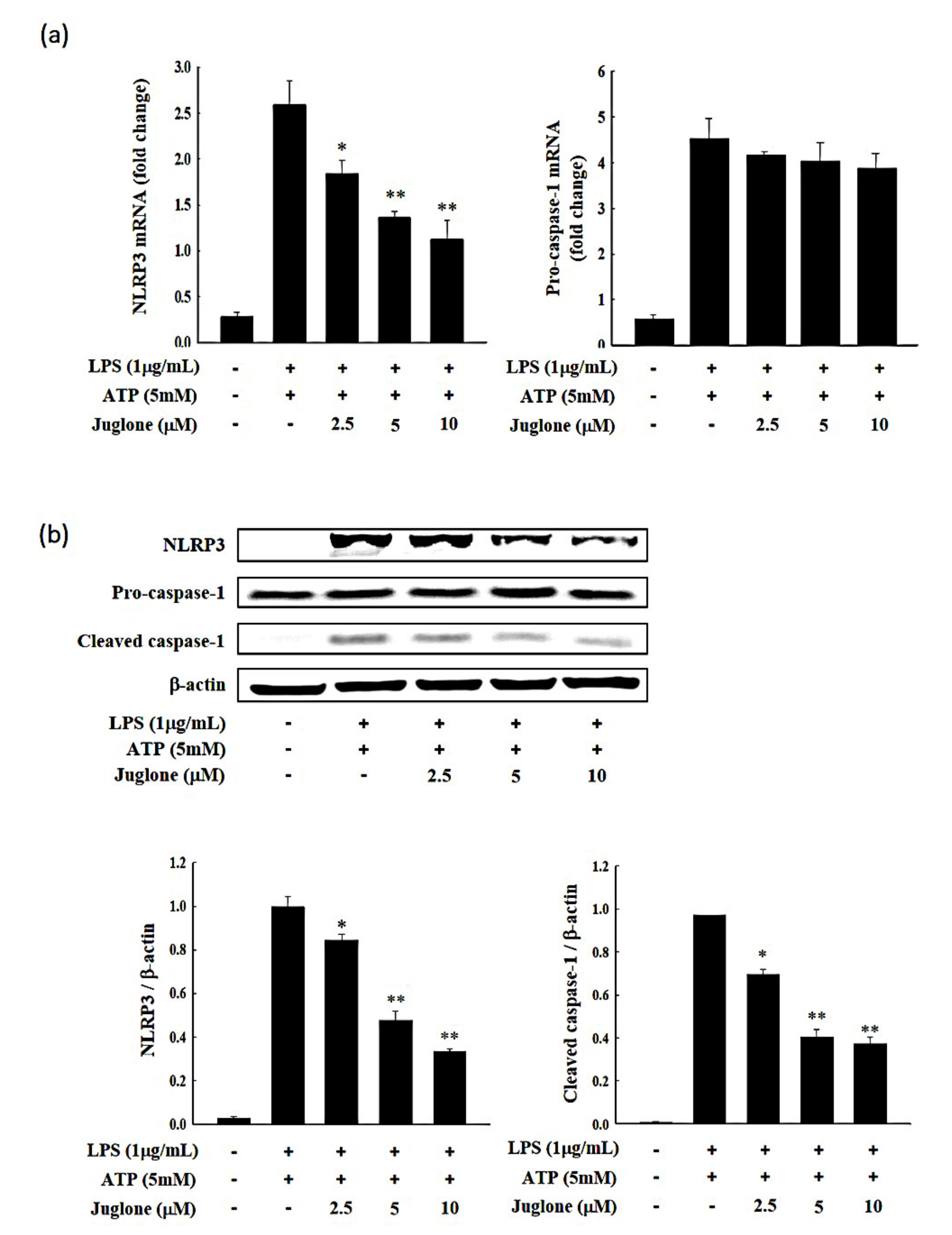
2.5. Effects of Juglone on NLRP3 and Caspase-1 Expression
2.6. Effects of Juglone on the ATPase Activity of NLRP3
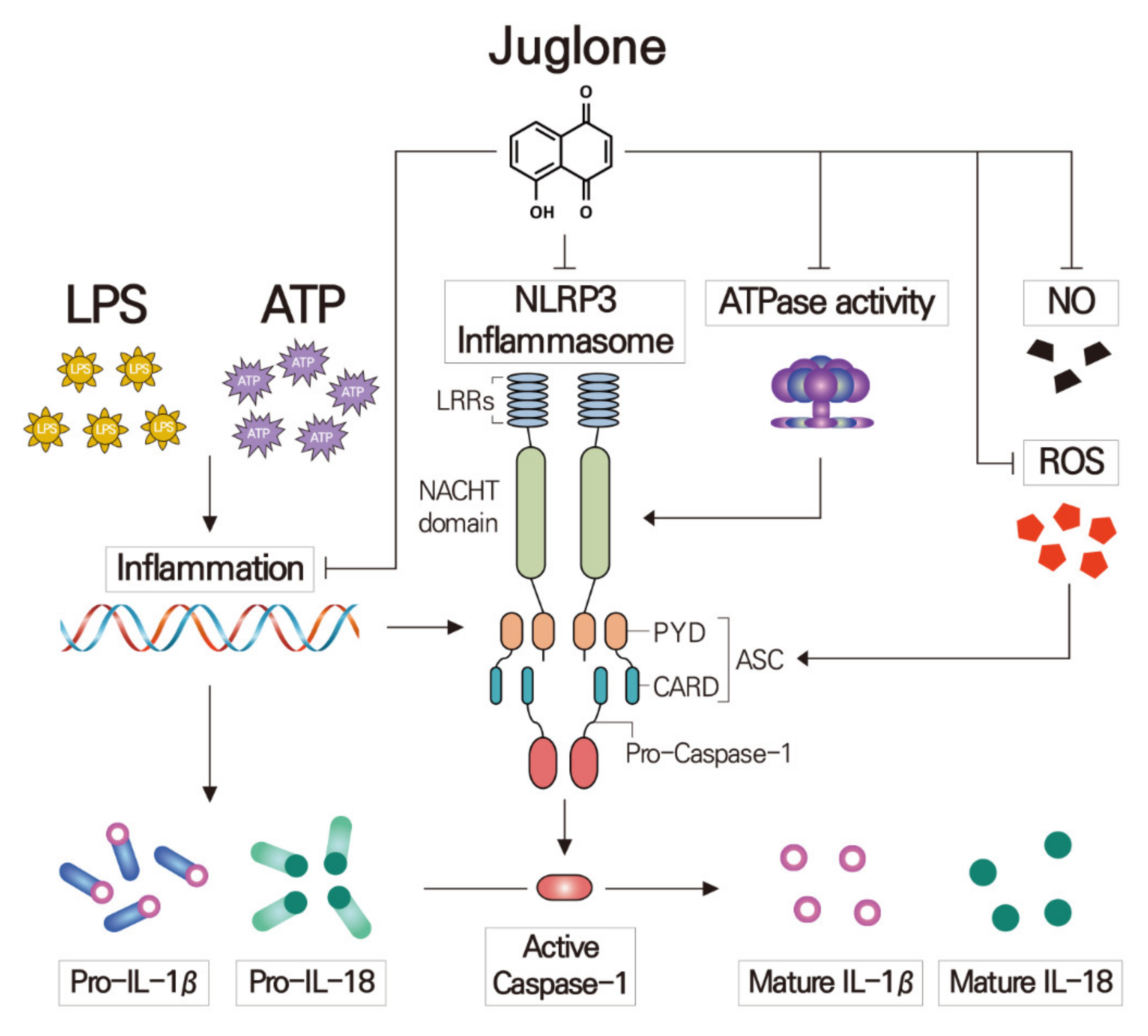
3. Discussion
4. Materials and Methods
4.1. Chemical Compounds
4.2. Cell Cultures
4.3. Cell Viability
4.4. Measurement of Nitric Oxide (NO)
4.5. Measurement of the Antioxidative Capacity of Juglone
4.6. Measurement of Cytokine Release
4.7. RNA Isolation and cDNA Synthesis
4.8. Real-Time Reverse Transcriptase Quantitative Polymerase Chain Reaction (RT-qPCR)
4.9. Western Blot Analysis
4.10. ATPase Activity Measurement
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen Recognition by the Innate Immune System. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-Likereceptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between Toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, M.; Ceballos-Olvera, I.; Del Barrio, L.; Re, F. Role of the Inflammasome, IL-1β, and IL-18 in Bacterial Infections. Sci. World J. 2011, 11, 2037–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinon, F.; Mayor, A.; Tschopp, J. The Inflammasomes: Guardians of the Body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef] [Green Version]
- Perregaux, D.; Gabel, C.A. Interleukin-1 beta maturation and release in response to ATP and nigericin. Evidence that potassium depletion mediated by these agents is a necessary and common feature of their activity. J. Boil. Chem. 1994, 269, 15195–15203. [Google Scholar]
- Walev, I.; Klein, J.; Husmann, M.; Valeva, A.; Strauch, S.; Wirtz, H.; Weichel, O.; Bhakdi, S. Potassium Regulates IL-1β Processing Via Calcium-Independent Phospholipase A2. J. Immunol. 2000, 164, 5120–5124. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2010, 469, 221–225. [Google Scholar] [CrossRef]
- Sorbara, M.T.; Girardin, S.E. Mitochondrial ROS fuel the inflammasome. Cell Res. 2011, 21, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Heid, M.E.; Keyel, P.A.; Kamga, C.; Shiva, S.; Watkins, S.C.; Salter, R.D. Mitochondrial reactive oxygen species induces NLRP3-dependent lysosomal damage and inflammasome activation. J. Immunol. 2013, 191, 5230–5238. [Google Scholar] [CrossRef] [Green Version]
