
LeTetR Positively Regulates 3-Hydroxylation of the Antifungal HSAF and Its Analogs in Lysobacter enzymogenes OH11


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
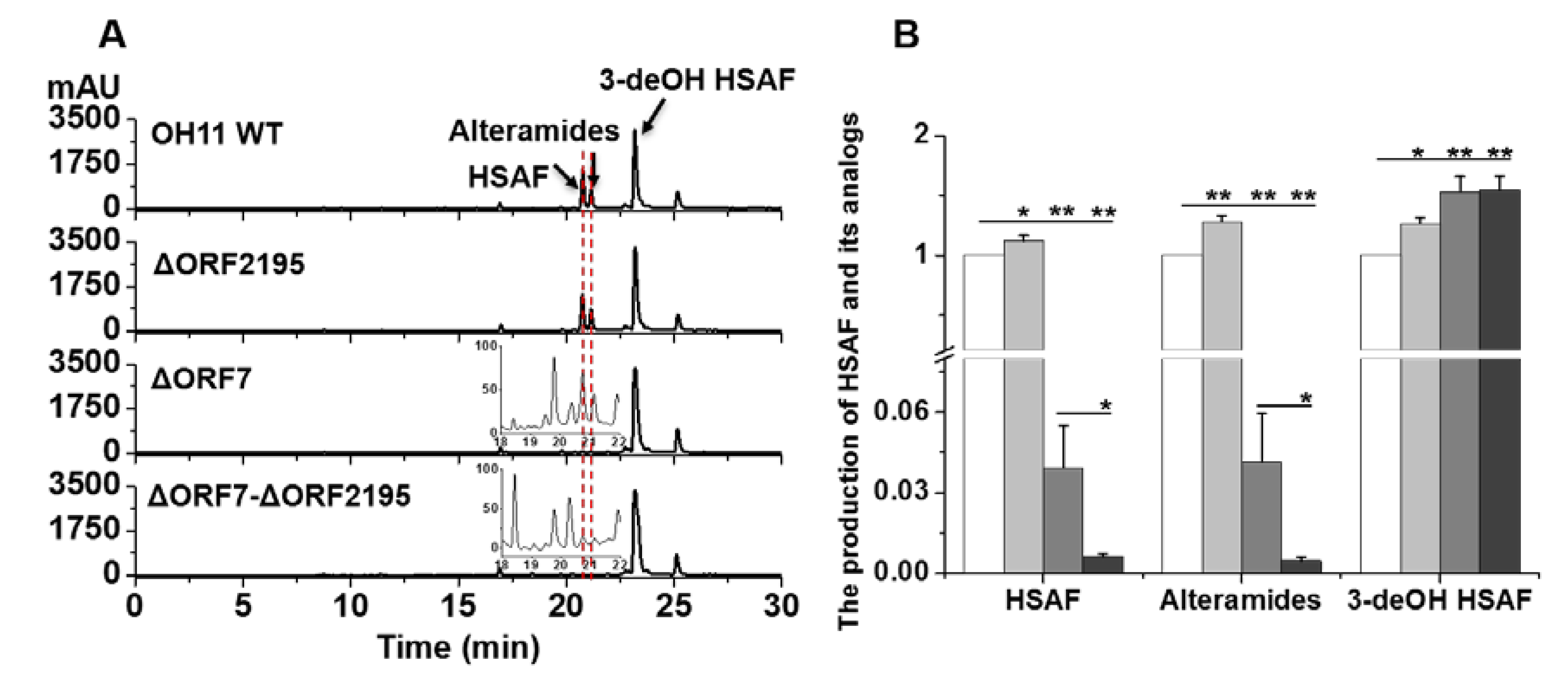
2.1. LeTetR Regulates the Production of HSAF and Alteramides
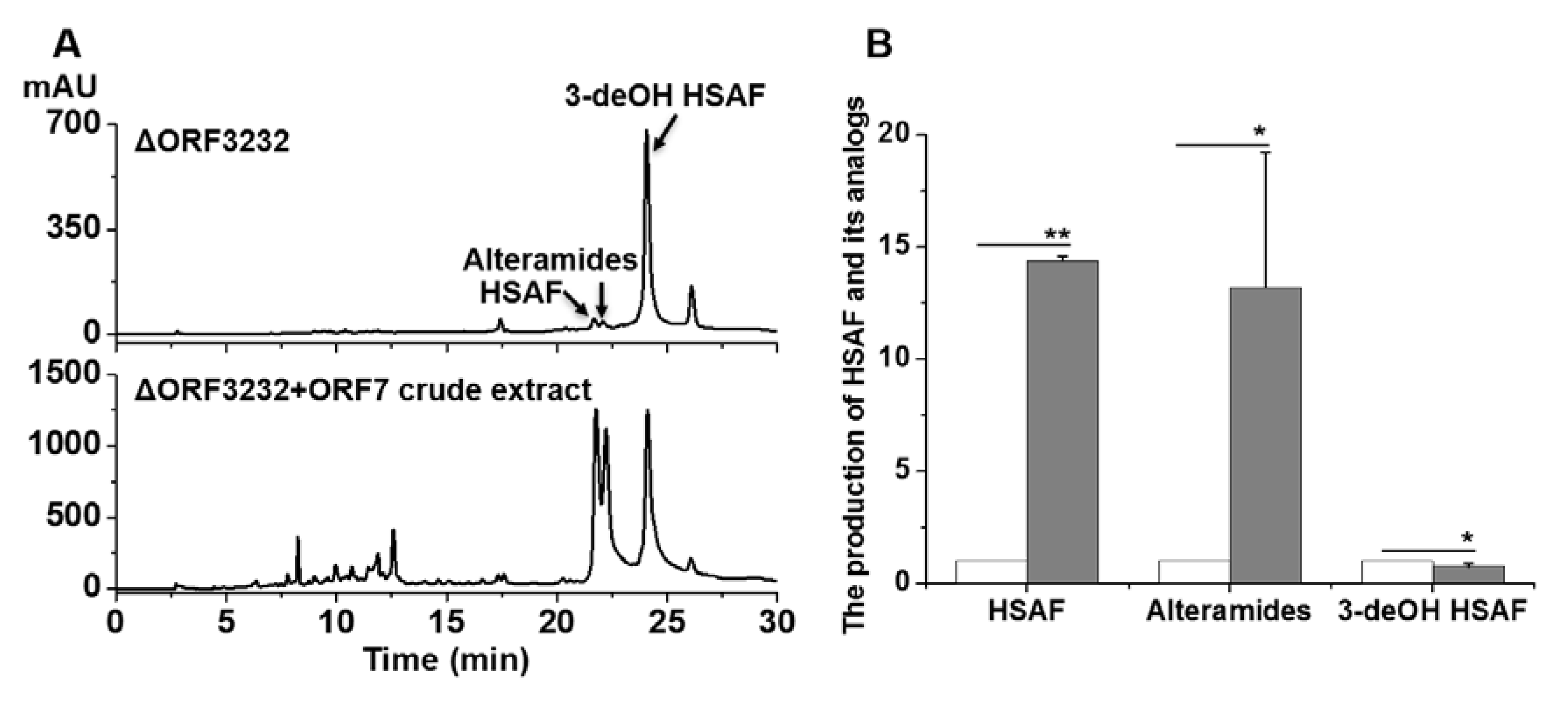
2.2. Bioconversion of 3-deOH Compounds in the LeTetR Mutant to HSAF and Alteramides Using an Enzyme Extract from the E. coli-Expressing orf7 Gene
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
4.2. DNA Manipulation and Lysobacter Transformation
4.3. Primers and PCR
4.4. Generation of the Deletion Mutant of orf3232 and Its Complementary Strain
4.5. Construction of the Deletion Mutant of Fatty Acid Hydroxylase Genes
4.6. RNA Extraction and Real-Time PCR
4.7. Extraction and HPLC Analysis of HSAF and Its Analogs
4.8. Preparation of Protein Crude Extract Containing ORF7
4.9. Exogenous Addition of the ORF7 Crude Extract to the ΔORF3232 Strain
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Christensen, P.; Cook, F.D. Lysobacter, a new genus of non-fruiting, gliding bacteria with a high base ratio. Int. J. Syst. Bacteriol. 1978, 28, 367–393. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wright, S.; Shen, Y.; Du, L. Bioactive natural products from Lysobacter. Nat. Prod. Rep. 2012, 19, 1277–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Zaleta-Rivera, K.; Zhu, X.; Huffman, J.; Millet, J.C.; Harris, S.D.; Yuen, G.; Li, X.C.; Du, L. Structure and biosynthesis of heat-stable antifungal factor (HSAF), a broad-spectrum antimycotic with a novel mode of action. Antimicrob. Agents Chemother. 2007, 51, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, L.; Qian, G.; Xie, Y.; Hang, J.; Chen, H.; Zaleta-Rivera, K.; Li, Y.; Shen, Y.; Dussault, P.H.; Liu, F.; et al. Biosynthesis of hsaf, a tetramic acid-containing macrolactam from Lysobacter enzymogenes. J. Am. Chem. Soc. 2011, 133, 643–645. