
Dietary Compounds Influencing the Sensorial, Volatile and Phytochemical Properties of Bovine Milk


 , ,
, , 

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Microbial Analyses
2.2. Pasteurized Milk Compositions
2.3. Free Fatty Acid Analyses
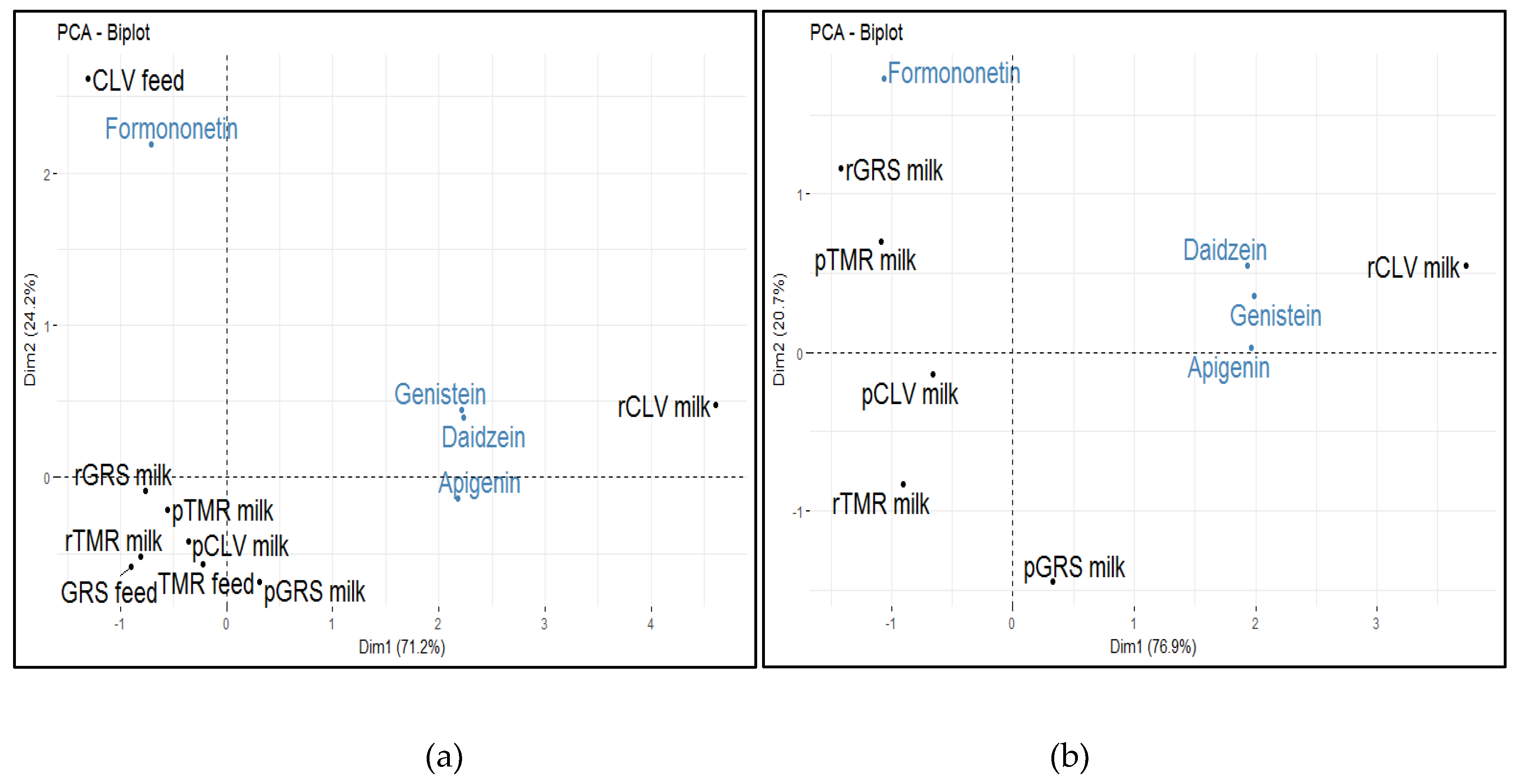
2.4. Phytochemical Analyses
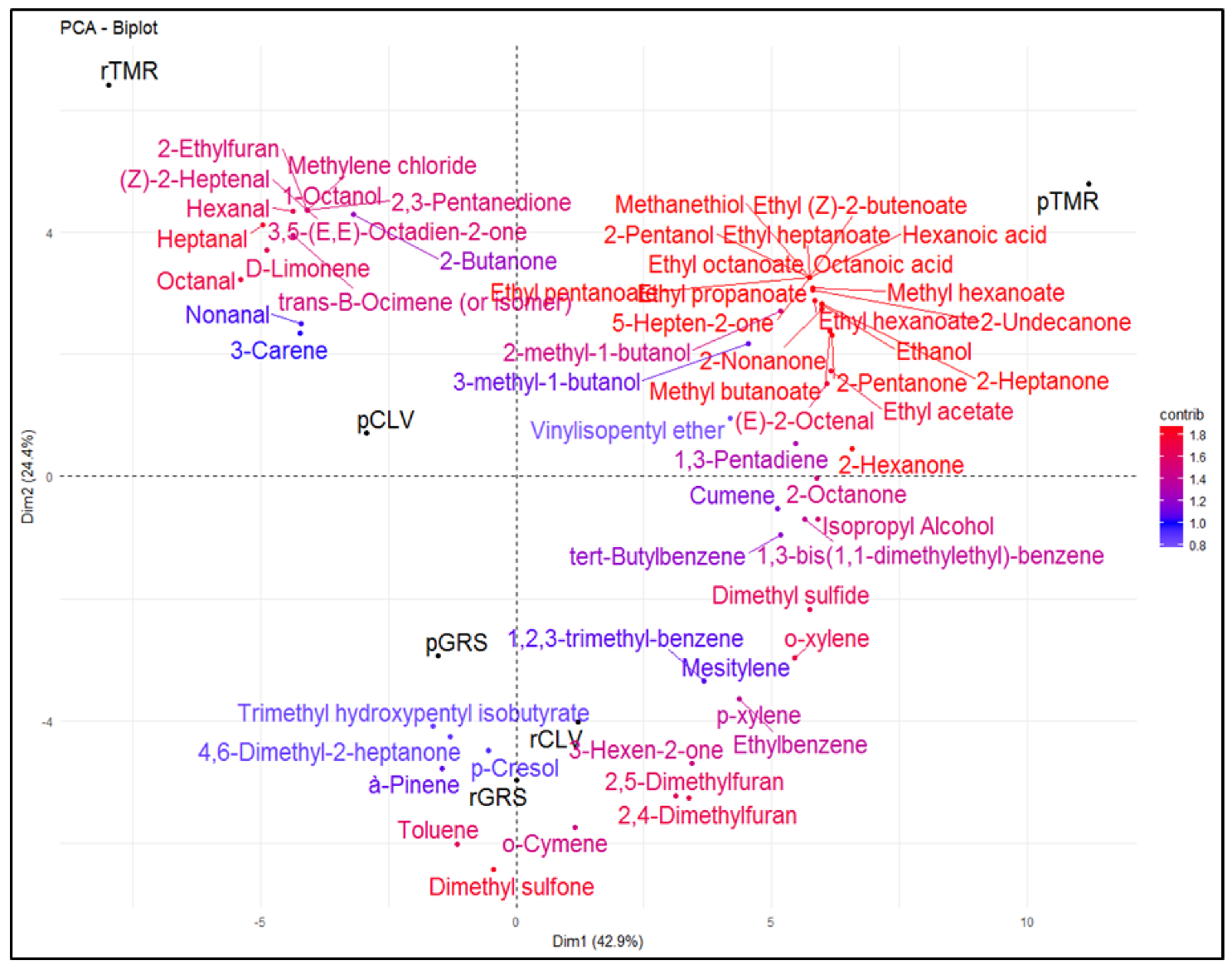
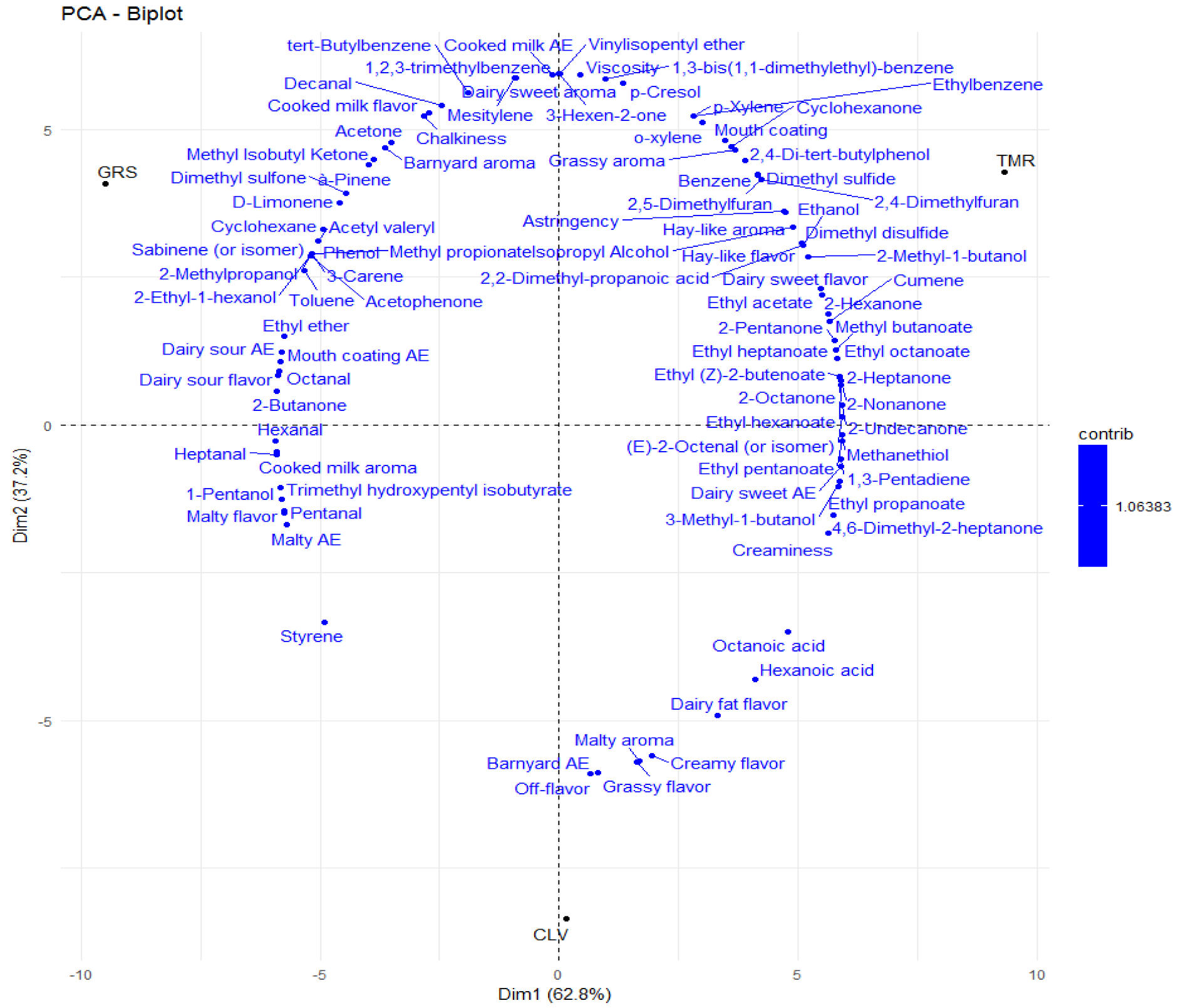
2.5. Volatile Analyses (Feed, Raw and Pasteurized Milk)
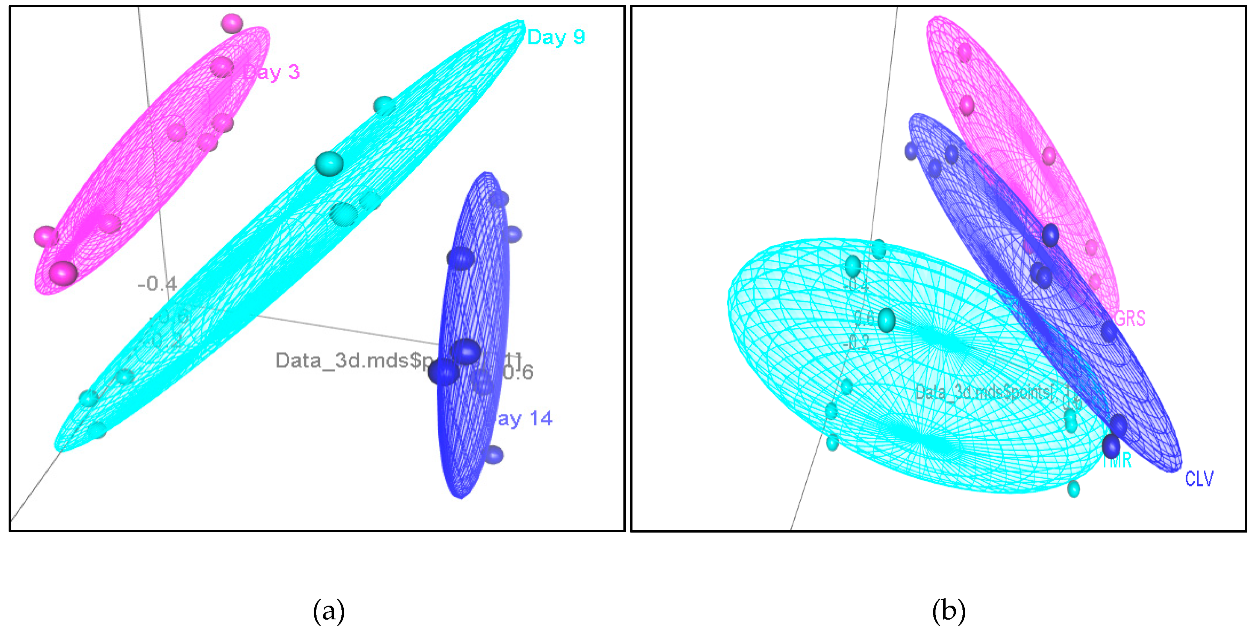
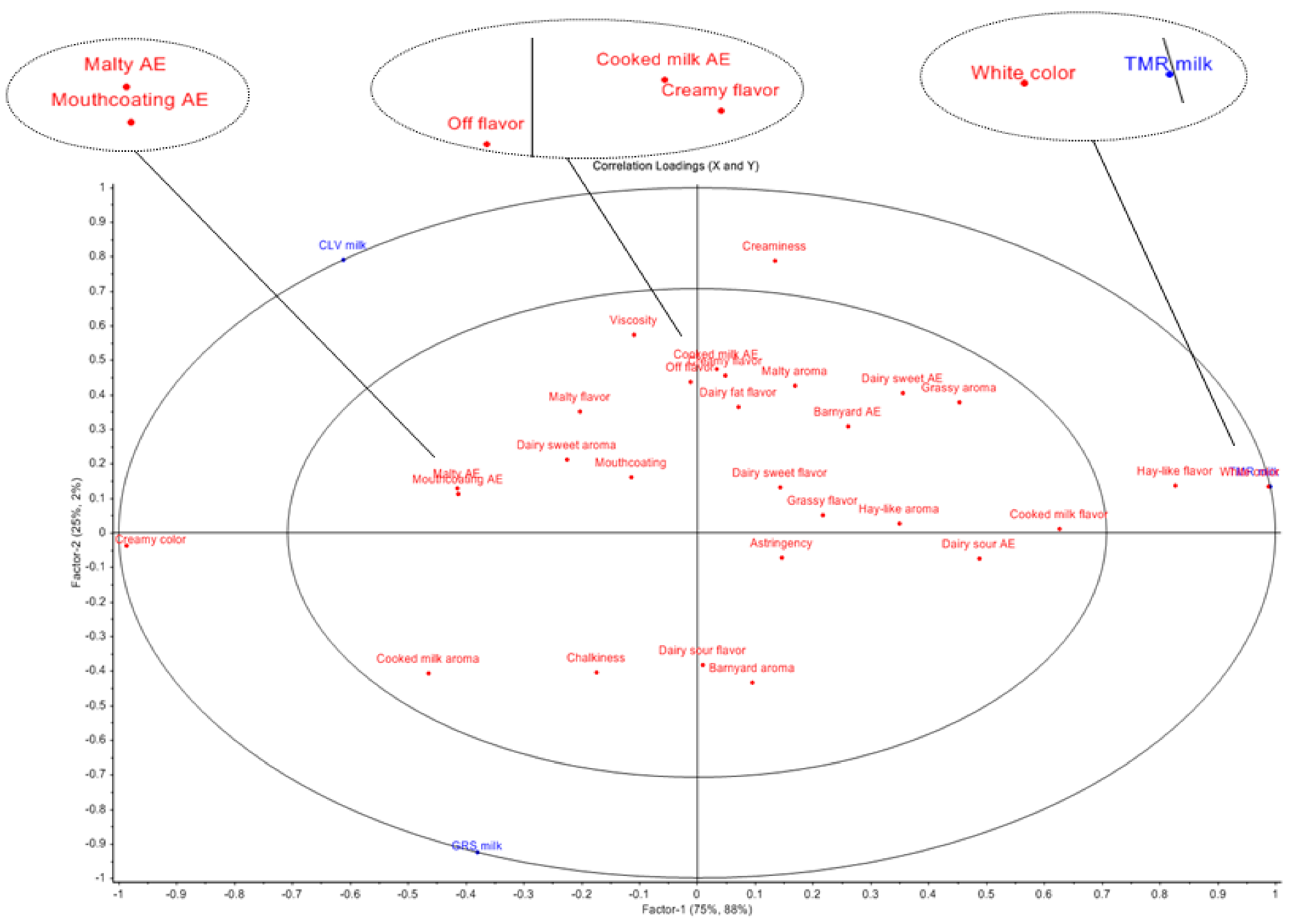
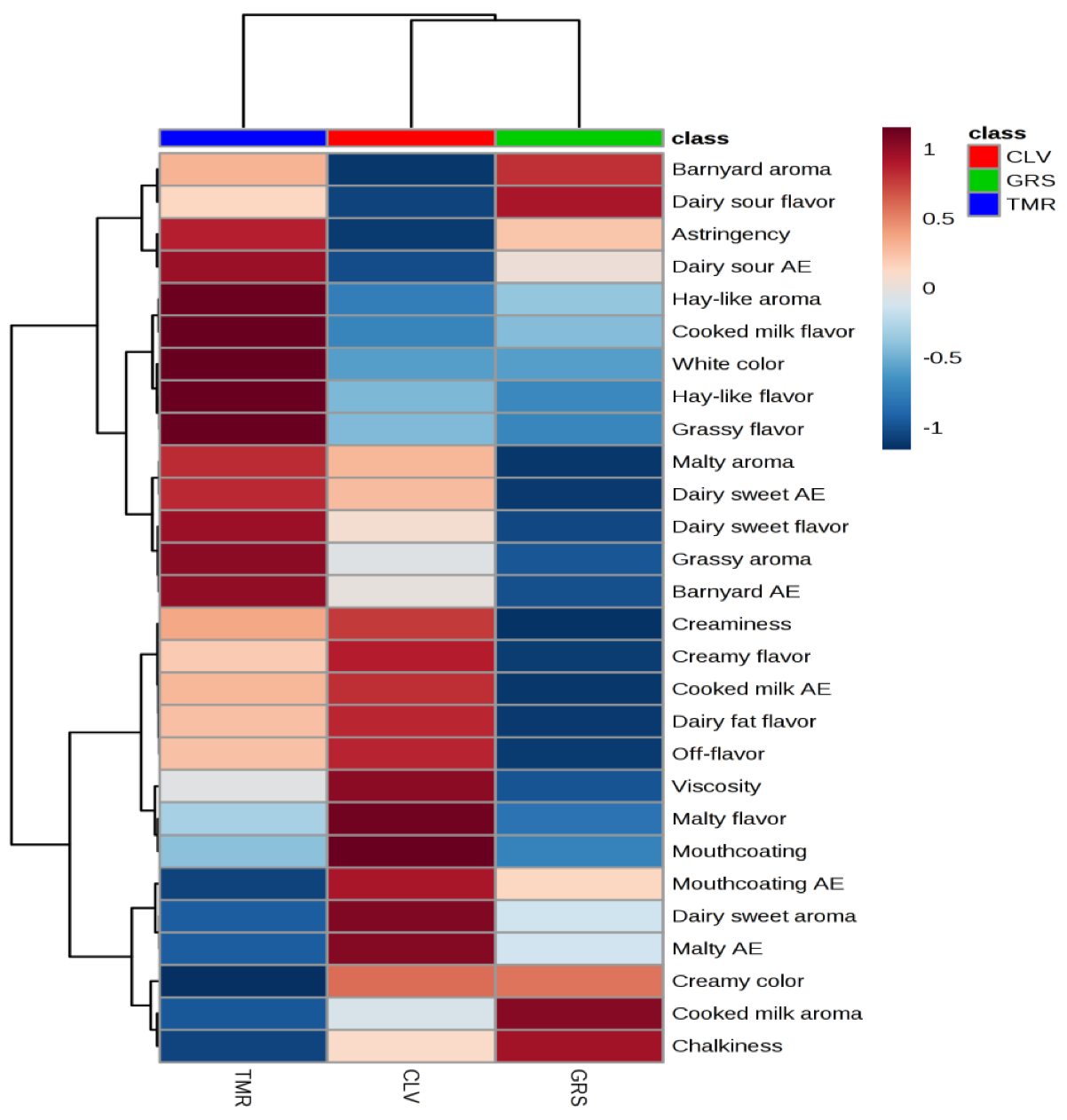
2.6. Sensory Analyses of Pasteurized Milk Samples
3. Materials and Methods
3.1. Feed Samples
3.2. Milk Samples and Processing
3.3. Microbial Analyses
3.4. Raw and Pasteurized Milk Compositions
3.5. Free Fatty Acid Analyses
3.6. Phytochemical Extraction and Analyses
3.7. Volatile Analyses
3.8. Sensory Analyses
3.9. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faulkner, H.; O’Callaghan, T.F.; McAuliffe, S.; Hennessy, D.; Stanton, C.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Effect of different forage types on the volatile and sensory properties of bovine milk. J. Dairy Sci. 2018, 101, 1034–1047. [Google Scholar] [CrossRef]
- Wang, C.; Liu, J.; Duan, B.; Lao, Y.; Qi, P.X.; Ren, D. Effects of dietary antioxidant supplementation of feed, milk processing and storage on the lutein content and sensory quality of bovine milk. Int. J. Dairy Technol. 2018, 71, 849–856. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev. 2004, 44, 467–492. [Google Scholar] [CrossRef] [PubMed]
- Coppa, M.; Martin, B.; Pradel, P.; Leotta, B.; Priolo, A.; Vasta, V. Effect of a hay-based diet or different upland grazing systems on milk volatile compounds. J. Agric. Food Chem. 2011, 59, 4947–4954. [Google Scholar] [CrossRef]
- Kilcawley, K.N.; Faulkner, H.; Clarke, H.J.; O’Sullivan, M.G.; Kerry, J.P. Factors Influencing the Flavour of Bovine Milk and Cheese from Grass Based versus Non-Grass Based Milk Production Systems. Foods 2018, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmquist, D.; Beaulieu, A.D.; Barbano, D. Feed and animal factors influencing milk fat composition. J. Dairy Sci. 1993, 76, 1753–1771. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Doreau, M. Effect of different types of forages, animal fat or marine oils in cow’s diet on milk fat secretion and composition, especially conjugated linoleic acid (CLA) and polyunsaturated fatty acids. Livest. Prod. Sci. 2001, 70, 31–48. [Google Scholar] [CrossRef]
- Caroprese, M.; Mancino, R.; Ciliberti, M.G.; Di Luccia, A.; La Gatta, B.; Albenzio, M. Fatty acid profile and coagulating ability of milk from Jersey and Friesian cows fed whole flaxseed. J. Dairy Res. 2017, 84, 14–22. [Google Scholar] [CrossRef]
- Bodkowski, R.; Czyż, K.; Kupczyński, R.; Patkowska-Sokoła, B.; Nowakowski, P.; Wiliczkiewicz, A. Lipid complex effect on fatty acid profile and chemical composition of cow milk and cheese. J. Dairy Sci. 2016, 99, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Danes, M.D.A.C.; Chagas, L.; Pedroso, A.; Santos, F.A.P. Effect of protein supplementation on milk production and metabolism of dairy cows grazing tropical grass. J. Dairy Sci. 2013, 96, 407–419. [Google Scholar] [CrossRef]
- Schöne, F.; Spörl, K.; Leiterer, M. Iodine in the feed of cows and in the milk with a view to the consumer’s iodine supply. J. Trace Elem. Med. Biol. 2017, 39, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Glover, K.; Budge, S.; Rose, M.; Rupasinghe, H.; MacLaren, L.; Green-Johnson, J.; Fredeen, A. Effect of feeding fresh forage and marine algae on the fatty acid composition and oxidation of milk and butter. J. Dairy Sci. 2012, 95, 2797–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lejonklev, J.; Kidmose, U.; Jensen, S.; Petersen, M.A.; Helwing, A.; Mortensen, G.; Weisbjerg, M.R.; Larsen, M.K. Effect of oregano and caraway essential oils on the production and flavor of cow milk. J. Dairy Sci. 2016, 99, 7898–7903. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ferreira, G.; Teets, C.; Corl, B.; Thomason, W.; Griffey, C. Effects of feeding hull-less barley on production performance, milk fatty acid composition, and nutrient digestibility of lactating dairy cows. J. Dairy Sci. 2017, 100, 3576–3583. [Google Scholar] [CrossRef]
- AbuGhazaleh, A.; Holmes, L. Diet supplementation with fish oil and sunflower oil to increase conjugated linoleic acid levels in milk fat of partially grazing dairy cows. J. Dairy Sci. 2007, 90, 2897–2904. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Mannion, D.; Apopei, D.; McCarthy, N.A.; Hogan, S.A.; Kilcawley, K.N.; Egan, M. Influence of Supplemental Feed Choice for Pasture-Based Cows on the Fatty Acid and Volatile Profile of Milk. Foods 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Paludetti, L.; Jordan, K.; Kelly, A.; Gleeson, D. Evaluating the effect of storage conditions on milk microbiological quality and composition. Irish J. Agric. Food Res. 2018, 57, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Gao, J.; Ku, T.; Liu, Y. Assessment of milk quality during storage based on DNA extracted from milk. CYTA-J Food. 2018, 16, 786–792. [Google Scholar] [CrossRef] [Green Version]
- Marsili, R. Flavors and off-flavors in dairy foods. In Encyclopedia of Dairy Science; Roginski, H., Fuquay, J.W., Fox, P.F., Eds.; Academic Press: New York, NY, USA, 2003; Volume 1, pp. 1069–1081. [Google Scholar]
- Santos, M.; Ma, Y.; Caplan, Z.; Barbano, D. Sensory threshold of off-flavors caused by proteolysis and lipolysis in milk. J. Dairy Sci. 2003, 86, 1601–1607. [Google Scholar] [CrossRef] [Green Version]
- Mannion, D.T.; Furey, A.; Kilcawley, K.N. Free fatty acids quantification in dairy products. Int. J. Dairy Technol. 2016, 69, 1–12. [Google Scholar] [CrossRef] [Green Version]
- International Dairy Federation (IDF). Significance of Lipolysis in the Manufacture and Storage of Dairy Products; International Dairy Federation: Brussels, Belgium, 1987. [Google Scholar]
- Hanuš, O.; Vegricht, J.; Frelich, J.; Macek, A.; Bjelka, M.; Louda, F.; Janů, L. Analysis of raw cow milk quality according to free fatty acid contents in the Czech Republic. Czech J. Anim. Sci. 2008, 53, 17–30. [Google Scholar] [CrossRef]
