Inhibitory Effects of AF-343, a Mixture of Cassia tora L., Ulmus pumila L., and Taraxacum officinale, on Compound 48/80-Mediated Allergic Responses in RBL-2H3 Cells

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
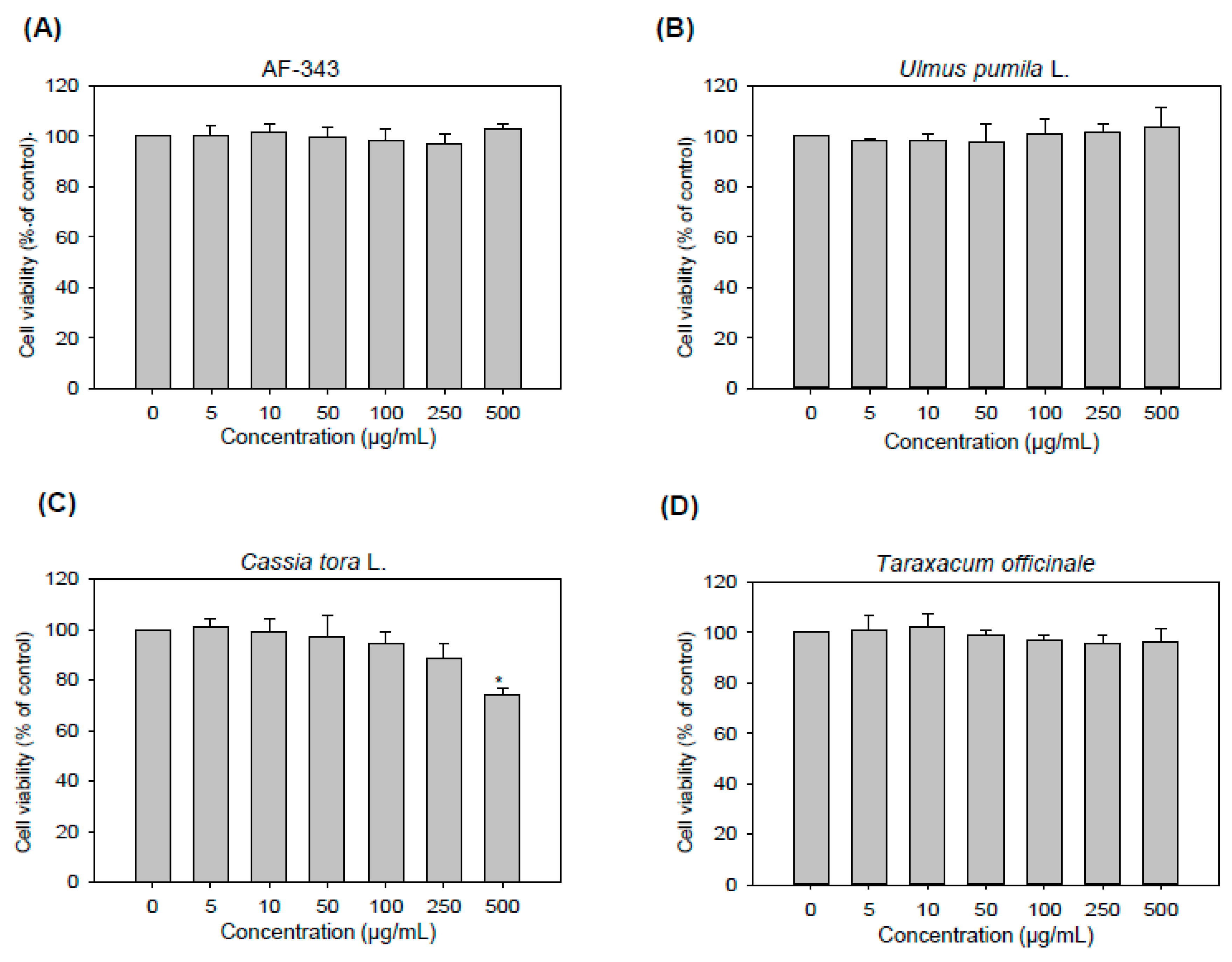
2.1. Cytotoxicity of AF-343 toward RBL-2H3 Cells
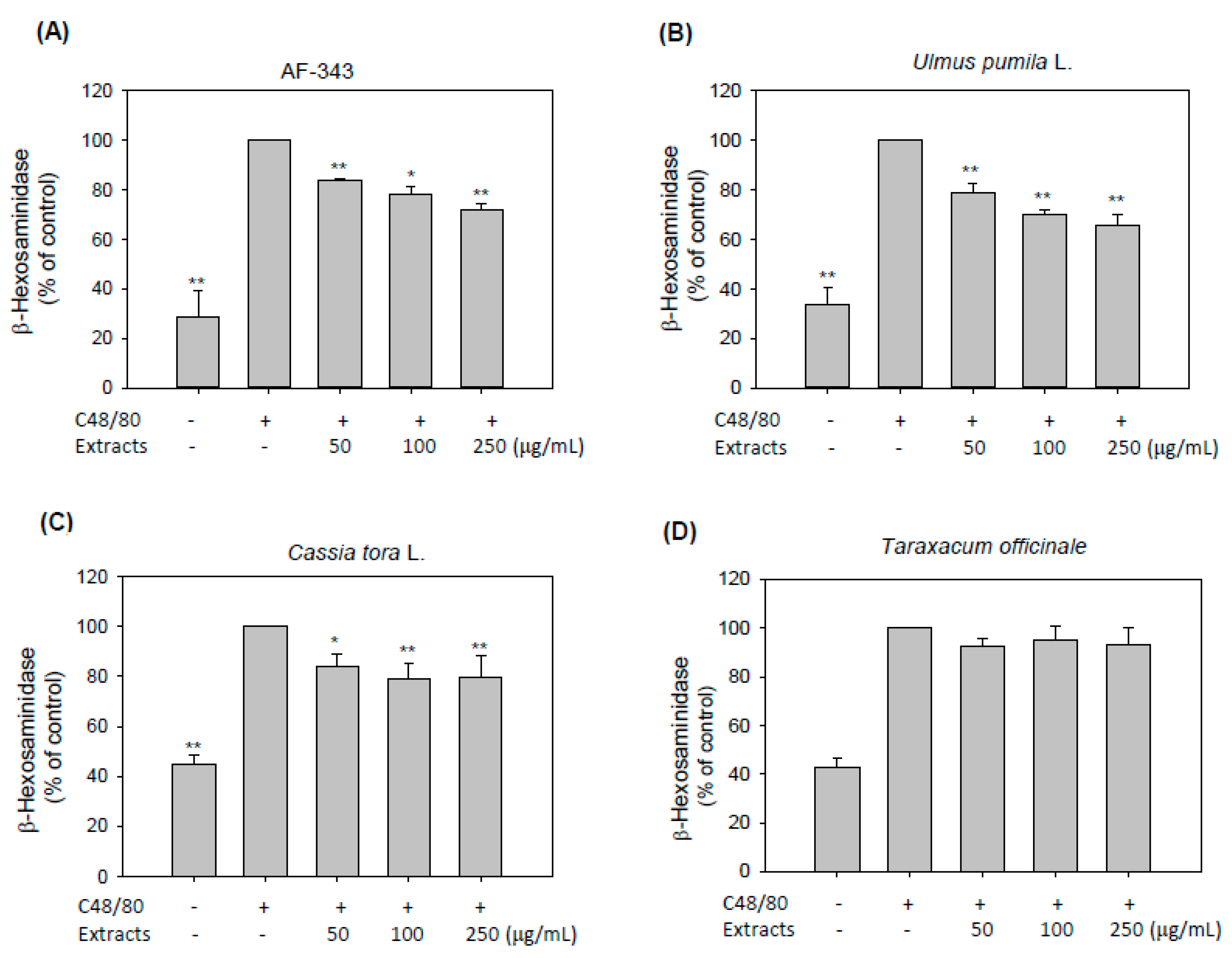
2.2. Inhibitory Effects of AF-343 on Compound 48/80-Induced Degranulation in RBL-2H3 Cells
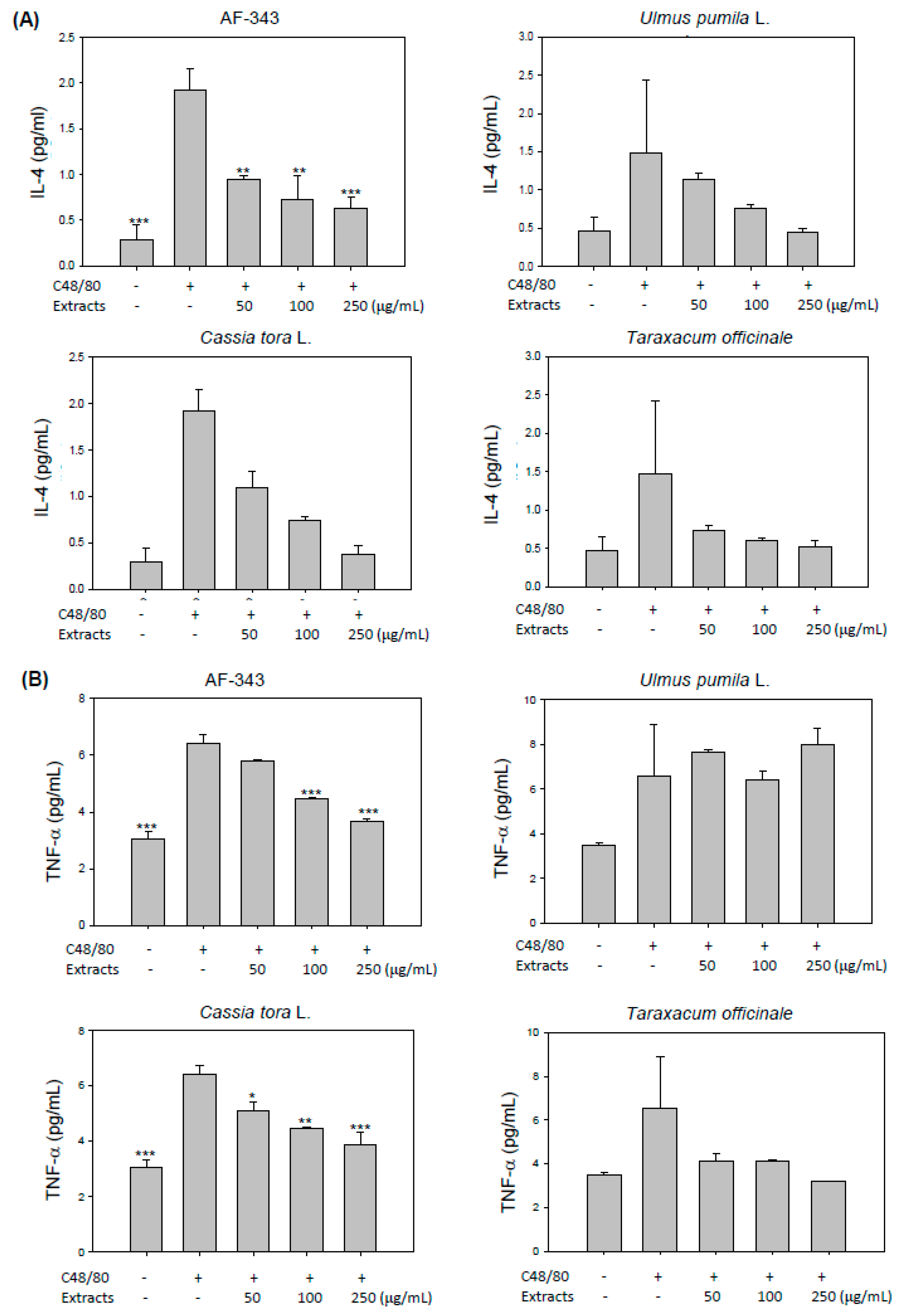
2.3. Ameliorative Effects of AF-343 on Compound 48/80-Mediated Cytokine Production in RBL-2H3 Cells
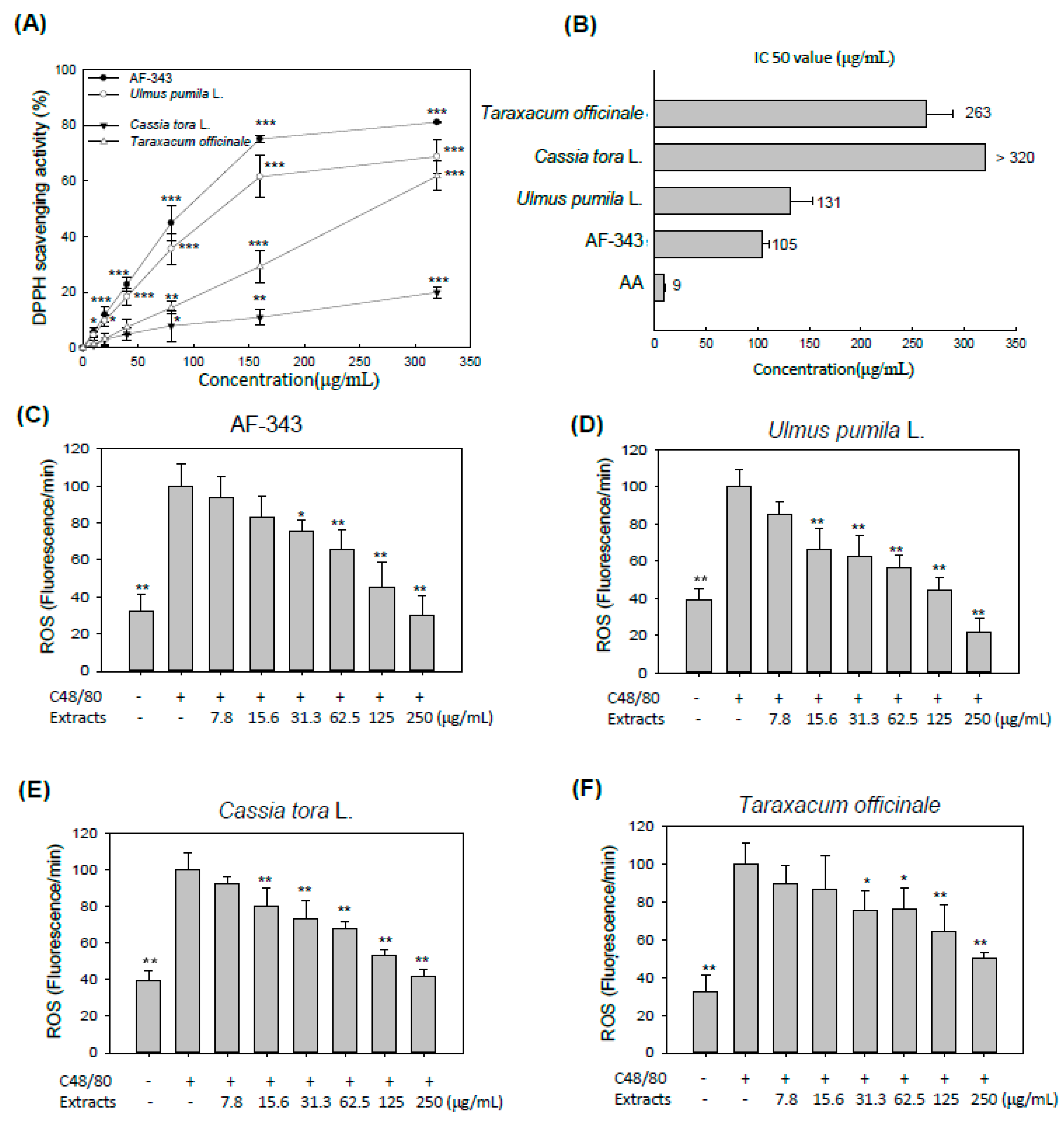
2.4. Antioxidant Activity of AF-343 In Vitro and in a Cell Culture System
3. Conclusions
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Cell Line and Culture Conditions
4.3. Cell Viability Assay
4.4. Cell Degranulation
4.5. Determination of Proinflammatory Cytokines Levels
4.6. DPPH Scavenging Activity
4.7. Measurement of Intracellular ROS Generation
4.8. Sample Preparation for Chromatography
4.9. HPLC–UV Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Averbeck, M.; Gebhardt, C.; Emmrich, F.; Treudler, R.; Simon, J.C. Immunologic principles of allergic disease. J. Der Dtsch. Dermatol. Ges. J. Ger. Soc. Dermatol. Jddg 2007, 5, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Licari, A.; Ciprandi, G.; Marseglia, A.; Castagnoli, R.; Barberi, S.; Caimmi, S.; Marseglia, G.L. Current recommendations and emerging options for the treatment of allergic rhinitis. Expert Rev. Clin. Immunol. 