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Du, L.; Yuen, G.; Harris, S.D. Distinct ceramide synthases regulate polarized growth in the filamentous fungus Aspergillus nidulans. Mol. Biol. Cell 2006, 17, 1218–1227. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Li, Z.; Li, Y.; Lu, C.; Wang, H.; Shen, Y.; Du, L. HSAF-induced antifungal effects in Candida albicans through ROS-mediated apoptosis. RSC Adv. 2016, 6, 30895–30904. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Li, Y.; Li, Z.; Zhang, J.; Lu, C.; Wang, H.; Shen, Y.; Du, L. Alteramide B is a microtubule antagonist of inhibiting Candida albicans. Biochim. Biophys. Acta 2016, 1860, 2097–2106. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wang, Y.; Tombosa, S.; Wright, S.; Huffman, J.; Yuen, G.; Qian, G.; Liu, F.; Shen, Y.; Du, L. Identification of a small molecule signaling factor that regulates the biosynthesis of the antifungal polycyclic tetramate macrolactam HSAF in Lysobacter enzymogenes. Appl. Microbiol. Biotechnol. 2015, 99, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Wang, Y.; Liu, Y.; Xu, F.; He, Y.W.; Du, L.; Venturi, V.; Fan, J.; Hu, B.; Liu, F. Lysobacter enzymogenes uses two distinct cell-cell signaling systems for differential regulation of secondary-metabolite biosynthesis and colony morphology. Appl. Environ. Microbiol. 2013, 79, 6604–6616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, Y.; Zhang, J.; Zhao, Y.; Shen, Y.; Su, Z.; Xu, G.; Du, L.; Huffman, J.M.; Venturi, V.; et al. Transcriptomic analysis reveals new regulatory roles of Clp signaling in secondary metabolite biosynthesis and surface motility in Lysobacter enzymogenes OH11. Appl. Microbiol. Biotechnol. 2014, 98, 9009–9020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Chen, H.; Wang, P.; Tombosa, S.; Du, L.; Han, Y.; Shen, Y.; Qian, G.; Liu, F. 4-Hydroxybenzoic acid is a diffusible factor that connects metabolic shikimate pathway to the biosynthesis of a unique antifungal metabolite in Lysobacter enzymogenes. Mol. Microbiol. 