- Jacobsen, C. Oxidative Rancidity. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2019; Volume 2, pp. 261–269. [Google Scholar]
- Deeth, H.C. Lipoprotein lipase and lipolysis in milk. Int. Dairy J. 2006, 16, 555–562. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Holland, R.; Crow, V. Esters and their biosynthesis in fermented dairy products: A review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]
- Skaanild, M.T.; Nielsen, T.S. The influence of bovine milk high or low in isoflavones on hepatic gene expression in mice. J. Toxicol. 2010, 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkerk, G. Pasture-based dairying: Challenges and rewards for New Zealand producers. Theriogenology 2003, 59, 553–561. [Google Scholar] [CrossRef]
- Haskell, M.; Rennie, L.; Bowell, V.; Bell, M.; Lawrence, A. Housing system, milk production, and zero-grazing effects on lameness and leg injury in dairy cows. J. Dairy Sci. 2006, 89, 4259–4266. [Google Scholar] [CrossRef] [Green Version]
- Fregonesi, J.; Veira, D.; Von Keyserlingk, M.; Weary, D. Effects of bedding quality on lying behavior of dairy cows. J. Dairy Sci. 2007, 90, 5468–5472. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, T.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.; O’Donovan, M.; Dillon, P.; Ross, R.; Stanton, C. Effect of pasture versus indoor feeding systems on raw milk composition and quality over an entire lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef]
- Villeneuve, M.-P.; Lebeuf, Y.; Gervais, R.; Tremblay, G.; Vuillemard, J.; Fortin, J.; Chouinard, P. Milk volatile organic compounds and fatty acid profile in cows fed timothy as hay, pasture, or silage. J. Dairy Sci. 2013, 96, 7181–7194. [Google Scholar] [CrossRef]
- Kilic, M.; Lindsay, R. Distribution of conjugates of alkylphenols in milk from different ruminant species. J. Dairy Sci. 2005, 88, 7–12. [Google Scholar] [CrossRef]
- Stressler, T.; Leisibach, D.; Lutz-Wahl, S.; Kuhn, A.; Fischer, L. Homologous expression and biochemical characterization of the arylsulfatase from Kluyveromyces lactis and its relevance in milk processing. Appl. Microbiol. Biotechnol. 2016, 100, 5401–5414. [Google Scholar] [CrossRef] [PubMed]
- Steinshamn, H.; Purup, S.; Thuen, E.; Hansen-Møller, J. Effects of clover-grass silages and concentrate supplementation on the content of phytoestrogens in dairy cow milk. J. Dairy Sci. 2008, 91, 2715–2725. [Google Scholar] [CrossRef] [PubMed]
- Daems, F.; Romnee, J.-M.; Heuskin, S.; Froidmont, É.; Lognay, G. Analytical methods used to quantify isoflavones in cow’s milk: A review. Dairy Sci. Technol. 2016, 96, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Mace, T.A.; Ware, M.B.; King, S.A.; Loftus, S.; Farren, M.R.; McMichael, E.; Scoville, S.; Geraghty, C.; Young, G.; Carson, W.E. Soy isoflavones and their metabolites modulate cytokine-induced natural killer cell function. Sci. Rep. 2019, 9, 5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone, K.; Simon Mills, M. Principles and Practice of Phytotherapy: Modern Herbal Medicine, 2nd ed.; Churchill Livingstone Elsevier: Edinburgh, UK; London, UK; New York, NY, USA, 2013. [Google Scholar]
- Smit, S.; Szymańska, E.; Kunz, I.; Roldan, V.G.; van Tilborg, M.W.; Weber, P.; Prudence, K.; van der Kloet, F.M.; van Duynhoven, J.P.; Smilde, A.K. Nutrikinetic modeling reveals order of genistein phase II metabolites appearance in human plasma. Mol. Nutr. Food Res. 2014, 58, 2111–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, H.; Viala, D.; Ollier, A.; Combeau, A.; Besle, J.-M. Isoflavones in several clover species and in milk from goats fed clovers. Biofactors 2004, 22, 237–239. [Google Scholar] [CrossRef]
- Breinholt, V.; Offord, E.; Brouwer, C.; Nielsen, S.; Brøsen, K.; Friedberg, T. In vitro investigation of cytochrome P450-mediated metabolism of dietary flavonoids. Food Chem. Toxicol. 2002, 40, 609–616. [Google Scholar] [CrossRef]
- Jones, K.H.; Trudgill, P.W.; Hopper, D.J. 4-Ethylphenol metabolism by Aspergillus fumigatus. Appl. Environ. Microbiol. 1994, 60, 1978–1983. [Google Scholar]
- Turner, C. Estrogen content of colostrum and milk of dairy cattle. J. Dairy Sci. 1958, 41, 630–640. [Google Scholar] [CrossRef]
- Vialloninsta, C.; Martin, B.; Verdier-Metz, I.; Pradel, P.; Garel, J.-P.; Coulon, J.-B.; Berdagué, J.-L. Transfer of monoterpenes and sesquiterpenes from forages into milk fat. Le lait. 2000, 80, 635–641. [Google Scholar] [CrossRef]
- Romeu-Nadal, M.; Castellote, A.; López-Sabater, M. Headspace gas chromatographic method for determining volatile compounds in infant formulas. J. Chromatogr. A 2004, 1046, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.-M.; de la Hoz, L. Flavour of heated milks. A review. Int. Dairy J. 1992, 2, 69–81. [Google Scholar] [CrossRef]
- Griffiths, M. Improving the Safety and Quality of Milk: Milk Production and Processing; Elsevier: Shaston, UK; Cambridge, UK, 2010. [Google Scholar]
- Bigelis, R.; Weir, P.D.; Jones, R.R.; Umbarger, H. Exogenous valine reduces conversion of leucine to 3-methyl-1-butanol in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1983, 45, 658–664. [Google Scholar] [PubMed]
- Dan, T.; Wang, D.; Wu, S.; Jin, R.; Ren, W.; Sun, T. Profiles of volatile flavor compounds in milk fermented with different proportional combinations of Lactobacillus delbrueckii subsp. Bulgaricus and Streptococcus thermophilus. Molecules 2017, 22, 1633. [Google Scholar]
- Howard, P.H.; Sage, G.; Jarvis, W.; Gray, D. Handbook of Environmental Fate and Exposure Data for Organic Chemicals. Volume II: Solvents; Lewis Publishers: Chelsea, MI, USA, 1990. [Google Scholar]
- Janasik, B.; Jakubowski, M.; Jałowiecki, P. Excretion of unchanged volatile organic compounds (toluene, ethylbenzene, xylene and mesitylene) in urine as result of experimental human volunteer exposure. Int. Arch. Occup. Environ. Health 2008, 81, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Vranova, J.; Ciesarova, Z. Furan in food—A review. Czech. J. Food Sci. 2009, 27, 1–10. [Google Scholar] [CrossRef]
- Wardhan, R. Carcinogenic and organo toxic compounds in milk. Int. J. Health Life Sci. 2016, 2. [Google Scholar] [CrossRef]
- Yue, J.; Zheng, Y.; Liu, Z.; Deng, Y.; Jing, Y.; Luo, Y.; Yu, W.; Zhao, Y. Characterization of volatile compounds in microfiltered pasteurized milk using solid-phase microextraction and GC× GC-TOFMS. Int. J. Food Prop. 2015, 18, 2193–2212. [Google Scholar] [CrossRef]
