
Dual miRNases for Triple Incision of miRNA Target: Design Concept and Catalytic Performance

Abstract
:1. Introduction
2. Results
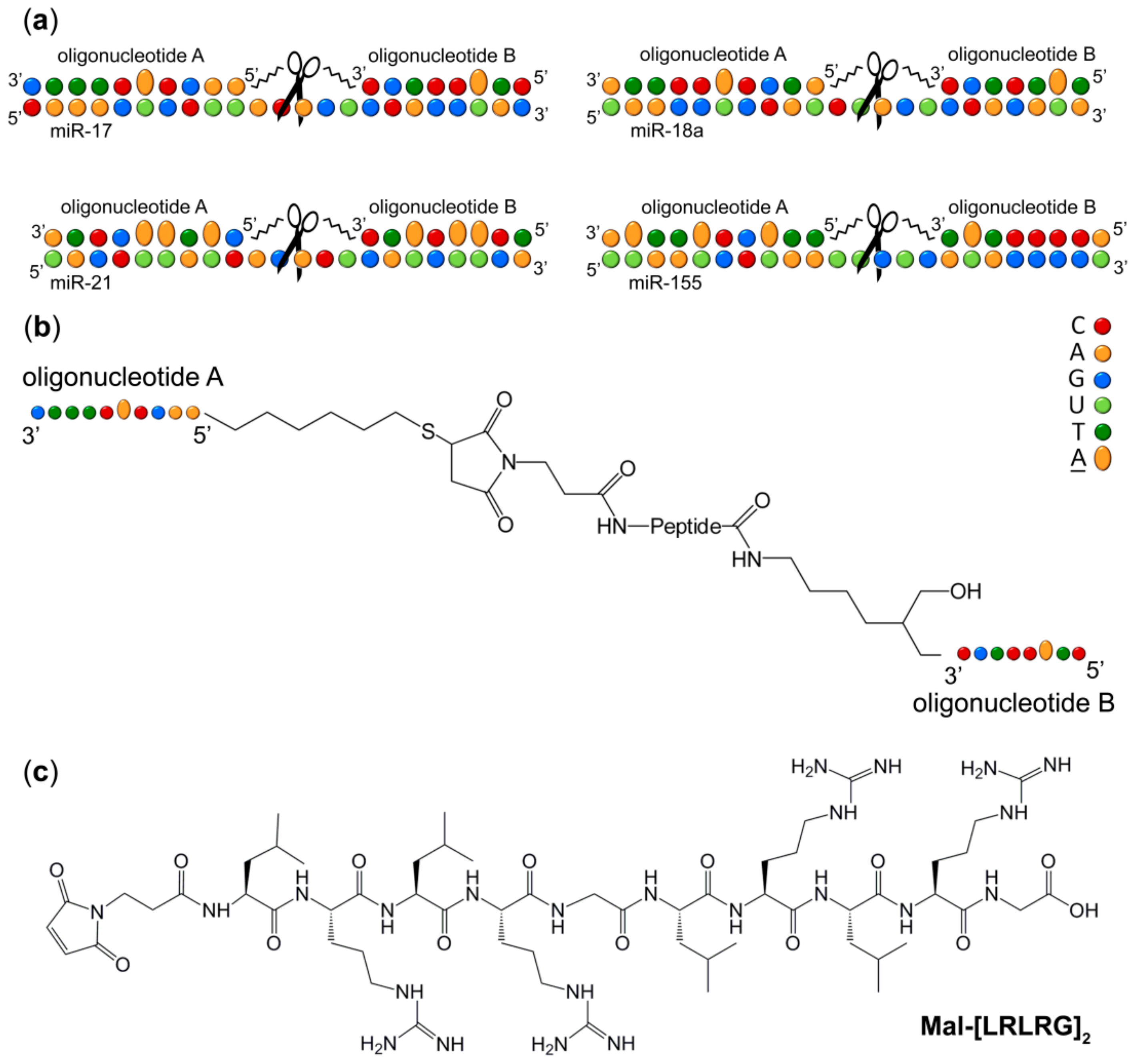
2.1. Design Strategy of Dual Conjugates
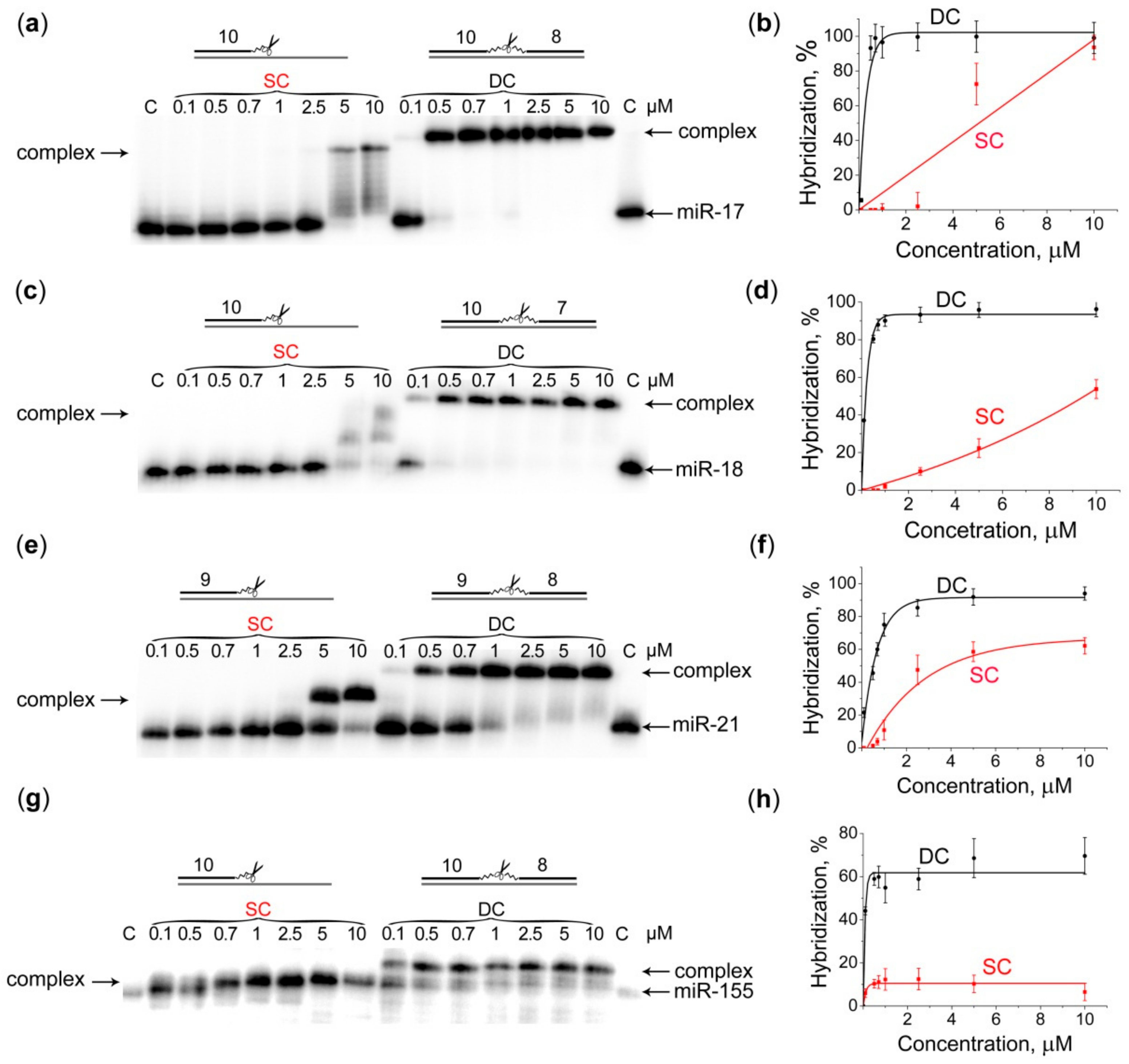
2.2. Hybridization of Dual Conjugates with miRNA Targets
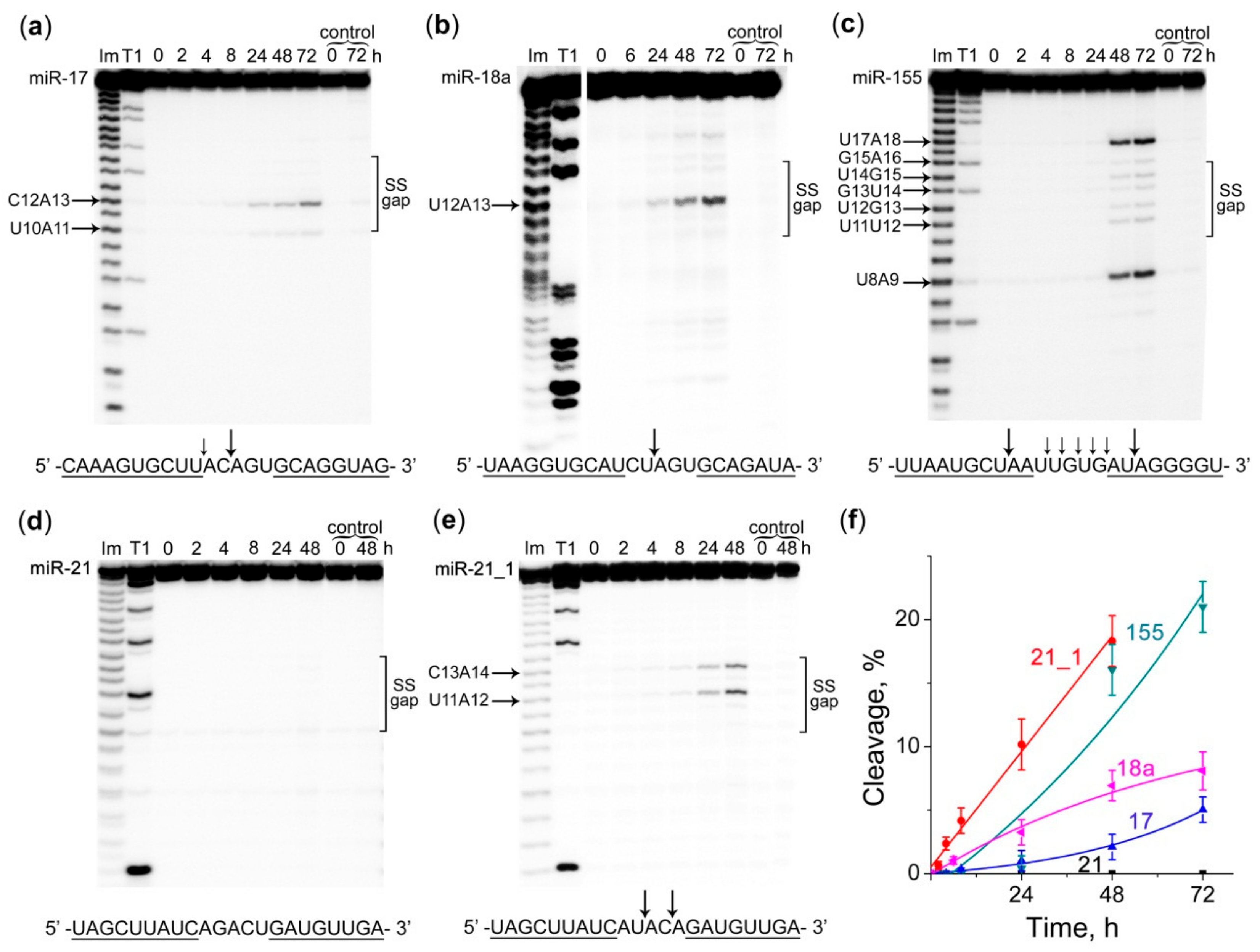
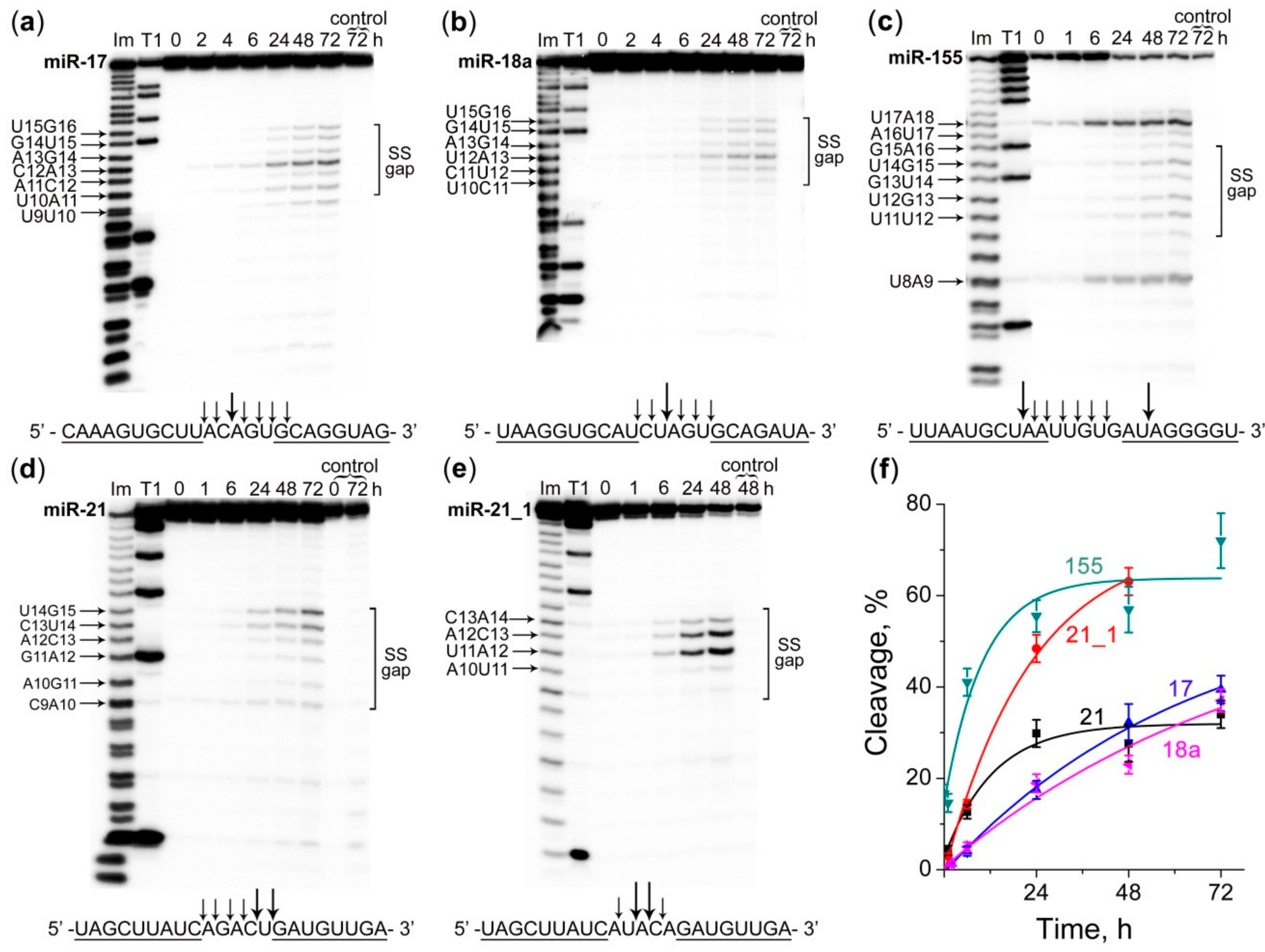
2.3. Cleavage Profile and Kinetics of miRNA Cleavage by Dual Conjugates
2.4. The Effect of Mg2+ and K+ Concentration on the Ribonuclease Activity of Dual Conjugates
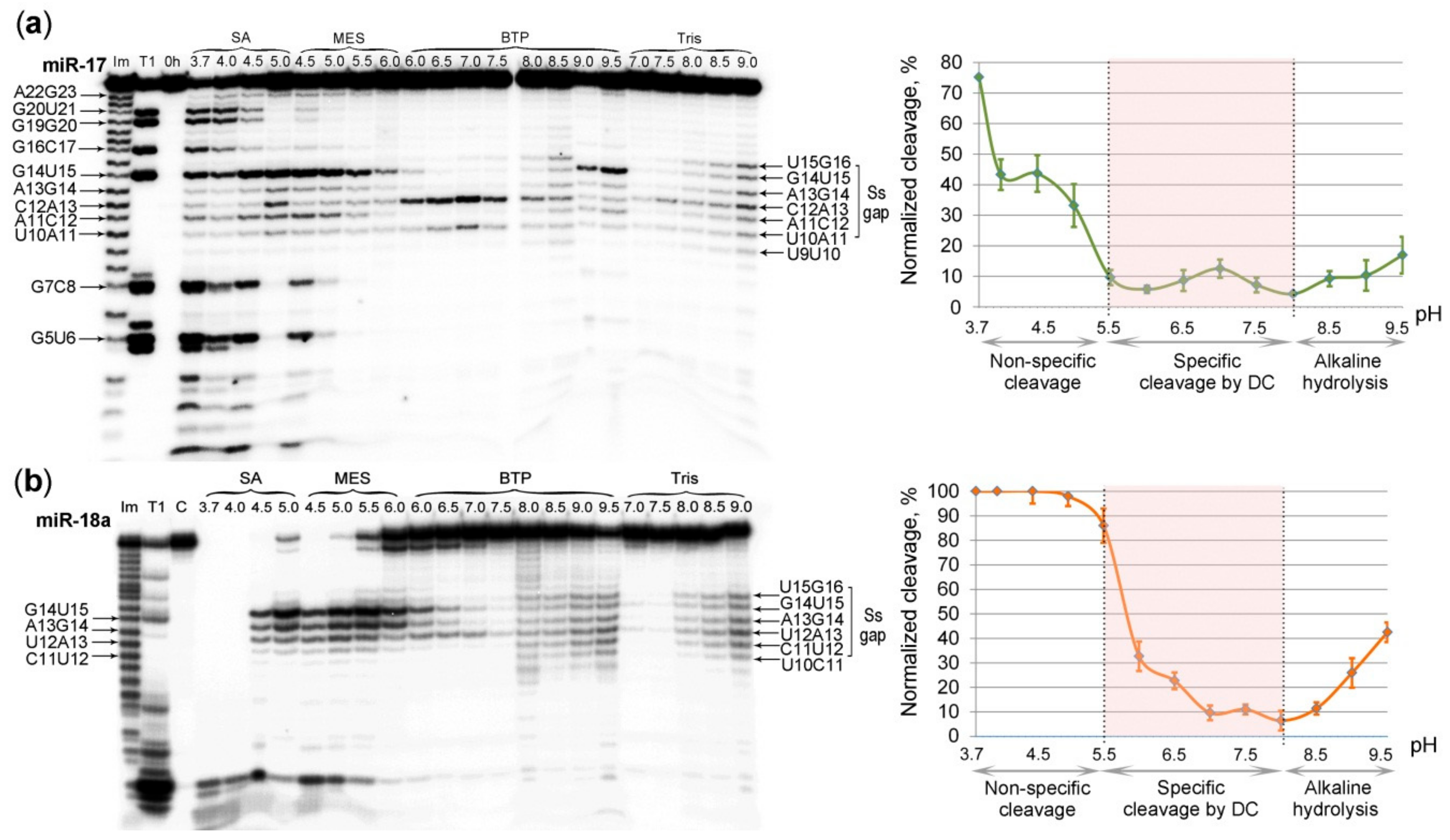
2.5. The Effect of pH on the Ribonuclease Activity of Dual Conjugates
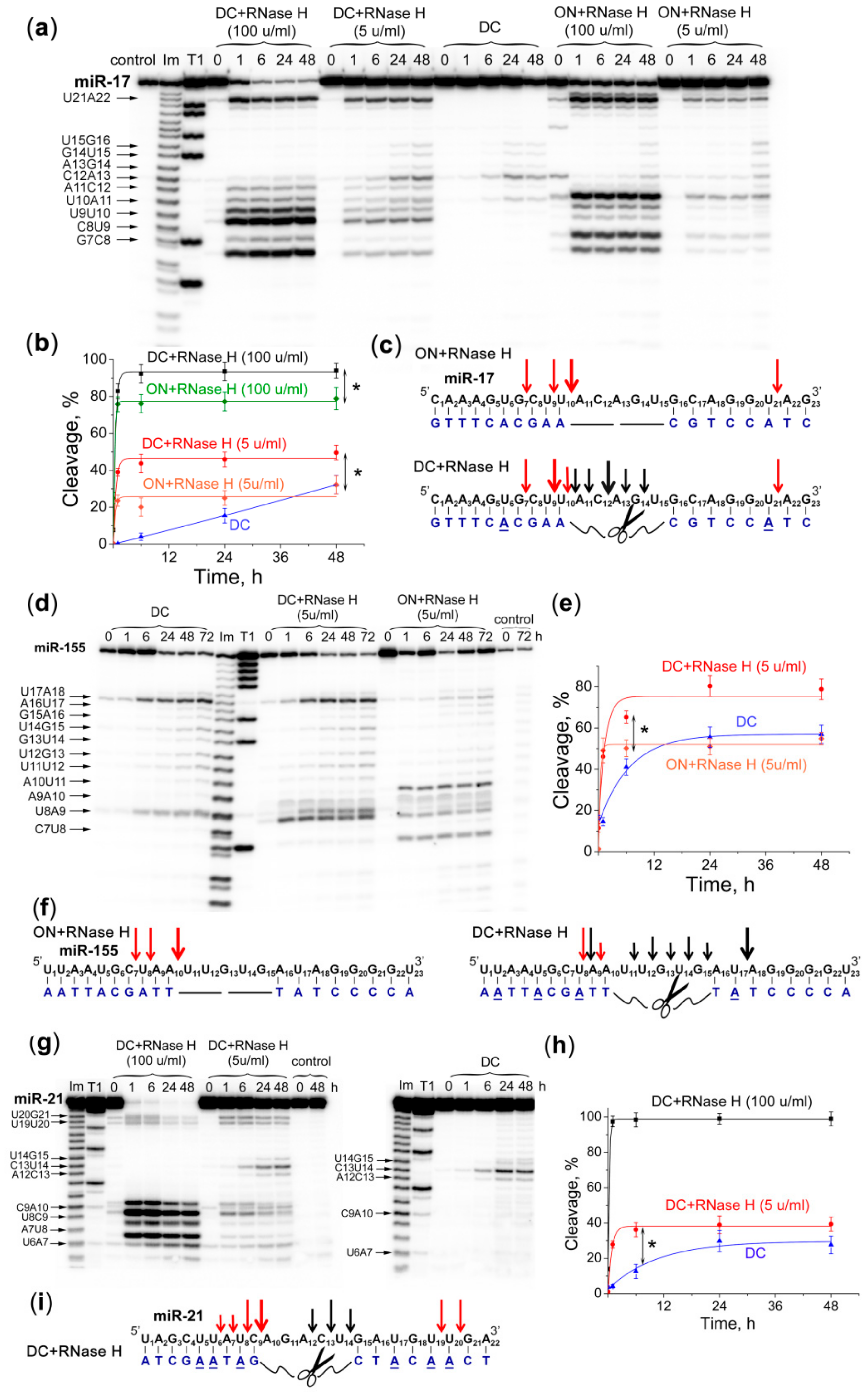
2.6. Efficiency and Specificity of miRNA Cleavage by Dual Conjugates in the Presence of RNase H
3. Discussion
4. Materials and Methods
4.1. Oligonucleotides
4.2. Synthesis of the Conjugates
4.3. RNA Labelling
4.4. Gel-Retardation assay
4.5. Ribonuclease Activity Assay
4.6. Ribonuclease Activity Assay Using Dual Conjugates and RNase H
4.7. Ribonuclease Activity Assay at Different Concentrations of MgCl2
4.8. Ribonuclease Activity Assay at Different Concentrations of KCl
4.9. pH Profile of Ribonuclease Activity of DCs
4.10. Stability of DCs and ONs in 10% FBS
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sheu-Gruttadauria, J.; MacRae, I.J. Phase Transitions in the Assembly and Function of Human miRISC. Cell 2018, 173, 946–957.e16. [Google Scholar] [CrossRef] [PubMed]
- Sheu-Gruttadauria, J.; Pawlica, P.; Klum, S.M.; Wang, S.; Yario, T.A.; Schirle Oakdale, N.T.; Steitz, J.A.; MacRae, I.J. Structural Basis for Target-Directed MicroRNA Degradation. Mol. Cell 2019, 75, 1243–1255.e7. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, S.; Patutina, O. Enhanced inhibition of tumorigenesis using combinations of miRNA-targeted therapeutics. Front. Pharmacol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Hogan, D.J.; Vincent, T.M.; Fish, S.; Marcusson, E.G.; Bhat, B.; Nelson Chau, B.; Zisoulis, D.G. Anti-miRs competitively inhibit microRNAs in argonaute complexes. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef]