2017, 104, 163–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Han, S.; Fu, Z.Q.; Qian, G.; Liu, F. Heat-stable antifungal factor (HSAF) biosynthesis in Lysobacter enzymogenes is controlled by the interplay of two transcription factors and a diffusible molecule. Appl. Environ. Microbiol. 2018, 84, e01754-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Chen, H.; Qian, G.; Liu, F. LetR is a TetR family transcription factor from Lysobacter controlling antifungal antibiotic biosynthesis. Appl. Microbiol. Biotechnol. 2017, 101, 3273–3282. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Qian, G.; Chen, Y.; Du, L.; Liu, F.; Yuen, G.Y. PilG is involved in the regulation of twitching motility and antifungal antibiotic biosynthesis in the biological control agent Lysobacter enzymogenes. Phytopathology 2015, 105, 1318–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Xia, J.; Su, Z.; Xu, G.; Gomelsky, M.; Qian, G.; Liu, F. Lysobacter PilR, the regulator of type IV pilus synthesis, controls antifungal antibiotic production via a cyclic di-GMP pathway. Appl. Environ. Microbiol. 2017, 83, e03397-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Han, S.; Huo, C.; Chin, K.H.; Chou, S.H.; Gomelsky, M.; Qian, G.; Liu, F. Signaling specificity in the c-di-GMP-dependent network regulating antibiotic synthesis in Lysobacter. Nucl. Acids Res. 2018, 46, 9276–9288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yu, L.J.; Li, F.Q.; Du, L. Spermidine-regulated biosynthesis of heat-stable antifungal factor (HSAF) in Lysobacter enzymogenes OH11. Front. Microbiol. 2018, 9, 2984. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, T.T.; Ning, Y.; Shen, D.Y.; Yang, N.D.; Li, Y.; Chou, S.H.; Yang, L.; Qian, G. Spermidine plays a significant role in stabilizing a master transcription factor Clp to promote antifungal activity in Lysobacter enzymogenes. Appl. Microbiol. Biotechnol. 2019, 103, 1811–1822. [Google Scholar] [CrossRef]