- Zepka, L.Q.; Garruti, D.S.; Sampaio, K.L.; Mercadante, A.Z.; Da Silva, M.A.A. Aroma compounds derived from the thermal degradation of carotenoids in a cashew apple juice model. Food Res. Int. 2014, 56, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Buchin, S.; Delague, V.; Duboz, G.; Berdague, J.; Beuvier, E.; Pochet, S.; Grappin, R. Influence of pasteurization and fat composition of milk on the volatile compounds and flavor characteristics of a semi-hard cheese. J. Dairy Sci. 1998, 81, 3097–3108. [Google Scholar] [CrossRef]
- Shipe, W.; Ledford, R.; Peterson, R.; Scanlan, R.; Geerken, H.; Dougherty, R.; Morgan, M. Physiological mechanisms involved in transmitting flavors and odors to milk. II. Transmission of some flavor components of silage. J. Dairy Sci. 1962, 45, 477–480. [Google Scholar] [CrossRef]
- Day, E.; Lindsay, R.; Forss, D. Dimethyl sulfide and the flavor of butter. J. Dairy Sci. 1964, 47, 197–199. [Google Scholar] [CrossRef]
- Yoch, D.C. Dimethylsulfoniopropionate: Its sources, role in the marine food web, and biological degradation to dimethylsulfide. Appl. Environ. Microbiol. 2002, 68, 5804–5815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Landaverde, P.A.; Velazquez, G.; Torres, J.; Qian, M. Quantitative determination of thermally derived off-flavor compounds in milk using solid-phase microextraction and gas chromatography. J. Dairy Sci. 2005, 88, 3764–3772. [Google Scholar] [CrossRef]
- Toso, B.; Procida, G.; Stefanon, B. Determination of volatile compounds in cows’ milk using headspace GC-MS. J. Dairy Res. 2002, 69, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, B.R.; Taylor, R.; Madsen, R.; Hyldig, G.; Blenkiron, P.; Jacobsen, C. Investigation of lipid oxidation in the raw materials of a topical skin formulation: A topical skin formulation containing a high lipid content. J. Am. Oil Chem. Soc. 2018, 95, 185–196. [Google Scholar] [CrossRef]
- Centeno, J.A.; Fernández-García, E.; Gaya, P.; Tomillo, J.; Medina, M.; Nunez, M. Volatile compounds in cheeses made from raw ewes’ milk ripened with a lactic culture. J. Dairy Res. 2004, 71, 380–384. [Google Scholar] [CrossRef]
- Fernandez-Garcia, E.; Serrano, C.; Nunez, M. Volatile fraction and sensory characteristics of Manchego cheese. 2. Seasonal variation. J. Dairy Res. 2002, 69, 595–604. [Google Scholar] [CrossRef]
- Rashid, A.; Javed, I.; Rasco, B.; Sablani, S.; Ayaz, M.; Ali, M.A.; Abdullah, M.; Imran, M.; Gondal, T.A.; Afzal, M.I. Measurement of Off-Flavoring Volatile Compounds and Microbial Load as a Probable Marker for Keeping Quality of Pasteurized Milk. Appl. Sci. 2019, 9, 959. [Google Scholar] [CrossRef] [Green Version]
- Havemose, M.S.; Weisbjerg, M.R.; Bredie, W.; Poulsen, H.D.; Nielsen, J.H. Oxidative stability of milk influenced by fatty acids, antioxidants, and copper derived from feed. J. Dairy Sci. 2006, 89, 1970–1980. [Google Scholar] [CrossRef]
- Friedrich, J.E.; Acree, T.E. Gas chromatography olfactometry (GC/O) of dairy products. Int. Dairy J. 1998, 8, 235–241. [Google Scholar] [CrossRef]
- Moio, L.; Langlois, D.; Etievant, P.; Addeo, F. Powerful odorants in bovine, ovine, caprine and water buffalo milk determined by means of gas chromatography–olfactometry. J. Dairy Res. 1993, 60, 215–222. [Google Scholar] [CrossRef]
- Wellnitz-Ruen, W.; Reineccius, G.A.; Thomas, E.L. Analysis of the fruity off-flavor in milk using headspace concentration capillary column gas chromatography. J. Agric. Food Chem. 1982, 30, 512–514. [Google Scholar] [CrossRef]
- Whitfield, F.B.; Jensen, N.; Shaw, K.J. Role of Yersinia intermedia and Pseudomonas putida in the development of a fruity off-flavour in pasteurized milk. J. Dairy Res. 2000, 67, 561–569. [Google Scholar] [CrossRef]
- Al-Attabi, Z.; D’Arcy, B.R.; Deeth, H.C. Volatile sulfur compounds in pasteurised and UHT milk during storage. Dairy Sci. Technol. 2014, 94, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.; Peinado, R. Enological Chemistry; Academic Press: Cambridge, MA, USA, 2012; p. 35. [Google Scholar]
- Brothersen, C.; McMahon, D.; Legako, J.; Martini, S. Comparison of milk oxidation by exposure to LED and fluorescent light. J. Dairy Sci. 2016, 99, 2537–2544. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Sowndhararajan, K.; Jung, J.-W.; Jhoo, J.-W.; Kim, S. Fragrance chemicals in the essential oil of Mentha arvensis reduce levels of mental stress. J. Life Sci. 2013, 23, 933–940. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, A.P.; Guimaratilde, L.; Turatti, I.C.C.; Lopes, N.P.; da Silva Almeida, J.R.G. GC-MS analysis of esterified fatty acids obtained from leaves of wild and cultivated specimens of Leonotis nepetifolia. J. Med. Plant Res. 2015, 9, 525–530. [Google Scholar]
- Wright, M.H.; Lee, C.J.; Arnold, M.S.J.; Shalom, J.; White, A.; Greene, A.C.; Cock, I.E. GC-MS analysis of Tasmannia lanceolata Extracts which Inhibit the Growth of the Pathogenic Bacterium Clostridium perfringens. Pharmacogn. J. 2017, 9, 626–637. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Wang, C.; Huang, A.; Lv, B. Characterization of odors of wood by gas chromatography-olfactometry with removal of extractives as attempt to control indoor air quality. Molecules 2018, 23, 203. [Google Scholar] [CrossRef] [Green Version]
- Menahan, L.; Schultz, L. Metabolism of leucine and valine within the rumen. J. Dairy Sci. 1964, 47, 1080–1085. [Google Scholar] [CrossRef]
- Clayton, G.D.; Clayton, F.E. Patty’s Industrial Hygiene and Toxicology. Vol. 2A. Toxicology; John Wiley & Sons Inc.: Chichester, Sussex, UK, 1981. [Google Scholar]
- Bingham, E.; Cohrssen, B.; Powell, C.H. Patty’s Toxicology. Volume 2: Toxicological Issues Related to Metals, Neurotoxicology and Radiation Metals and Metal Compounds, 5th ed.; Bingham, E., Cohrssen, B., Powell, C.H., Eds.; John Wiley and Sons: Chichester, UK; New York, NY, USA, 2001; Volume 2, p. 712. [Google Scholar]
- Dan, T.; Jin, R.; Ren, W.; Li, T.; Chen, H.; Sun, T. Characteristics of milk fermented by Streptococcus thermophilus MGA45-4 and the profiles of associated volatile compounds during fermentation and storage. Molecules 2018, 23, 878. [Google Scholar]