- Ariyoshi, J.; Momokawa, D.; Eimori, N.; Kobori, A.; Murakami, A.; Yamayoshi, A. Development of Novel Antisense Oligonucleotides for the Functional Regulation of RNA-Induced Silencing Complex (RISC) by Promoting the Release of microRNA from RISC. Bioconjug. Chem. 2015, 26, 2454–2460. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, J.; Matsuyama, Y.; Kobori, A.; Murakami, A.; Sugiyama, H.; Yamayoshi, A. Effective Anti-miRNA Oligonucleotides Show High Releasing Rate of MicroRNA from RNA-Induced Silencing Complex. Nucleic Acid Ther. 2017, 27, 303–308. [Google Scholar] [CrossRef]
- Patutina, O.A.; Miroshnichenko, S.K.; Mironova, N.L.; Sen’kova, A.V.; Bichenkova, E.V.; Clarke, D.J.; Vlassov, V.V.; Zenkova, M.A. Catalytic knockdown of MIR-21 by artificial Ribonuclease: Biological performance in tumor model. Front. Pharmacol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Patutina, O.A.; Bazhenov, M.A.; Miroshnichenko, S.K.; Mironova, N.L.; Pyshnyi, D.V.; Vlassov, V.V.; Zenkova, M.A. Peptide-oligonucleotide conjugates exhibiting pyrimidine-X cleavage specificity efficiently silence miRNA target acting synergistically with RNase, H. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Patutina, O.A.; Bichenkova, E.V.; Miroshnichenko, S.K.; Mironova, N.L.; Trivoluzzi, L.T.; Burusco, K.K.; Bryce, R.A.; Vlassov, V.V.; Zenkova, M.A. miRNases: Novel peptide-oligonucleotide bioconjugates that silence miR-21 in lymphosarcoma cells. Biomaterials 2017, 122, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Danneberg, F.; Ghidini, A.; Dogandzhiyski, P.; Kalden, E.; Strömberg, R.; Göbel, M.W. Sequence-specific RNA cleavage by PNA conjugates of the metal-free artificial ribonuclease tris(2-aminobenzimidazole). Beilstein J. Org. Chem. 2015, 11, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaglione, M.; Milano, G.; Chambery, A.; Moggio, L.; Romanelli, A.; Messere, A. PNA-based artificial nucleases as antisense and anti-miRNA oligonucleotide agents. Mol. Biosyst. 2011, 7, 2490–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staroseletz, Y.; Williams, A.; Burusco, K.K.; Alibay, I.; Vlassov, V.V.; Zenkova, M.A.; Bichenkova, E.V. ‘Dual’ peptidyl-oligonucleotide conjugates: Role of conformational flexibility in catalytic cleavage of RNA. Biomaterials 2017, 112, 44–61. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, Y.B.; Zhang, X.H.; Yu, X.L.; Wang, Z.B.; Cheng, X.C. MicroRNA-21 gene and cancer. Med. Oncol. 2013, 30, 1–9. [Google Scholar] [CrossRef]