- Bauernfeind, F.; Bartok, E.; Rieger, A.; Franchi, L.; Núñez, G.; Hornung, V. Cutting edge: Reactive oxygen species inhibitors block priming, but not activation, of the NLRP3 inflammasome. J. Immunol. 2011, 187, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aithal, B.K.; Kumar, M.S.; Rao, B.N.; Udupa, N.; Rao, B.S. Juglone, a naphthoquinone from walnut, exerts cytotoxic and genotoxic effects against cultured melanoma tumor cells. Cell Boil. Int. 2009, 33, 1039–1049. [Google Scholar] [CrossRef]
- Soderquist, C.J. Juglone and allelopathy. J. Chem. Educ. 1973, 50, 782. [Google Scholar] [CrossRef]
- Seetha, A.; Devaraj, H.; Sudhandiran, G. Indomethacin and juglone inhibit inflammatory molecules to induce apoptosis and colon cnacer cells. J. Biochem. Mol. Toxicol. 2020, 34, e22433. [Google Scholar] [CrossRef]
- Peng, X.; Nie, Y.; Wu, J.; Huang, Q.; Cheng, Y. Juglone prevents metabolic endotoxemia-induced hepatitis and neuroinflammation via suppressing TLR4/NF-κB signaling pathway in high-fat diet rats. Biochem. Biophys. Res. Commun. 2015, 462, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Reese, S.; Vidyasagar, A.; Jacobson, L.; Acun, Z.; Esnault, S.; Hullett, D.; Malter, J.S.; Djamali, A. The Pin 1 inhibitor juglone attenuates kidney fibrogenesis via Pin 1-independent mechanisms in the unilateral ureteral occlusion model. Fibrogenesis Tissue Repair 2010, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-B.; Zou, C.-L.; Duan, Y.-X.; Wu, F.; Li, G. Activity guided isolation and modification of juglone from Juglans regia as potent cytotoxic agent against lung cancer cell lines. BMC Complement. Altern. Med. 2015, 15, 396. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.T.C.; Osman, H.; Mohamad, S.; Kamaruddin, A.H. Synthesis and antibacterial activity of juglone derivatives. J. Chem. Chem. Eng. 2012, 6, 84–89. [Google Scholar]