2014, 10, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K.; Iwata, A.; Tamachi, T.; Nakajima, H. Allergic airway inflammation: Key players beyond the Th2 cell pathway. Immunol. Rev. 2017, 278, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.R.; Wala, S.M. Inflammation, allergy and asthma, complex immune origin diseases: Mechanisms and therapeutic agents. Recent Pat. Inflamm. Allergy Drug Discov. 2013, 7, 62–95. [Google Scholar] [CrossRef]
- Verbsky, J.W.; McAllister, P.K.; Malone, D.G. Mast cell activation in human synovium explants by calcium ionophore A23187, compound 48/80, and rabbit IgG anti-human IgE, but not morphine sulfate. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 1996, 45, 35–41. [Google Scholar] [CrossRef]
- Koch, G.; Habermann, B.; Mohr, C.; Just, I.; Aktories, K. ADP-ribosylation of rho proteins is inhibited by melittin, mast cell degranulating peptide and compound 48/80. Eur. J. Pharmacol. 1992, 226, 87–91. [Google Scholar] [CrossRef]
- Takeuchi, T.; Harada, Y.; Moriyama, S.; Furuta, K.; Tanaka, S.; Miyaji, T.; Omote, H.; Moriyama, Y.; Hiasa, M. Vesicular Polyamine Transporter Mediates Vesicular Storage and Release of Polyamine from Mast Cells. J. Biol. Chem. 2017, 292, 3909–3918. [Google Scholar] [CrossRef] [Green Version]