- Li, S.; Liang, Z.; Xu, L.; Zou, F. MicroRNA-21: A ubiquitously expressed pro-survival factor in cancer and other diseases. Mol. Cell. Biochem. 2012, 360, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Eis, P.S.; Tam, W.; Sun, L.; Chadburn, A.; Li, Z.; Gomez, M.F.; Lund, E.; Dahlberg, J.E. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc. Natl. Acad. Sci. USA 2005, 102, 3627–3632. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.Y.; Xu, Z.H.; Chen, Y.L.; Lu, Z.Y.; Shen, D.Y.; Lu, J.Y.; Zheng, Q.M.; Wang, L.Y.; Xu, L.W.; et al. The prognostic value of miRNA-18a-5p in clear cell renal cell carcinoma and its function via the miRNA-18a-5p/HIF1A/PVT1 pathway. J. Cancer 2020, 11, 2737–2748. [Google Scholar] [CrossRef]
- Mai, S.J.; Xiao, R.W.; Shi, L.; Zhou, X.M.; Yang, T.; Zhang, M.Y.; Weng, N.Q.; Zhao, X.G.; Wang, R.Q.; Liu, J.; et al. MicroRNA-18a promotes cancer progression through SMG1 suppression and mTOR pathway activation in nasopharyngeal carcinoma. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, R.; Van Roosbroeck, K. miR-155 in cancer drug resistance and as target for miRNA-based therapeutics. Cancer Metastasis Rev. 2018, 37, 33–44. [Google Scholar] [CrossRef]
- Williams, A.; Staroseletz, Y.; Zenkova, M.A.; Jeannin, L.; Aojula, H.; Bichenkova, E.V. Peptidyl-Oligonucleotide Conjugates Demonstrate Efficient Cleavage of RNA in a Sequence-Specific Manner. Bioconjug. Chem. 2015, 26, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.; Tinoco, I.; Chollet, A. Thermodynamic studies of base pairing involving 2,6-diaminopurine. Nucleic Acids Res. 1988, 16, 5115–5122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirnos, M.D.; Khudyakov, Y.I.; Alexandrushkina, N.I.; Vanyushin, B.F. 2-aminoadenine is an adenine substituting for a base in S-2L cyanophage DNA. Nature 1977, 270, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Haaima, G.; Hansen, H.F.; Christensen, L.; Dahl, O.; Nielsen, P.E. Increased DNA binding and sequence discrimination of PNA oligomers containing 2,6-diaminopurine. Nucleic Acids Res. 1997, 25, 4639–4643. [Google Scholar] [CrossRef] [Green Version]