- Zakavi, F.; Hagh, L.G.; Daraeighadikolaei, A.; Sheikh, A.F.; Daraeighadikolaei, A.; Shooshtari, Z.L. Antibacterial effect of Juglans regia bark against oral pathologic bacteria. Int. J. Dent. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, M.; Mak, K.-K.; Devadoss, T.; Thanikachalam, P.V.; Sakirolla, R.; Choudhury, H.; Pichika, M.R. Construction of a novel quinoxaline as a new class of Nrf2 activator. BMC Chem. 2019, 13, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariathasan, S.; Weiss, D.S.; Newton, K.; McBride, J.; O’Rourke, K.; Roose-Girma, M.; Lee, W.P.; Weinrauch, Y.; Monack, D.M.; Dixit, V.M. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 2006, 440, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Benko, S.; Philpott, D.J.; Girardin, S.E. The microbial and danger signals that activate Nod-like receptors. Cytokine 2008, 43, 368–373. [Google Scholar] [CrossRef]
- Galas, M.; Dourlen, P.; Bégard, S.; Ando, K.; Blum, D.; Hamdane, M.; Buée, L. The Peptidylprolylcis/trans-Isomerase Pin1 Modulates Stress-induced Dephosphorylation of Tau in Neurons. J. Boil. Chem. 2006, 281, 19296–19304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.-J.; Mu, D.; Jiang, M.-D.; Zheng, S.-M.; Zhang, Y.; He, S.; Weng, M.; Zeng, W.-Z. Hepatoprotective effect of juglone on dimethylnitrosamine-induced liver fibrosis and its effect on hepatic antioxidant defence and the expression levels of α-SMA and collagen III. Mol. Med. Rep. 2015, 12, 4095–4102. [Google Scholar] [CrossRef] [Green Version]
- Sugie, S.; Okamoto, K.; Rahman, K.; Tanaka, T.; Kawai, K.; Yamahara, J.; Mori, H. Inhibitory effects of plumbagin and juglone on azoxymethane-induced intestinal carcinogenesis in rats. Cancer Lett. 1998, 127, 177–183. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin 1 and interleukin 18 as mediators of inflammation and the aging process. Am. J. Clin. Nutr. 2006, 83, 447S–455S. [Google Scholar] [CrossRef] [Green Version]
- Martinon, F. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 2010, 40, 616–619. [Google Scholar] [CrossRef]
- Valencia, A.; Morán, J. Reactive oxygen species induce different cell death mechanisms in cultured neurons. Free Radic. Boil. Med. 2004, 36, 1112–1125. [Google Scholar] [CrossRef] [PubMed]