- Holland, R.; Liu, S.-Q.; Crow, V.; Delabre, M.-L.; Lubbers, M.; Bennett, M.; Norris, G. Esterases of lactic acid bacteria and cheese flavour: Milk fat hydrolysis, alcoholysis and esterification. Int. Dairy J. 2005, 15, 711–718. [Google Scholar] [CrossRef]
- Zhang, X.; Ai, N.; Wang, J.; Tong, L.; Zheng, F.; Sun, B. Lipase-catalyzed modification of the flavor profiles in recombined skim milk products by enriching the volatile components. J. Dairy Sci. 2016, 99, 8665–8679. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, L.; Wang, W. Formation of Aldehyde and Ketone Compounds during Production and Storage of Milk Powder. Molecules 2012, 17, 9900. [Google Scholar] [CrossRef] [Green Version]
- Clarke, H.J.; Mannion, D.T.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Development of a headspace solid-phase microextraction gas chromatography mass spectrometry method for the quantification of volatiles associated with lipid oxidation in whole milk powder using response surface methodology. Food Chem. 2019, 292, 75–80. [Google Scholar] [CrossRef]
- Weimer, B.C. Improving the Flavour of Cheese; Woodhead Publishing: Cambridge, UK, 2007; p. 30. [Google Scholar]
- Shirai, K.; Hisatsuka, K.-I. Isolation and Identification of Styrene Assimilating Bactiria. J. Agricult. Food Chem. 1979, 43, 1595–1596. [Google Scholar]
- Shimada, K.; Kimura, E.; Yasui, Y.; Tanaka, H.; Matsushita, S.; Hagihara, H.; Nagakura, M.; Kawahisa, M. Styrene formation by the decomposition by Pichia carsonii of trans-cinnamic acid added to a ground fish product. Appl. Environ. Microbiol. 1992, 58, 1577–1582. [Google Scholar]
- Im, S.; Hayakawa, F.; Kurata, T. Identification and sensory evaluation of volatile compounds in oxidized porcine liver. J. Agric. Food Chem. 2004, 52, 300–305. [Google Scholar] [CrossRef]
- Repetto, M.; Semprine, J.; Boveris, A. Lipid peroxidation: Chemical mechanism, biological implications and analytical determination. In Lipid Peroxidation; Catala, A., Ed.; Intech Open: London, UK, 2012; Volume 1, pp. 3–24. [Google Scholar]
- Moio, L.; Dekimpe, J.; Etievant, P.; Addeo, F. Neutral volatile compounds in the raw milks from different species. J. Dairy Res. 1993, 60, 199–213. [Google Scholar] [CrossRef]
- Khanal, R.; Dhiman, T.; Ure, A.; Brennand, C.; Boman, R.; McMahon, D.J. Consumer acceptability of conjugated linoleic acid-enriched milk and cheddar cheese from cows grazing on pasture. J. Dairy Sci. 2005, 88, 1837–1847. [Google Scholar] [CrossRef] [Green Version]
- Mathus, T.L.; Townsend, D.E.; Miller, K.W. Anaerobic biogenesis of phenol and p-cresol from L-tyrosine. Fuel 1995, 74, 1505–1508. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M.; Leardi, R.; Toppino, P.M. Influence of heat treatment on the volatile compounds of milk. J. Agric. Food Chem. 1997, 45, 3171–3177. [Google Scholar] [CrossRef]
- Frøst, M.B.; Janhøj, T. Understanding creaminess. Int. Dairy J. 2007, 17, 1298–1311. [Google Scholar] [CrossRef] [Green Version]
- Mela, D.J. Sensory assessment of fat content in fluid dairy products. Appetite 1988, 10, 37–44. [Google Scholar] [CrossRef]
- Martin, B.; Verdier-Metz, I.; Buchin, S.; Hurtaud, C.; Coulon, J.-B. How do the nature of forages and pasture diversity influence the sensory quality of dairy livestock products? Anim. Sci. 2005, 81, 205–212. [Google Scholar] [CrossRef]
- Sapers, G.; Panasiuk, O.; Talley, F.; Shaw, R. Flavor, quality and stability of potato flakes. Effects of Raw Material and Processing. J. Food Sci. 1973, 38, 586–589. [Google Scholar] [CrossRef]
- Murray, K.E.; Shipton, J.; Whitfield, F.B.; Last, J.H. The volatiles of off-flavoured unblanched green peas (Pisum sativum). J. Sci. Food Agric. 1976, 27, 1093–1107. [Google Scholar] [CrossRef]
- Masanetz, C.; Grosch, W. Hay-like off-flavour of dry parsley. Z. Lebensm. Unters. Forsch. A 1998, 206, 114–120. [Google Scholar] [CrossRef]
- Grosch, W.; Konopka, U.; Guth, H. Characterization of off-flavors by aroma extract dilution analysis. In Proceedings of the ACS Symposium Series-American Chemical Society (USA), Washington, DC, USA, 5 August 1992. [Google Scholar]
- Spanier, A.; Shahidi, F.; Parliment, T.; Mussinan, C.; Ho, C.-T.; Contis, E.T.; Servili, M.; Selvaggini, R.; Taticchi, A.; Montedoro, G. Headspace composition of virgin olive oil evaluated by solid phase Microextraction: Relationship with the oil sensory characteristics. In Food Flavors and Chemistry: Advances of the New Millennium; Spanier, A.M., Shahidi, F., Parliment, T.H., Mussinan, C., Ho, C.-T., Tratras Contis, E., Eds.; The Royal Society of Chemistry Publishers: Cambridge, UK, 2001; pp. 236–247. [Google Scholar]
- Suyama, K.; Yeow, T.; Nakai, S. Vitamin A oxidation products responsible for haylike flavor production in nonfat dry milk. J. Agric. Food Chem. 1983, 31, 22–26. [Google Scholar] [CrossRef]
- Sigrist, I.A.; Manzardo, G.G.; Amadò, R. Aroma compounds formed from 3-methyl-2, 4-nonanedione under photooxidative conditions. J. Agric. Food Chem. 2003, 51, 3426–3428. [Google Scholar] [CrossRef] [PubMed]
- Gulati, A.; Galvin, N.; Lewis, E.; Hennessy, D.; O’Donovan, M.; McManus, J.J.; Fenelon, M.A.; Guinee, T.P. Outdoor grazing of dairy cows on pasture versus indoor feeding on total mixed ration: Effects on gross composition and mineral content of milk during lactation. J. Dairy Sci. 2018, 101, 2710–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannion, D.T.; Furey, A.; Kilcawley, K.N. Development and Validation of a Novel Free Fatty Acid Butyl Ester Gas Chromatography Method for the Determination of Free Fatty Acids in Dairy Products. J. Agric. Food Chem. 2019, 67, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Antignac, J.-P.; Cariou, R.; Le Bizec, B.; André, F. New data regarding phytoestrogens content in bovine milk. Food Chem. 2004, 87, 275–281. [Google Scholar] [CrossRef]
- Salum, P.; Erbay, Z.; Kelebek, H.; Selli, S. Optimization of headspace solid-phase microextraction with different fibers for the analysis of volatile compounds of white-brined cheese by using response surface methodology. Food Anal. Methods 2017, 10, 1956–1964. [Google Scholar] [CrossRef]
- Tunick, M.; Iandola, S.; Van Hekken, D. Comparison of SPME Methods for Determining Volatile Compounds in Milk, Cheese, and Whey Powder. Foods 2013, 2, 534. [Google Scholar] [CrossRef]
- Van den Dool, H. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 15 September 2019).