- Li, Y.; Huffman, J.; Li, Y.; Du, L.; Shen, Y. 3-Hydroxylation of the polycyclic tetramate macrolactam in the biosynthesis of antifungal HSAF from Lysobacter enzymogenes C3. MedChemComm 2012, 9, 982–986. [Google Scholar] [CrossRef]
- Ramos, J.L.; Martinez-Bueno, M.; Molina-Henares, A.J.; Teran, W.; Watanabe, K.; Zhang, X.D.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–358. [Google Scholar] [CrossRef] [Green Version]
- Quandt, J.; Hynes, M.F. Versatile suicide vectors which allow direct selection for gene replacement in Gram-negative bacteria. Gene 1993, 127, 15–21. [Google Scholar] [CrossRef]
- Liu, G.; Tian, Y.Q.; Yang, H.H.; Tan, H.R. A pathway-specific transcriptional regulatory gene for nikkomycin biosynthesis in Streptomyces ansochromogenes that also influences colony development. Mol. Microbiol. 2005, 55, 1855–1866. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(t)(-delta delta c) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds HSAF are available from the authors. |




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Khetrapal, V.; Liu, F.; Du, L. LeTetR Positively Regulates 3-Hydroxylation of the Antifungal HSAF and Its Analogs in Lysobacter enzymogenes OH11. Molecules 2020, 25, 2286. https://doi.org/10.3390/molecules25102286
Yu L, Khetrapal V, Liu F, Du L. LeTetR Positively Regulates 3-Hydroxylation of the Antifungal HSAF and Its Analogs in Lysobacter enzymogenes OH11. Molecules. 2020; 25(10):2286. https://doi.org/10.3390/molecules25102286
Chicago/Turabian StyleYu, Lingjun, Vimmy Khetrapal, Fengquan Liu, and Liangcheng Du. 2020. "LeTetR Positively Regulates 3-Hydroxylation of the Antifungal HSAF and Its Analogs in Lysobacter enzymogenes OH11" Molecules 25, no. 10: 2286. https://doi.org/10.3390/molecules25102286