- Pounder, R.J.; Stanford, M.J.; Brooks, P.; Richards, S.P.; Dove, A.P. Metal free thiol-maleimide “Click” reaction as a mild functionalisation strategy for degradable polymers. Chem. Commun. 2008, 5158–5160. [Google Scholar] [CrossRef]
- Mironova, N.L.; Pyshnyi, D.V.; Shtadler, D.V.; Fedorova, A.A.; Vlassov, V.V.; Zenkova, M.A. RNase T1 mimicking artificial ribonuclease. Nucleic Acids Res. 2007, 35, 2356–2367. [Google Scholar] [CrossRef] [Green Version]
- Vlassov, V.V.; Zuber, G.; Felden, B.; Behr, J.P.; Giege, R. Cleavage of tRNA With Imidazole and Spermine Imidazole Constructs: A New Approach for Probing RNA Structure. Nucleic Acids Res. 1995, 23, 3161–3167. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, I.L.; Zenkova, M.A.; Gross, H.J.; Vlassov, V.V. Enhanced RNA Cleavage Within Bulge-Loops by an Artificial Ribonuclease. Nucleic Acids Res. 2005, 33, 1201–1212. [Google Scholar] [CrossRef]
- Callaghan, A.J.; Marcaida, M.J.; Stead, J.A.; McDowall, K.J.; Scott, W.G.; Luisi, B.F. Structure of Escherichia coli RNase E catalytic domain and implications for RNA turnover. Nature 2005, 437, 1187–1191. [Google Scholar] [CrossRef]
- Johnson-Buck, A.E.; McDowell, S.E.; Walter, N.G. Metal Ions: Supporting actors in the playbook of small ribozymes. Met. Ions Life Sci. 2011, 9, 175–196. [Google Scholar] [CrossRef] [Green Version]
- Haddy, F.J.; Vanhoutte, P.M.; Feletou, M. Role of potassium in regulating blood flow and blood pressure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.F.; Swayze, E.E. RNA Targeting Therapeutics: Molecular Mechanisms of Antisense Oligonucleotides as a Therapeutic Platform. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Compagno, D.; Lampe, J.N.; Bourget, C.; Kutyavin, I.V.; Yurchenko, L.; Lukhtanov, E.A.; Gorn, V.V.; Gamper, H.B.; Toulmé, J.J. Antisense oligonucleotides containing modified bases inhibit in vitro translation of Leishmania amazonensis mRNAs by invading the mini-exon hairpin. J. Biol. Chem. 1999, 274, 8191–8198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murtola, M.; Ghidini, A.; Virta, P.; Strömberg, R. Zinc Ion-Dependent peptide nucleic acid-based artificial enzyme that cleaves RNA-Bulge size and sequence dependence. Molecules 2017, 22, 1856. [Google Scholar] [CrossRef] [Green Version]