Sample Availability: Samples of the compounds analyzed in this study are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | LRI 1 | Ref LRI 2 | CAS No. | Grass | Grass/Clover | TMR | p-Value |
|---|---|---|---|---|---|---|---|
| Aldehyde | |||||||
| 2-Methyl butanal | 700 | 700 | 96-17-3 | 6.25 × 108 | 2.03 × 108 | 6.29 × 108 | NS 0.067 |
| 3-Methyl butanal | 690 | 692 | 590-86-3 | 5.69 × 108 | 7.97 × 108 | 2.28 × 109 | *** <0.001 |
| Acetaldehyde | 449 | 452 | 75-07-0 | 1.19 × 108 | 2.73 × 108 | 4.38 × 107 | *** 0.002 |
| Butanal | 627 | 622 | 123-72-8 | 8.78 × 107 | 1.81 × 108 | 0.00 × 00 | *** <0.001 |
| Decanal | 1251 | 1256 | 112-31-2 | 4.39 × 109 | 4.90 × 109 | 2.08 × 109 | NS 0.315 |
| Furfural | 870 | 899 | 98-01-1 | 1.09 × 108 | 4.31 × 107 | 2.20 × 107 | *** 0.017 |
| Heptanal | 941 | 943 | 111-71-7 | 1.83 × 109 | 2.75 × 109 | 4.26 × 107 | *** 0.004 |
| Hexanal | 837 | 839 | 66-25-1 | 1.69 × 1010 | 3.59 × 1010 | 1.15 × 109 | *** 0.001 |
| Nonanal | 1147 | 1150 | 124-19-6 | 2.60 × 109 | 5.02 × 109 | 2.43 × 108 | *** 0.001 |
| Octanal | 1044 | 1047 | 124-13-0 | 1.47 × 109 | 2.44 × 109 | 0.00 × 00 | *** 0.001 |
| Pentanal | 735 | 733 | 110-62-3 | 1.49 × 109 | 2.07 × 109 | 0.00 × 00 | *** 0.002 |
| Propanal | 526 | 523 | 123-38-6 | 1.83 × 108 | 4.92 × 108 | 0.00 × 00 | *** <0.001 |
| Methacrolein | 570 | 574 | 78-85-3 | 0.00 × 00 | 6.23 × 106 | 0.00 × 00 | *** 0.006 |
| Ketone | |||||||
| 1-Hydroxy-2-propanone | 734 | 734 | 116-09-6 | 0.00 × 00 | 2.80 × 108 | 4.22 × 107 | *** 0.001 |
| 2-Butanone | 638 | 639 | 78-93-3 | 0.00 × 00 | 0.00 × 00 | 1.08 × 108 | NS 0.080 |
| 2-Heptanone | 932 | 936 | 110-43-0 | 6.22 × 108 | 1.16 × 109 | 2.60 × 108 | *** 0.011 |
| 2-Hexanone | 831 | 834 | 591-78-6 | 1.10 × 108 | 1.94 × 108 | 0.00 × 00 | *** 0.017 |
| 2-Nonanone | 1137 | 1140 | 821-55-6 | 1.18 × 109 | 1.61 × 109 | 0.00 × 00 | *** 0.010 |
| 2-Pentanone | 728 | 730 | 107-87-9 | 5.36 × 108 | 6.40 × 108 | 4.79 × 107 | *** 0.027 |
| 4-Hydroxy-4-methyl-2-pentanone | 913 | 913 | 123-42-2 | 4.00 × 109 | 3.50 × 109 | 1.43 × 108 | *** 0.027 |
| 6-Methyl-5-hepten-2-one | 1031 | 1034 | 110-93-0 | 7.89 × 108 | 2.84 × 109 | 1.07 × 109 | *** <0.001 |
| Acetoin | 778 | 778 | 513-86-0 | 1.33 × 109 | 1.62 × 109 | 2.74 × 108 | NS 0.053 |
| Acetone | 532 | 533 | 67-64-1 | 3.31 × 108 | 2.85 × 109 | 5.43 × 107 | *** <0.001 |
| Acetophenone | 1141 | 1030 | 98-86-2 | 1.08 × 108 | 1.77 × 108 | 9.55 × 107 | NS 0.188 |
| Acetyl valeryl (2,3-heptanedione) | 875 | - | 96-04-8 | 0.00 × 00 | 4.32 × 108 | 0.00 × 00 | *** <0.001 |
| Cyclohexanone | 958 | 957 | 108-94-1 | 1.28 × 109 | 1.33 × 109 | 0.00 × 00 | *** 0.017 |
| Methyl Isobutyl Ketone | 781 | 784 | 108-10-1 | 1.80 × 108 | 0.00 × 00 | 0.00 × 00 | *** <0.001 |
| Ester | |||||||
| 2-Methylbutyl acetate | 906 | 906 | 624-41-9 | 0.00 × 00 | 0.00 × 00 | 1.44 × 108 | NS 0.076 |
| 2-Methylbutyl butanoate | 1080 | - | 51115-64-1 | 0.00 × 00 | 8.43 × 106 | 9.70 × 108 | *** 0.005 |
| Amyl isobutyrate (or isomer) | 1121 | - | 2445-72-9 | 0.00 × 00 | 2.73 × 109 | 2.60 × 1010 | *** <0.001 |
| Amyl propionate | 992 | - | 105-68-0 | 2.59 × 107 | 3.19 × 107 | 6.81 × 108 | *** <0.001 |
| Butyl acetate | 842 | 842 | 123-86-4 | 0.00 × 00 | 0.00 × 00 | 9.12 × 108 | *** <0.001 |
| B-Phenylethyl acetate | 1339 | - | 103-45-7 | 0.00 × 00 | 3.75 × 107 | 2.21 × 108 | *** 0.009 |
| Dimethyl succinate | 1081 | 1082 | 106-65-0 | 2.91 × 107 | 2.87 × 107 | 0.00 × 00 | *** 0.019 |
| Ethyl heptanoate | 1121 | - | 106-30-9 | 6.69 × 107 | 1.13 × 109 | 6.42 × 109 | *** <0.001 |
| Ethyl acetate | 641 | 642 | 141-78-6 | 4.27 × 108 | 1.06 × 109 | 1.10 × 109 | *** 0.002 |
| Ethyl benzoate | 1229 | - | 93-89-0 | 7.81 × 107 | 7.40 × 107 | 1.37 × 107 | NS 0.083 |
| Ethyl butanoate | 823 | 826 | 105-54-4 | 0.00 × 00 | 2.68 × 109 | 3.02 × 1010 | *** <0.001 |
| Ethyl decanoate | 1420 | 1422 | 110-38-3 | 0.00 × 00 | 2.22 × 108 | 1.45 × 108 | *** <0.001 |
| Ethyl dodecanoate | 1622 | 1621 | 106-33-2 | 0.00 × 00 | 1.93 × 108 | 3.05 × 108 | *** <0.001 |
| Ethyl hexanoate | 1021 | 1024 | 123-66-0 | 2.33 × 109 | 8.03 × 109 | 5.44 × 1010 | *** <0.001 |
| Ethyl lactate | 861 | 862 | 97-64-3 | 0.00 × 00 | 1.62 × 108 | 2.60 × 109 | *** <0.001 |
| Ethyl nonanoate | 1319 | - | 123-29-5 | 0.00 × 00 | 6.26 × 108 | 4.45 × 108 | NS 0.104 |
| Ethyl octanoate | 1220 | - | 106-32-1 | 4.51 × 108 | 1.80 × 109 | 4.76 × 109 | *** <0.001 |
| Ethyl pentanoate | 923 | 924 | 539-82-2 | 2.25 × 108 | 1.22 × 109 | 1.34 × 1010 | *** <0.001 |
| Ethyl propanoate | 735 | 737 | 105-37-3 | 2.65 × 108 | 7.16 ×108 | 1.26 × 109 | *** <0.001 |
| Hexyl acetate | 1038 | - | 142-92-7 | 1.83 × 108 | 3.43 × 108 | 2.09 × 109 | *** <0.001 |
| Isoamyl acetate | 902 | 902 | 123-92-2 | 2.48 × 108 | 3.21 × 108 | 5.19 × 108 | *** 0.029 |
| Isoamyl isobutanoate | 1038 | - | 2050-01-3 | 3.07 × 107 | 8.29 × 106 | 2.96 × 109 | *** <0.001 |
| Isobutyl butyrate | 978 | - | 539-90-2 | 0.00 × 00 | 0.00 × 00 | 7.09 × 108 | *** <0.001 |
| Isopentyl hexanoate | 1276 | - | 2198-61-0 | 0.00 × 00 | 2.15 × 107 | 6.22 × 108 | *** <0.001 |
| Methyl butanoate | 748 | - | 623-42-7 | 9.77 × 107 | 9.73 × 108 | 3.04 × 109 | *** <0.001 |
| Methyl decanoate | 1351 | - | 110-42-9 | 0.00 × 00 | 2.91 × 108 | 0.00 × 00 | *** <0.001 |
| Methyl dodecanoate | 1550 | - | 111-82-0 | 0.00 × 00 | 1.19 × 108 | 6.07 × 107 | *** <0.001 |
| Methyl hexanoate | 949 | - | 106-70-7 | 5.52 × 108 | 7.42 × 109 | 1.13 × 1010 | *** <0.001 |
| Methyl propionate | 657 | - | 554-12-1 | 1.95 × 107 | 1.63 × 108 | 1.08 × 108 | *** <0.001 |
| n-Propyl acetate | 739 | - | 109-60-4 | 1.60 × 109 | 1.97 × 109 | 3.43 × 109 | *** 0.008 |
| Pentyl acetate | 901 | - | 628-63-7 | 6.38 × 108 | 1.72 × 108 | 7.74 × 108 | *** 0.012 |
| Propyl 2-methylbutanoate | 969 | - | 37064-20-3 | 3.67 × 107 | 5.63 × 107 | 0.00 × 00 | NS 0.112 |
| Propyl butyrate | 921 | - | 644-49-5 | 2.57 × 109 | 3.56 × 109 | 3.28 × 1010 | *** <0.001 |
| Propyl hexanoate | 1118 | - | 626-77-7 | 1.53 × 109 | 3.07 × 109 | 3.57 × 1010 | *** <0.001 |
| Alcohol | *** <0.001 | ||||||
| 1-Hexanol | 903 | 916 | 111-27-3 | 7.07 × 108 | 1.95 × 109 | 8.42 × 108 | NS 0.502 |
| 1-Octanol | 1112 | 1118 | 111-87-5 | 1.56 × 109 | 2.39 × 109 | 0.00 × 00 | *** 0.008 |
| 1-Pentanol | 816 | 815 | 71-41-0 | 1.02 × 109 | 1.30 × 109 | 0.00 × 00 | *** 0.018 |
| 1-Propanol | 612 | 612 | 71-23-8 | 2.10 × 109 | 3.68 × 109 | 1.06 × 109 | *** 0.007 |
| 1-Methoxy-2-propanol | 713 | 713 | 107-98-2 | 4.91 × 108 | 4.80 × 108 | 1.49 × 107 | *** 0.043 |
| 2-Methyl-1-butanol | 783 | 789 | 137-32-6 | 2.75 × 108 | 3.89 × 108 | 2.77 × 108 | NS 0.418 |
| 2-Methyl-1-propanol | 678 | 678 | 78-83-1 | 4.48 × 107 | 8.06 × 107 | 7.45 × 106 | *** 0.005 |
| 2-Methyl propanol | 609 | - | 78-84-2 | 6.69 × 107 | 7.84 × 107 | 0.00 × 00 | NS 0.112 |
| 2-Butanol | 648 | 648 | 78-92-2 | 4.15 × 108 | 6.63 × 108 | 1.28 × 108 | *** 0.005 |
| 3-Methyl-1-butanol | 783 | 784 | 123-51-3 | 2.41 × 108 | 1.54 × 108 | 2.45 × 107 | *** 0.043 |
| Ethanol | 506 | 506 | 64-17-5 | 1.23 × 109 | 4.60 × 109 | 1.33 × 109 | *** <0.001 |
| Isopropyl Alcohol | 543 | - | 67-63-0 | 6.64 × 107 | 1.23 × 108 | 1.80 × 107 | *** 0.011 |
| Phenylethyl Alcohol | 1199 | - | 60-12-8 | 0.00 × 00 | 9.02 × 107 | 8.73 × 108 | *** <0.001 |