- Passante, E.; Frankish, N. The RBL-2H3 cell line: Its provenance and suitability as a model for the mast cell. Inflamm. Res. 2009, 58, 737–745. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, S.H.; Jhee, K.H.; Yang, S.A. Tricin Isolated from Enzyme-Treated Zizania latifolia Extract Inhibits IgE-Mediated Allergic Reactions in RBL-2H3 Cells by Targeting the Lyn/Syk Pathway. Molecules 2020, 25, 2084. [Google Scholar] [CrossRef]
- Alkanfari, I.; Freeman, K.B.; Roy, S.; Jahan, T.; Scott, R.W.; Ali, H. Small-Molecule Host-Defense Peptide Mimetic Antibacterial and Antifungal Agents Activate Human and Mouse Mast Cells via Mas-Related GPCRs. Cells 2019, 8, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeCorte, B.L. Underexplored Opportunities for Natural Products in Drug Discovery. J. Med. Chem. 2016, 59, 9295–9304. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.W.; Jeong, Y.S.; Han, J.; Chun, Y.J.; Kim, H.K.; Kim, M.Y.; Kim, B.J.; Park, K.M.; Kim, J.K.; Kim, J.H.; et al. Skin Hydration and Collagen Synthesis of AF-343 in HS68 Cell Line and NC/Nga Mice by Filaggrin Expression and Suppression of Matrix Metallopreteinase. Toxicol. Res. 2011, 27, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Park, H.J.; Seo, E.K.; Oh, J.Y.; Ji, S.Y.; Park, D.K.; Lim, Y. Effects of flavone derivatives on antigen-stimulated degranulation in RBL-2H3 cells. Chem. Biol. Drug Des. 2013, 81, 228–237. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Graham, A.C.; Temple, R.M.; Obar, J.J. Mast cells and influenza a virus: Association with allergic responses and beyond. Front. Immunol. 2015, 6, 238. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, N.; Wang, C.C.; Brochetta, C.; Ke, G.; Vita, F.; Qi, Z.; Rivera, J.; Soranzo, M.R.; Zabucchi, G.; Hong, W.; et al. VAMP-8 segregates mast cell-preformed mediator exocytosis from cytokine trafficking pathways. Blood 2008, 111, 3665–3674. [Google Scholar] [CrossRef] [Green Version]
- Hide, I.; Toriu, N.; Nuibe, T.; Inoue, A.; Hide, M.; Yamamoto, S.; Nakata, Y. Suppression of TNF-alpha secretion by azelastine in a rat mast (RBL-2H3) cell line: Evidence for differential regulation of TNF-alpha release, transcription, and degranulation. J. Immunol. 1997, 159, 2932–2940. [Google Scholar]
- Bowler, R.P.; Crapo, J.D. Oxidative stress in allergic respiratory diseases. J. Allergy Clin. Immunol. 2002, 110, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Wolfreys, K.; Oliveira, D.B. Alterations in intracellular reactive oxygen species generation and redox potential modulate mast cell function. Eur. J. Immunol. 1997, 27, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Karahan, F.; Kulak, M.; Urlu, E.; Gozuacik, H.G.; Boyumez, T.; Sekeroglu, N.; Doganturk, I.H. Total phenolic content, ferric reducing and DPPH scavenging activity of Arum dioscoridis. Nat. Prod. Res. 2015, 29, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Patro, B.S.; Bauri, A.K.; Mishra, S.; Chattopadhyay, S. Antioxidant activity of Myristica malabarica extracts and their constituents. J. Agric. Food Chem. 2005, 53, 6912–6918. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, D.; Asaduzzaman, M.; Young, F. Real time monitoring and quantification of reactive oxygen species in breast cancer cell line MCF-7 by 2’,7’-dichlorofluorescin diacetate (DCFDA) assay. J. Pharmacol. Toxicol. Methods 2018, 94, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134 (Suppl. 12), 3479S–3485S. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, I.; Kobayashi, M.; Hamada, T.; Tsuda, K.; Goto, H.; Imaizumi, K.; Nozawa, A.; Sugimoto, A.; Kakuda, T. Heat-epimerized tea catechins rich in gallocatechin gallate and catechin gallate are more effective to inhibit cholesterol absorption than tea catechins rich in epigallocatechin gallate and epicatechin gallate. J. Agric. Food Chem. 2003, 51, 7303–7307. [Google Scholar] [CrossRef]
- Yang, H.H.; Son, J.K.; Jung, B.; Zheng, M.; Kim, J.R. Epifriedelanol from the root bark of Ulmus davidiana inhibits cellular senescence in human primary cells. Planta Med. 2011, 77, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Shaterzadeh-Yazdi, H.; Farkhondeh, T.; Samarghandian, S. An Overview on Cardiovascular Protective Effects of Catechins. Cardiovasc. Hematol. Disord. Drug Targets 2017, 17, 154–160. [Google Scholar] [CrossRef]
- Kim, M.; Lim, S.J.; Lee, H.J.; Nho, C.W. Cassia tora Seed Extract and Its Active Compound Aurantio-obtusin Inhibit Allergic Responses in IgE-Mediated Mast Cells and Anaphylactic Models. J. Agric. Food Chem. 2015, 63, 9037–9046. [Google Scholar] [CrossRef]