- Luige, O.; Murtola, M.; Ghidini, A.; Strömberg, R. Further probing of Cu2+-dependent PNAzymes acting as artificial RNA restriction enzymes. Molecules 2019, 24, 672. [Google Scholar] [CrossRef] [Green Version]
- Zellmann, F.; Thomas, L.; Scheffer, U.; Hartmann, R.K.; Göbel, M.W. Site-specific cleavage of RNAs derived from the PIM1 30-UTR by a metal-free artificial ribonuclease. Molecules 2019, 24, 807. [Google Scholar] [CrossRef] [Green Version]
- Vormann, J. Magnesium: Nutrition and metabolism. Mol. Asp. Med. 2003, 24, 27–37. [Google Scholar] [CrossRef]
- Pohl, H.R.; Wheeler, J.S.; Murray, H.E. Sodium and potassium in health and disease. In Interrelations between Essential Metal Ions and Human Diseases; Metal Ions in Life Sciences; Springer: Berlin/Heidelberg, Germany, 2013; Volume 13, pp. 29–47. [Google Scholar] [CrossRef]
- Clausen, M.J.V.; Poulsen, H. Sodium/potassium homeostasis in the cell. In Metallomics and the Cell; Metal Ions in Life Sciences; Springer: Berlin/Heidelberg, Germany, 2013; Volume 12, pp. 41–67. [Google Scholar] [CrossRef]
- Mariani, A.; Bonfio, C.; Johnson, C.M.; Sutherland, J.D. PH-Driven RNA Strand Separation under Prebiotically Plausible Conditions. Biochemistry 2018, 57, 6382–6386. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, H.S.; Tate, W.P. Primordial soup or vinaigrette: Did the RNA world evolve at acidic pH? Biol. Direct 2012, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ageno, M.; Dore, E.; Frontali, C. The Alkaline Denaturation of DNA. Biophys. J. 1969, 9, 1281–1311. [Google Scholar] [CrossRef] [Green Version]
- Oivanen, M.; Kuusela, S.; Lönnberg, H. Kinetics and mechanisms for the cleavage and isomerization of the phosphodiester bonds of RNA by bronsted acids and bases. Chem. Rev. 1998, 98, 961–990. [Google Scholar] [CrossRef] [PubMed]