| Acid | |||||||
| 2,2-Dimethyl-propanoic acid | 837 | 869 | 75-98-9 | 0.00 × 00 | 2.13 × 108 | 0.00 × 00 | *** 0.004 |
| 3-Methyl-butanoic acid | 918 | - | 503-74-2 | 0.00 × 00 | 0.00 × 00 | 1.04 × 1010 | *** <0.001 |
| Acetic acid | 690 | 690 | 64-19-7 | 8.94 × 109 | 1.02 × 1010 | 9.85 × 1010 | *** <0.001 |
| Butanoic acid | 864 | 864 | 107-92-6 | 8.80 × 109 | 9.17 × 109 | 1.04 × 1011 | *** <0.001 |
| Pentanoic acid | 995 | - | 109-52-4 | 0.00 × 00 | 0.00 × 00 | 2.51 × 1010 | *** <0.001 |
| Propanoic acid | 778 | 802 | 79-09-4 | 1.51 × 109 | 1.09 × 109 | 2.02 × 1010 | *** <0.001 |
| Hexanoic acid | 1052 | 1052 | 142-62-1 | 3.23 × 109 | 2.90 × 109 | 2.49 × 1010 | *** <0.001 |
| Fatty acid esters | *** <0.001 | ||||||
| Propanoic acid, butyl ester | 931 | - | 590-01-2 | 1.79 × 108 | 0.00 × 00 | 9.44 × 108 | *** <0.001 |
| Butanoic acid, butyl ester | 978 | - | 109-21-7 | 0.00 × 00 | 0.00 × 00 | 6.04 × 109 | *** <0.001 |
| Terpene | |||||||
| 3-Carene | 1035 | 1027 | 13466-78-9 | 5.45 × 107 | 2.41 × 107 | 0.00 × 00 | NS 0.084 |
| α-Pinene | 954 | 951 | 80-56-8 | 1.34 × 107 | 0.00 × 00 | 8.17 × 106 | NS 0.270 |
| Cumene | 991 | - | 98-82-8 | 6.13 × 107 | 1.11 × 108 | 5.04 × 107 | NS 0.166 |
| Mesitylene | 1029 | - | 108-67-8 | 8.64 × 108 | 2.03 × 109 | 1.11 × 107 | *** 0.044 |
| Furan | |||||||
| 2-Ethyl furan | 717 | 720 | 3208-16-0 | 4.50 × 108 | 1.25 × 109 | 1.89 × 107 | *** <0.001 |
| 2-Methyl furan | 615 | 615 | 534-22-5 | 1.64 × 107 | 4.55 × 107 | 0.00 × 00 | *** 0.005 |
| 2-Pentyl furan | 1010 | 1012 | 3777-69-3 | 1.12 × 109 | 2.52 × 109 | 5.17 × 108 | *** 0.004 |
| 2-n-Butyl furan | 917 | - | 4466-24-4 | 0.00 × 00 | 6.00 × 107 | 0.00 × 00 | NS 0.079 |
| 2,4-Dimethyl furan | 733 | - | 3710-43-8 | 3.27 × 107 | 3.11 × 107 | 0.00 × 00 | *** 0.019 |
| Hydrocarbon | |||||||
| 1,3-bis(1,1-dimethylethyl)-benzene | 1284 | - | 1014-60-4 | 1.31 × 1011 | 1.73 × 1011 | 5.81 × 1010 | NS 0.060 |
| 2,4-Dimethyl-benzaldehyde | 1305 | - | 15764-16-6 | 7.46 × 108 | 9.18 × 108 | 2.93 × 108 | *** 0.019 |
| Benzaldehyde | 1027 | 1032 | 100-52-7 | 3.41 × 109 | 3.44 × 109 | 2.05 × 109 | NS 0.466 |
| Benzothiazole | 1320 | - | 95-16-9 | 4.06 × 108 | 6.04 × 108 | 5.28 × 108 | NS 0.156 |
| Ethylbenzene | 897 | 890 | 100-41-4 | 0.00 × 00 | 0.00 × 00 | 6.02 × 107 | *** <0.001 |
| Mesitylene | 1028 | - | 108-67-8 | 8.64 × 108 | 2.03 × 109 | 1.11 × 107 | *** 0.044 |
| o-Cymene | 1056 | - | 527-84-4 | 1.72 × 107 | 4.73 × 106 | 0.00 × 00 | NS 0.055 |
| o-xylene | 925 | 916 | 95-47-6 | 1.09 × 108 | 9.63 × 108 | 0.00 × 00 | NS 0.108 |
| p-Xylene | 895 | 895 | 106-42-3 | 7.52 × 108 | 1.32 × 109 | 5.47 × 108 | NS 0.146 |
| p-Cresol | 1193 | - | 106-44-5 | 4.52 × 108 | 2.89 × 108 | 4.40 × 108 | NS 0.113 |
| Styrene | 927 | 929 | 100-42-5 | 1.21 × 108 | 2.37 × 108 | 1.07 × 108 | NS 0.078 |
| tert-Butylbenzene | 1024 | - | 98-06-6 | 4.74 × 108 | 6.60 × 108 | 1.53 × 108 | *** 0.037 |
| Toluene | 792 | 794 | 108-88-3 | 8.30 × 107 | 1.31 × 108 | 3.62 × 107 | *** 0.040 |
| Phenolic | |||||||
| Phenol | 1096 | 1112 | 108-95-2 | 0.00 × 00 | 0.00 × 00 | 5.87 × 108 | *** <0.001 |
| 2-Methoxy-4-vinylphenol | 1150 | - | 7786-61-0 | 0.00 × 00 | 0.00 × 00 | 3.98 × 107 | *** <0.001 |
| Sulfur | |||||||
| Dimethyl sulfide | 537 | 538 | 75-18-3 | 1.78 × 108 | 2.90 × 108 | 6.17 × 107 | *** 0.004 |
| Dimethyl sulfone | 1054 | 1055 | 67-71-0 | 2.21 × 108 | 6.32 × 107 | 0.00 × 00 | *** 0.014 |
| Methanethiol | 459 | 462 | 74-93-1 | 9.29 × 106 | 1.02 × 107 | 8.84 × 106 | NS 0.855 |
| Ether | |||||||
| Vinylisopentyl ether | 765 | - | 39782-38-2 | 1.10 × 108 | 2.89 × 108 | 3.71 × 107 | *** 0.033 |
| Lactone | |||||||
| y-Hexalactone | 1166 | - | 695-06-7 | 8.51 × 108 | 1.07 × 109 | 4.06 × 108 | *** 0.040 |
| y-Nonalactone | 1489 | - | 104-61-0 | 4.69 × 108 | 6.44 × 108 | 3.75 × 108 | NS 0.279 |
| Pyrazine | |||||||
| 2,3,5-Trimethyl-6-ethylpyrazine | 1190 | - | 17398-16-2 | 0.00 × 00 | 0.00 × 00 | 1.48 × 108 | *** <0.001 |
| 2,3-Dimethyl-pyrazine | 961 | - | 5910-89-4 | 1.27 × 108 | 3.77 × 107 | 1.33 × 109 | *** <0.001 |
| 3-Ethyl-2,5-dimethyl-pyrazine | 1055 | - | 5910-89-4 | 0.00 × 00 | 0.00 × 00 | 4.71 × 107 | *** <0.001 |
| Pyrazine | 771 | - | 290-37-9 | 5.46 × 107 | 5.55 × 107 | 4.79 × 107 | NS 0.847 |
| Trimethyl-pyrazine | 1041 | 1041 | 14667-55-1 | 8.50 × 107 | 3.89 × 107 | 9.03 × 108 | *** <0.001 |
| Compound | CAS No. | LRI 1 | Grass d 3 | Grass/Clover d 3 | TMR d 3 | Grass d 9 | Grass/Clover d 9 | TMR Day 9 | Grass d 14 | Grass/Clover d 14 | TMR Day 14 | p-Value | p-Value (Grass) | p-Value (Grass/Clover) | p-Value (TMR) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aldehyde | |||||||||||||||
| (E)-2-Octenal (or isomer) | 2548-87-0 | 1094 | 0.00 × 00 | 0.00 × 00 | 1.74 × 107 | 7.29 × 107 | 2.51 × 108 | 2.22 × 107 | 2.85 × 108 | 1.11 × 109 | 2.18 × 108 | * <0.001 | NS 0.051 | * 0.003 | * 0.05 |
| (Z)-2-Heptenal (or isomer) | 57266-86-1 | 1012 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.03 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | ND | ND | * 0.006 |
| Acetaldehyde | 75-07-0 | 449 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.85 × 107 | 0.00 × 00 | 2.40 × 106 | * 0.029 | NS 0.302 | ND | NS 0.129 |
| 3-Methyl-butanal | 590-86-3 | 690 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 9.93 × 107 | 7.12 × 108 | 1.75 × 107 | * <0.001 | * <0.001 | * <0.001 | * 0.002 |
| Decanal | 112-31-2 | 1250 | 1.47 × 107 | 1.29 × 107 | 3.26 × 106 | 8.28 × 106 | 5.23 × 106 | 4.97 × 106 | 1.18 × 107 | 3.73 × 106 | 3.52 × 106 | NS 0.477 | NS 0.658 | NS 0.515 | NS 0.736 |
| Heptanal | 111-71-7 | 941 | 1.09 × 108 | 1.08 × 108 | 1.49 × 108 | 1.06 × 107 | 1.72 × 106 | 7.27 × 108 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * 0.001 | * <0.001 | * <0.001 |
| Hexanal | 66-25-1 | 838 | 3.69 × 108 | 4.02 × 108 | 1.72 × 109 | 0.00 × 00 | 0.00 × 00 | 3.56 × 109 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * 0.000 | * <0.001 | * <0.001 |
| Nonanal | 124-19-6 | 1147 | 5.31 × 107 | 5.02 × 107 | 6.91 × 107 | 3.66 × 107 | 2.81 × 107 | 1.23 × 108 | 3.98 × 107 | 1.90 × 107 | 3.44 × 107 | * <0.001 | NS 0.259 | * <0.001 | * <0.001 |
| Octanal | 124-13-0 | 1044 | 2.22 × 107 | 3.11 × 107 | 4.02 × 107 | 1.21 × 107 | 0.00 × 00 | 8.73 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * 0.007 | * 0.009 | * <0.001 |
| Pentanal | 110-62-3 | 733 | 1.39 × 108 | 1.96 × 108 | 9.84 × 106 | 0.00 × 00 | 0.00 × 00 | 1.82 × 108 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | * <0.001 | * <0.001 |
| Ketone | |||||||||||||||
| 2-Butanone | 78-93-3 | 637 | 3.86 × 107 | 1.05 × 108 | 1.49 × 108 | 5.86 × 107 | 1.11 × 108 | 1.48 × 108 | 2.64 × 107 | 9.64 × 107 | 7.42 × 107 | * <0.001 | NS 0.127 | * 0.007 | * <0.001 |
| 2-Heptanone | 110-43-0 | 933 | 3.37 × 107 | 3.60 × 107 | 3.20 × 107 | 4.37 × 108 | 8.14 × 108 | 4.67 × 107 | 1.52 × 109 | 3.63 × 109 | 9.04 × 109 | * <0.001 | * <0.001 | * <0.001 | * <0.001 |
| 2-Hexanone | 591-78-6 | 831 | 1.77 × 107 | 9.14 × 106 | 8.93 × 106 | 2.33 × 107 | 2.96 × 107 | 2.35 × 106 | 2.63 × 107 | 6.70 × 107 | 8.27 × 107 | * <0.001 | NS 0.603 | * <0.001 | * <0.001 |