- Hou, J.; Gu, Y.; Zhao, S.; Huo, M.; Wang, S.; Zhang, Y.; Qiao, Y.; Li, X. Anti-Inflammatory Effects of Aurantio-Obtusin from Seed of Cassia obtusifolia L. through Modulation of the NF-kappaB Pathway. Molecules 2018, 23, 3093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Li, Q.; Lv, X.; Liao, L.; Yang, W.; Lu, P.; Zhu, D. Aurantio-obtusin relaxes systemic arteries through endothelial PI3K/AKT/eNOS-dependent signaling pathway in rats. J. Pharmacol. Sci. 2015, 128, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, D.; Kim, S.J.; Kim, H.S. Anti-inflammatory evaluation of the methanolic extract of Taraxacum officinale in LPS-stimulated human umbilical vein endothelial cells. BMC Complementary Altern. Med. 2017, 17, 508. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Meng, N.; Liu, Z.; Guo, L.; Dong, L.; Li, B.; Ye, Q. Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways. Nutrients 2018, 10, 926. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, M.; Poljacki, M.; Mimica-Dukic, N.; Boza, P.; Vujanovic, L.; Duran, V.; Stojanovic, S. Sesquiterpene lactone mix patch testing supplemented with dandelion extract in patients with allergic contact dermatitis, atopic dermatitis and non-allergic chronic inflammatory skin diseases. Contact Dermat. 2004, 51, 101–110. [Google Scholar] [CrossRef]
- Dearman, R.J.; Skinner, R.A.; Deakin, N.; Shaw, D.; Kimber, I. Evaluation of an in vitro method for the measurement of specific IgE antibody responses: The rat basophilic leukemia (RBL) cell assay. Toxicology 2005, 206, 195–205. [Google Scholar] [CrossRef]