- Fitch, C.A.; Platzer, G.; Okon, M.; Garcia-Moreno, B.E.; McIntosh, L.P. Arginine: Its pKa value revisited. Protein Sci. 2015, 24, 752–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wike-Hooley, J.L.; Haveman, J.; Reinhold, H.S. The relevance of tumour pH to the treatment of malignant disease. Radiother. Oncol. 1984, 2, 343–366. [Google Scholar] [CrossRef]
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood Flow, Oxygen and Nutrient Supply, and Metabolic Microenvironment of Human Tumors: A Review. Canser Res. 1989, 49, 6449–6465. [Google Scholar]
- De Milito, A.; Fais, S. Tumor acidity, chemoresistance and proton pump inhibitors. Futur. Oncol. 2005, 1, 779–786. [Google Scholar] [CrossRef]
- McCarty, M.F.; Whitaker, J. Manipulating Tumor Acidification as a Cancer Treatment Strategy. Altern. Med. Rev. 2010, 15, 264–272. [Google Scholar]
- Cleland, W.W. Dithiothreitol, a New Protective Reagent for SH Groups. Biochemistry 1964, 3, 480–482. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, M.S.; Khanahmadi, S. Optimizing the preparation conditions and characterization of a stable and recyclable cross-linked enzyme aggregate (CLEA)-protease. Bioresour. Bioprocess. 2016, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yusof, F.; Khanahmadi, S.; Amid, A.; Mahmod, S.S. Cocoa pod husk, a new source of hydrolase enzymes for preparation of cross-linked enzyme aggregate. Springerplus 2016, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Mochidzuki, K.; Kobayashi, S.; Hiraide, H.; Wang, X.; Cui, Z. Effect of bovine serum albumin (BSA) on enzymatic cellulose hydrolysis. Appl. Biochem. Biotechnol. 2013, 170, 541–551. [Google Scholar] [CrossRef]
- Gaus, H.J.; Gupta, R.; Chappell, A.E.; Ostergaard, M.E.; Swayze, E.E.; Seth, P.P. Characterization of the Interactions of Chemically-Modified Therapeutic Nucleic Acids With Plasma Proteins Using a Fluorescence Polarization Assay. Nucleic Acids Res. 2019, 47, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, A.; Edwardson, T.G.W.; Hancock, M.A.; Dore, M.D.; Sleiman, H.F. Development of DNA Nanostructures for High-Affinity Binding to Human Serum Albumin. J. Am. Chem. Soc. 2017, 139, 7355–7362. [Google Scholar] [CrossRef] [PubMed]