- Sharif, H.; Wang, L.; Wang, W.L.; Magupalli, V.G.; Andreeva, L.; Qiao, Q.; Hauenstein, A.V.; Wu, Z.; Núñez, G.; Mao, Y.; et al. Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome. Nature 2019, 570, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.M.; Rinna, A.; Forman, H.J.; Ventura, A.L.M.; Persechini, P.; Ojcius, D.M. ATP activates a reactive oxygen species-dependent oxidative stress response and secretion of proinflammatory cytokines in macrophages. J. Boil. Chem. 2006, 282, 2871–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantino, S.; Paneni, F.; Lüscher, T.F.; Cosentino, F. Pin1 inhibitor Juglone prevents diabetic vascular dysfunction. Int. J. Cardiol. 2016, 203, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.L.; Lenz, L.L. Distinct Licensing of IL-18 and IL-1β Secretion in Response to NLRP3 Inflammasome Activation. PLoS ONE 2012, 7, e45186. [Google Scholar] [CrossRef] [Green Version]
- Pisha, E.; Chai, H.; Lee, I.-S.; Chagwedera, T.E.; Farnsworth, N.R.; Cordell, G.A.; Beecher, C.W.W.; Fong, H.H.; Kinghorn, A.D.; Brown, D.M.; et al. Discovery of betulinic acid as a selective inhibitor of human melanoma that functions by induction of apoptosis. Nat. Med. 1995, 1, 1046–1051. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Rapid Peroxyl Radical Scavenging Capacity (PSC) Assay for Assessing both Hydrophilic and Lipophilic Antioxidants. J. Agric. Food Chem. 2005, 53, 6572–6580. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample compound (juglone) is available from the authors or Merck (Darmstadt, Germany). |






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.-H.; Kim, H.-K.; Lee, J.-H.; Jo, S.-I.; Won, H.-M.; Lee, G.-S.; Lee, H.-S.; Nam, K.-W.; Kim, W.-J.; Han, M.-D. Juglone Suppresses LPS-induced Inflammatory Responses and NLRP3 Activation in Macrophages. Molecules 2020, 25, 3104. https://doi.org/10.3390/molecules25133104
Kim N-H, Kim H-K, Lee J-H, Jo S-I, Won H-M, Lee G-S, Lee H-S, Nam K-W, Kim W-J, Han M-D. Juglone Suppresses LPS-induced Inflammatory Responses and NLRP3 Activation in Macrophages. Molecules. 2020; 25(13):3104. https://doi.org/10.3390/molecules25133104
Chicago/Turabian StyleKim, Nam-Hun, Hong-Ki Kim, Ji-Hak Lee, Seung-Il Jo, Hye-Min Won, Gyeong-Seok Lee, Hyoun-Su Lee, Kung-Woo Nam, Wan-Jong Kim, and Man-Deuk Han. 2020. "Juglone Suppresses LPS-induced Inflammatory Responses and NLRP3 Activation in Macrophages" Molecules 25, no. 13: 3104. https://doi.org/10.3390/molecules25133104