- Hatano, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licorice root: Their relative astringency and radical scavenging effects. Chem. Pharm. Bull. 1988, 36, 2090–2097. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catechin, g/kg (mean ± SD) | Aurantio-Obtusin, g/kg (mean ± SD) | Chicoric Acid, g/kg (mean ± SD) | |
|---|---|---|---|
| AF-343 | 0.40 ± 0.01 | 1.60 ± 0.09 | 0.76 ± 0.01 |
| Ulmus pumila L. | 0.32 ± 0.02 | ND | ND |
| Cassia tora L. | ND | 0.49 ± 0.03 | ND |
| Taraxacum officinale | ND | ND | 11.19 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.K.; Song, J.; Seo, Y.; Koh, E.M.; Kim, S.-H.; Jung, K.J. Inhibitory Effects of AF-343, a Mixture of Cassia tora L., Ulmus pumila L., and Taraxacum officinale, on Compound 48/80-Mediated Allergic Responses in RBL-2H3 Cells. Molecules 2020, 25, 2434. https://doi.org/10.3390/molecules25102434
Lee EK, Song J, Seo Y, Koh EM, Kim S-H, Jung KJ. Inhibitory Effects of AF-343, a Mixture of Cassia tora L., Ulmus pumila L., and Taraxacum officinale, on Compound 48/80-Mediated Allergic Responses in RBL-2H3 Cells. Molecules. 2020; 25(10):2434. https://doi.org/10.3390/molecules25102434
Chicago/Turabian StyleLee, Eun Kyeong, Jeongah Song, Youjin Seo, Eun Mi Koh, Seon-Hee Kim, and Kyung Jin Jung. 2020. "Inhibitory Effects of AF-343, a Mixture of Cassia tora L., Ulmus pumila L., and Taraxacum officinale, on Compound 48/80-Mediated Allergic Responses in RBL-2H3 Cells" Molecules 25, no. 10: 2434. https://doi.org/10.3390/molecules25102434