| 2-Nonanone | 821-55-6 | 1137 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.25 × 108 | 1.49 × 108 | 0.00 × 00 | 5.16 × 108 | 5.69 × 108 | 2.46 × 109 | * <0.001 | * 0.005 | * 0.002 | * <0.001 |
| 2-Octanone | 111-13-7 | 1034 | 6.82 × 106 | 1.01 × 107 | 1.13 × 107 | 1.04 × 107 | 2.62 × 107 | 2.21 × 106 | 2.31 × 107 | 5.00 × 107 | 5.10 × 107 | * <0.001 | * 0.027 | * <0.001 | * 0.001 |
| 2-Pentanone | 107-87-9 | 727 | 5.70 × 107 | 5.47 × 107 | 5.36 × 107 | 1.06 × 108 | 2.17 × 108 | 4.00 × 107 | 1.94 × 108 | 5.48 × 108 | 6.91 × 108 | * <0.001 | * <0.001 | * <0.001 | * <0.001 |
| 2-Undecanone | 112-12-9 | 1353 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 5.97 × 106 | 4.63 × 105 | 0.00 × 00 | 3.82 × 107 | 1.52 × 107 | 2.78 × 108 | * <0.001 | NS 0.262 | * 0.047 | * <0.001 |
| 2,3-Pentanedione | 600-14-6 | 736 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.03 × 108 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | ND | ND | * 0.007 |
| 3,5-(E,E)-Octadien-2-one (or isomer) | 30086-02-3 | 1130 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.17 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | ND | ND | * 0.015 |
| 3-Hexen-2-one | 763-93-9 | 839 | 6.97 × 106 | 7.68 × 106 | 2.76 × 106 | 1.59 × 107 | 1.54 × 107 | 0.00 × 00 | 1.53 × 107 | 8.90 × 106 | 1.58 × 107 | NS 0.177 | NS 0.557 | NS 0.517 | NS 0.051 |
| 4-Methyl-3-pentene-2-one (tentative) | 141-79-7 | 839 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.15 × 107 | 5.55 × 106 | 0.00 × 00 | 9.92 × 106 | 1.23 × 107 | 1.58 × 107 | * 0.047 | NS 0.500 | ND 0.065 | * 0.025 |
| 5-Hepten-2-one (tentative) | 6714-00-7 | 921 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.42 × 107 | 5.23 × 107 | * <0.001 | ND | * <0.001 | * <0.001 |
| Acetone | 67-64-1 | 532 | 9.16 × 108 | 8.04 × 108 | 1.14 × 109 | 7.81 × 108 | 7.97 × 108 | 1.22 × 109 | 3.00 × 108 | 7.20 × 108 | 6.65 × 108 | * <0.001 | * 0.003 | NS 0.538 | * <0.001 |
| Acetophenone | 98-86-2 | 1030 | 6.38 × 106 | 2.05 × 106 | 1.64 × 106 | 0.00 × 00 | 4.23 × 106 | 3.22 × 106 | 1.77 × 106 | 1.06 × 106 | 0.00 × 00 | * 0.044 | NS 0.114 | NS 0.113 | NS 0.251 |
| Cyclohexanone | 110-82-7 | 956 | 7.99 × 106 | 0.00 × 00 | 0.00 × 00 | 1.37 × 107 | 1.67 × 106 | 0.00 × 00 | 6.52 × 106 | 1.33 × 106 | 1.33 × 106 | * 0.046 | NS 0.728 | NS 0.623 | NS 0.422 |
| Acetyl valeryl (2,3-heptanedione) | 96-04-8 | 875 | 3.78 × 105 | 2.52 × 105 | 4.29 × 105 | 3.31 × 106 | 3.56 × 107 | 1.77 × 105 | 0.00 × 00 | 2.39 × 107 | 1.31 × 106 | * <0.001 | NS 0.194 | * 0.001 | NS 0.582 |
| Methyl Isobutyl Ketone | 108-10-1 | 780 | 2.05 × 108 | 1.47 × 108 | 2.00 × 108 | 2.14 × 108 | 1.73 × 108 | 2.46 × 108 | 1.74 × 108 | 1.78 × 108 | 1.75 × 108 | * <0.001 | * 0.035 | * 0.023 | * 0.006 |
| Ester | |||||||||||||||
| Ethyl heptanoate | 106-30-9 | 1120 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.08 × 107 | 0.00 × 00 | 1.35 × 108 | * <0.001 | * <0.001 | ND | * <0.001 |
| Ethyl (Z)-2-butenoate | 6776-19-8 | 875 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.12 × 108 | 2.51 × 106 | 1.54 × 108 | * <0.001 | * <0.001 | NS 0.422 | * <0.001 |
| Ethyl 2-methylbutanoate | 7452-79-1 | 872 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.38 × 107 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | ND | ND |
| Ethyl 3-methylbutanoate | 108-64-5 | 876 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.81 × 108 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | ND | ND |
| Ethyl acetate | 141-78-6 | 639 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.02 × 107 | 8.47 × 106 | 0.00 × 00 | 2.44 × 108 | 6.08 × 107 | 1.85 × 108 | * <0.001 | * <0.001 | * <0.001 | * <0.001 |
| Ethyl butanoate | 105-54-4 | 823 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 6.10 × 109 | 0.00 × 00 | 1.02 × 1010 | * <0.001 | * <0.001 | ND | * <0.001 |
| Ethyl decanoate | 110-38-3 | 1419 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.30 × 108 | 9.37 × 106 | 1.59 × 109 | * <0.001 | * 0.019 | NS 0.155 | * <0.001 |
| Ethyl hexanoate | 123-66-0 | 1021 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.18 × 108 | 0.00 × 00 | 0.00 × 00 | 6.90 × 109 | 2.80 × 107 | 8.91 × 109 | * <0.001 | * <0.001 | * 0.005 | * <0.001 |
| Ethyl octanoate | 106-32-1 | 1220 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 7.85 × 108 | 3.72 × 106 | 3.07 × 109 | * <0.001 | * <0.001 | NS 0.105 | * <0.001 |
| Ethyl pentanoate | 539-82-2 | 923 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 5.50 × 107 | 0.00 × 00 | 8.08 × 107 | * <0.001 | * 0.001 | ND | * <0.001 |
| Ethyl propanoate | 105-37-3 | 735 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.73 × 106 | 0.00 × 00 | 2.51 × 107 | * <0.001 | NS 0.465 | ND | * <0.001 |
| Methyl butanoate | 623-42-7 | 747 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.25 × 106 | 4.90 × 106 | 0.00 × 00 | 9.84 × 105 | 2.78 × 107 | 1.47 × 107 | * <0.001 | NS 0.590 | * <0.001 | * <0.001 |
| Methyl decanoate | 110-42-9 | 1350 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.52 × 106 | NS 0.084 | ND | ND | NS 0.143 |
| Methyl hexanoate | 106-70-7 | 949 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.34 × 106 | 0.00 × 00 | 0.00 × 00 | 3.65 × 106 | 0.00 × 00 | 3.00 × 107 | * <0.001 | NS 0.244 | ND | * <0.001 |
| Methyl methacrylate | 80-62-6 | 736 | 7.47 × 106 | 2.22 × 106 | 3.88 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * 0.002 | * <0.001 | NS 0.422 | * <0.001 |
| Alcohol | |||||||||||||||
| 1-Butanol | 71-36-3 | 715 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.08 × 106 | 7.33 × 105 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * 0.040 | ND | NS 0.080 | NS 0.422 |
| 2-Methyl-1-butanol | 137-32-6 | 765 | 2.27 × 107 | 2.15 × 107 | 1.97 × 107 | 2.23 × 107 | 2.02 × 107 | 1.95 × 107 | 0.00 × 00 | 0.00 × 00 | 1.18 × 108 | * <0.001 | * 0.003 | * 0.003 | * <0.001 |
| 3-Methyl-1-butanol | 123-51-3 | 767 | 5.73 × 107 | 6.92 × 107 | 1.11 × 108 | 5.48 × 107 | 3.01 × 107 | 5.04 × 107 | 1.05 × 109 | 2.56 × 109 | 3.33 × 108 | * <0.001 | * 0.011 | * <0.001 | * <0.001 |
| 1-Hexanol | 111-27-3 | 894 | 0.00 × 00 | 0.00 × 00 | 5.83 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | ND | NS 0.080 | * 0.004 |
| 2-Ethyl-1-hexanol | 104-76-7 | 1075 | 3.69 × 107 | 3.34 × 107 | 1.96 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | * 0.001 | NS 0.108 |
| 1-Octanol | 111-87-5 | 1116 | 0.00 × 00 | 0.00 × 00 | 2.06 × 107 | 0.00 × 00 | 0.00 × 00 | 2.22 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | ND | ND | * 0.012 |
| 1-Pentanol | 71-41-0 | 794 | 6.10 × 107 | 9.15 × 107 | 1.57 × 107 | 2.95 × 107 | 5.39 × 107 | 8.62 × 107 | 0.00 × 00 | 2.52 × 106 | 2.52 × 107 | * <0.001 | NS 0.075 | * <0.001 | * <0.001 |
| Ethanol | 64-17-5 | 505 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.35 × 107 | 0.00 × 00 | 2.10 × 108 | 4.36 × 108 | 1.14 × 109 | * 0.000 | * 0.005 | NS 0.070 | * 0.003 |
| Isopropyl Alcohol | 67-63-0 | 541 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.54 × 107 | 2.33 × 107 | 0.00 × 00 | 4.29 × 107 | 2.82 × 107 | 6.11 × 107 | * <0.001 | NS 0.194 | NS 0.146 | * <0.001 |
| Acid | |||||||||||||||
| Butanoic acid | 107-92-6 | 863 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 9.61 × 108 | NS 0.526 | ND | ND | NS 0.385 |