- Gebrezgiabher, M.; Zalloum, W.A.; Clarke, D.J.; Miles, S.M.; Fedorova, A.A.; Zenkova, M.A.; Bichenkova, E.V. RNA knockdown by synthetic peptidyl-oligonucleotide ribonucleases: behavior of recognition and cleavage elements under physiological conditions. J. Biomol. Struct. Dyn. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalev, N.A.; Medvedeva, D.A.; Zenkova, M.A.; Vlassov, V.V. Cleavage of RNA by an amphiphilic compound lacking traditional catalytic groups. Bioorg. Chem. 2008, 36, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.S.; Meschaninova, M.I.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. 2’-O-methyl-modified anti-MDR1 fork-siRNA duplexes exhibiting high nuclease resistance and prolonged silencing activity. Oligonucleotides 2010, 20, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Corey, D.R. Chemistry, mechanism and clinical status of antisense oligonucleotides and duplex RNAs. Nucleic Acids Res. 2018, 46, 1584–1600. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of microRNA-target recognition. PLoS Biol. 2005, 3, 0404–0418. [Google Scholar] [CrossRef]
- Grimson, A.; Farh, K.K.H.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA Targeting Specificity in Mammals: Determinants beyond Seed Pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Robertson, B.; Dalby, A.B.; Karpilow, J.; Khvorova, A.; Leake, D.; Vermeulen, A. Specificity and functionality of microRNA inhibitors. Silence 2010, 1. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Juranek, S.; Li, H.; Sheng, G.; Wardle, G.S.; Tuschl, T.; Patel, D.J. Nucleation, propagation and cleavage of target RNAs in Ago silencing complexes. Nature 2009, 461, 754–761. [Google Scholar] [CrossRef]
- Hughes, R.A.; Miklos, A.E.; Ellington, A.D. Gene synthesis: Methods and applications. Methods Enzymol. 2011, 498, 277–309. [Google Scholar] [CrossRef]
- Caruthers, M.H. A brief review of DNA and RNA chemical synthesis. Biochem. Soc. Trans. 2011, 39, 575–580. [Google Scholar] [CrossRef]
- Beaucage, S.L.; Iyer, R.P. Advances in the Synthesis of Oligonucleotides by the Phosphoramidite Approach. Tetrahedron 1992, 48, 2223–2311. [Google Scholar] [CrossRef] [Green Version]
- Silberklang, M.; Gillum, A.M.; RajBhandary, U.L. Use of in vitro 32P labeling in the sequence analysis of nonradioactive tRNAs. Methods Enzymol. 1979, 59, 58–109. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.V.; Vlassov, V.V.; Giege, R. RNA Hydrolysis Catalyzed by Imidazole as a Reaction for Studying the Secondary Structure of RNA and Complexes of RNA With Oligonucleotides. Dokl. Akad. Nauk. 1996, 349, 411–413. [Google Scholar]
- Donis-Keller, H.; Maxam, A.M.; Gilbert, W. Mapping adenines, guanines, andpyrimidines in RNA. Nucleic Acids Res. 1977, 4, 2528–2538. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conjugate | Sequence 5′→3′ |
|---|---|
| DC | |
| miRNA-21-DC | TCAACATC–linker 1–[GRLRL]2–linker 2–GATAAGCTA |
| miRNA-18a-DC | TATCTGC–linker 1–[GRLRL]2–linker 2–ATGCACCTTA |
| miRNA-17-DC | CTACCTGC–linker 1–[GRLRL]2–linker 2–AAGCACTTTG |
| miRNA-155-DC | ACCCCTAT–linker 1–[GRLRL]2–linker 2–TTAGCATTAA |
| SC | |
| miRNA-21-SC | [GRLRL]2–linker 2–GATAAGCTA |
| miRNA-18a-SC | [GRLRL]2–linker 2–ATGCACCTTA |
| miRNA-17-SC | [GRLRL]2–linker 2–AAGCACTTTG |
| miRNA-155-SC | [GRLRL]2–linker 2–TTAGCATTAA |
| ON | |
| miRNA-21-ON | TCAACATC–TEG–TEG–GATAAGCTA |
| miRNA-18a-ON | TATCTGC–TEG–TEG–ATGCACCTTA |
| miRNA-17-ON | CTACCTGC–TEG–TEG–AAGCACTTTG |
| miRNA-155-ON | ACCCCTAT–TEG–TEG–TTAGCATTAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patutina, O.; Chiglintseva, D.; Bichenkova, E.; Gaponova, S.; Mironova, N.; Vlassov, V.; Zenkova, M. Dual miRNases for Triple Incision of miRNA Target: Design Concept and Catalytic Performance. Molecules 2020, 25, 2459. https://doi.org/10.3390/molecules25102459
Patutina O, Chiglintseva D, Bichenkova E, Gaponova S, Mironova N, Vlassov V, Zenkova M. Dual miRNases for Triple Incision of miRNA Target: Design Concept and Catalytic Performance. Molecules. 2020; 25(10):2459. https://doi.org/10.3390/molecules25102459
Chicago/Turabian StylePatutina, Olga, Daria Chiglintseva, Elena Bichenkova, Svetlana Gaponova, Nadezhda Mironova, Valentin Vlassov, and Marina Zenkova. 2020. "Dual miRNases for Triple Incision of miRNA Target: Design Concept and Catalytic Performance" Molecules 25, no. 10: 2459. https://doi.org/10.3390/molecules25102459