| Hexanoic acid | 142-62-1 | 1052 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 2.70 × 108 | 0.00 × 00 | 6.72 × 109 | NS 0.371 | NS 0.259 | ND | NS 0.306 |
| Octanoic acid | 124-07-2 | 1245 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 1.67 × 109 | NS 0.471 | ND | ND | NS 0.358 |
| Propanoic acid, 2-methyl-, 3-hydroxy-2,2,4-trimethylpentyl ester | 77-68-9 | 1460 | 6.78 × 107 | 5.62 × 107 | 3.46 × 107 | 1.62 × 107 | 2.71 × 107 | 1.01 × 107 | 3.01 × 107 | 5.71 × 106 | 7.09 × 105 | * 0.003 | NS 0.026 | * 0.015 | NS 0.181 |
| Terpene | |||||||||||||||
| 3-Carene | 13466-78-9 | 1035 | 0.00 × 00 | 0.00 × 00 | 1.34 × 107 | 0.00 × 00 | 9.50 × 105 | 7.02 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | NS 0.144 | ND | NS 0.144 | NS 0.285 |
| α-Pinene | 80-56-8 | 953 | 7.81 × 106 | 6.03 × 106 | 4.56 × 106 | 7.50 × 107 | 6.39 × 107 | 4.88 × 107 | 1.62 × 107 | 2.73 × 107 | 1.10 × 107 | * <0.001 | * 0.001 | * 0.012 | * 0.003 |
| Cumene | 98-82-8 | 990 | 2.26 × 106 | 2.80 × 106 | 3.47 × 106 | 5.58 × 106 | 7.86 × 106 | 3.13 × 106 | 3.71 × 106 | 1.06 × 107 | 1.64 × 107 | * 0.035 | NS 0.323 | NS 0.220 | * 0.037 |
| D-Limonene | 5989-27-5 | 1055 | 0.00 × 00 | 2.78 × 107 | 1.61 × 107 | 1.26 × 106 | 3.05 × 105 | 1.77 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | NS 0.465 | * <0.001 | 0.013 |
| Mesitylene | 108-67-8 | 1028 | 4.47 × 107 | 3.71 × 107 | 5.38 × 107 | 5.14 × 107 | 4.54 × 107 | 2.98 × 107 | 7.23 × 107 | 6.79 × 107 | 8.22 × 107 | * 0.017 | NS 0.104 | NS 0.070 | * 0.042 |
| trans-β-Ocimene (or isomer) | 3779-61-1 | 1035 | 0.00 × 00 | 1.74 × 107 | 1.34 × 107 | 0.00 × 00 | 0.00 × 00 | 1.77 × 107 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * 0.020 | ND | NS 0.088 | NS 0.067 |
| Furan | |||||||||||||||
| 2,4-Dimethylfuran | 3710-43-8 | 732 | 8.90 × 106 | 3.64 × 106 | 7.27 × 106 | 1.11 × 107 | 1.36 × 107 | 0.00 × 00 | 8.34 × 106 | 1.56 × 107 | 1.17 × 107 | * <0.001 | NS 0.357 | * <0.001 | * <0.001 |
| 2,5-Dimethylfuran | 625-86-5 | 734 | 8.90 × 106 | 3.64 × 106 | 7.27 × 106 | 1.11 × 107 | 1.36 × 107 | 0.00 × 00 | 8.34 × 106 | 1.56 × 107 | 1.17 × 107 | * <0.001 | NS 0.357 | * 0.002 | * <0.001 |
| 2-Ethylfuran | 3208-16-0 | 717 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 3.04 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | NS 0.090 | ND | NS 0.422 | NS 0.172 |
| Hydrocarbon | |||||||||||||||
| 2,6-Bis(1,1-dimethylethyl)-4-(1-oxopropyl)phenol (tentative) | 14035-34-8 | 1684 | 1.13 × 107 | 1.88 × 107 | 1.35 × 107 | 2.41 × 107 | 3.89 × 107 | 2.39 × 107 | 5.18 × 106 | 1.63 × 107 | 2.83 × 107 | * 0.012 | NS 0.347 | * 0.008 | NS 0.171 |
| 2,4-Dimethyl-benzaldehyde | 15764-16-6 | 1305 | 6.81 × 106 | 2.24 × 106 | 2.48 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | * 0.028 | * 0.028 |
| Benzene | 71-43-2 | 684 | 2.55 × 106 | 2.57 × 106 | 4.69 × 106 | 3.63 × 106 | 8.95 × 107 | 1.98 × 105 | 3.21 × 106 | 4.13 × 107 | 2.80 × 106 | * <0.001 | NS 0.943 | * 0.017 | NS 0.232 |
| 1,2,3-Trimethyl-benzene | 526-73-8 | 1028 | 4.47 × 107 | 3.71 × 107 | 5.38 × 107 | 5.14 × 107 | 4.54 × 107 | 2.98 × 107 | 7.23 × 107 | 6.79 × 107 | 8.22 × 107 | * 0.017 | NS 0.104 | NS 0.070 | * 0.042 |
| 1,3-Bis(1,1-dimethylethyl)-benzene | 1014-60-4 | 1284 | 3.49 × 108 | 3.08 × 108 | 2.33 × 108 | 3.67 × 108 | 3.10 × 108 | 2.01 × 108 | 4.52 × 108 | 4.41 × 108 | 6.52 × 108 | * <0.001 | NS 0.185 | * 0.007 | * <0.001 |
| Ethylbenzene | 100-41-4 | 897 | 1.01 × 108 | 8.28 × 107 | 9.66 × 107 | 1.69 × 108 | 2.08 × 108 | 7.05 × 107 | 2.07 × 108 | 3.05 × 108 | 3.32 × 108 | * <0.001 | * 0.004 | * <0.001 | * <0.001 |
| o-Cymene | 527-84-4 | 1055 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 5.76 × 106 | 3.30 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * <0.001 | * <0.001 | NS 0.128 | ND |
| p-Cresol | 106-44-5 | 1182 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 8.49 × 108 | 3.62 × 108 | 2.48 × 108 | 6.35 × 107 | 5.22 × 107 | 5.46 × 107 | * <0.001 | * 0.003 | * <0.001 | * <0.001 |
| p-Xylene | 106-42-3 | 895 | 1.01 × 108 | 8.28 × 107 | 9.66 × 107 | 1.69 × 108 | 2.08 × 108 | 7.05 × 107 | 2.07 × 108 | 3.05 × 108 | 3.32 × 108 | * <0.001 | * 0.004 | * <0.001 | * <0.001 |
| Styrene | 100-42-5 | 927 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 6.15 × 106 | 0.00 × 00 | 2.92 × 106 | 4.11 × 106 | 0.00 × 00 | 8.62 × 106 | NS 0.223 | NS 0.631 | ND | NS 0.214 |
| tert-Butylbenzene | 98-06-6 | 1024 | 9.63 × 106 | 8.24 × 106 | 6.27 × 106 | 8.48 × 106 | 6.51 × 106 | 3.21 × 106 | 8.80 × 106 | 1.23 × 107 | 1.32 × 107 | NS 0.072 | NS 0.815 | NS 0.223 | * 0.044 |
| Toluene | 108-88-3 | 792 | 1.94 × 109 | 1.27 × 109 | 4.98 × 107 | 1.97 × 109 | 1.41 × 109 | 4.51 × 107 | 1.59 × 109 | 1.22 × 109 | 4.97 × 107 | * <0.001 | NS 0.075 | * 0.008 | NS 0.695 |
| Phenolic | |||||||||||||||
| Phenol | 108-95-2 | 1093 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.65 × 106 | 0.00 × 00 | 5.29 × 106 | 9.74 × 106 | 0.00 × 00 | 5.77 × 106 | * 0.021 | NS 0.205 | ND | NS 0.220 |
| 2,4-Di-tert-butylphenol | 96-76-4 | 1595 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 5.79 × 107 | 1.39 × 107 | 1.38 × 107 | 2.37 × 107 | 2.00 × 106 | 1.01 × 107 | * 0.043 | NS 0.363 | NS 0.065 | NS 0.130 |
| Sulfur | |||||||||||||||
| Dimethyl sulfide | 75-18-3 | 536 | 1.11 × 107 | 1.05 × 107 | 6.30 × 106 | 2.39 × 107 | 1.75 × 107 | 0.00 × 00 | 1.66 × 108 | 1.38 × 108 | 5.51 × 107 | * <0.001 | * <0.001 | * <0.001 | * <0.001 |
| Dimethyl sulfone | 67-71-0 | 1052 | 3.96 × 107 | 2.94 × 107 | 0.00 × 00 | 3.88 × 107 | 2.80 × 107 | 0.00 × 00 | 3.60 × 107 | 2.41 × 107 | 0.00 × 00 | * <0.001 | NS 0.961 | NS 0.675 | ND |
| Dimethyl disulfide | 624-92-0 | 776 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * 0.033 | ND | NS 0.093 | NS 0.422 |
| Methanethiol | 74-93-1 | 459 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 4.79 × 107 | 0.00 × 00 | * <0.001 | ND | * 0.013 | ND |
| Ether | |||||||||||||||
| Ethyl ether | 60-29-7 | 514 | 1.86 × 107 | 1.09 × 107 | 6.58 × 106 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | 0.00 × 00 | * 0.006 | NS 0.013 | NS 0.086 | NS 0.232 |
| Vinylisopentyl ether | 39782-38-2 | 767 | 5.73 × 107 | 7.61 × 107 | 7.42 × 107 | 4.76 × 107 | 3.01 × 107 | 2.71 × 107 | 0.00 × 00 | 0.00 × 00 | 1.35 × 108 | NS 0.377 | NS 0.645 | NS 0.234 | NS 0.337 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clarke, H.J.; Griffin, C.; Rai, D.K.; O’Callaghan, T.F.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Dietary Compounds Influencing the Sensorial, Volatile and Phytochemical Properties of Bovine Milk. Molecules 2020, 25, 26. https://doi.org/10.3390/molecules25010026
Clarke HJ, Griffin C, Rai DK, O’Callaghan TF, O’Sullivan MG, Kerry JP, Kilcawley KN. Dietary Compounds Influencing the Sensorial, Volatile and Phytochemical Properties of Bovine Milk. Molecules. 2020; 25(1):26. https://doi.org/10.3390/molecules25010026
Chicago/Turabian StyleClarke, Holly J., Carol Griffin, Dilip K. Rai, Tom F. O’Callaghan, Maurice G. O’Sullivan, Joseph P. Kerry, and Kieran N. Kilcawley. 2020. "Dietary Compounds Influencing the Sensorial, Volatile and Phytochemical Properties of Bovine Milk" Molecules 25, no. 1: 26. https://doi.org/10.3390/molecules25010026