Isolation of Myricitrin and 3,5-di-O-Methyl Gossypetin from Syzygium samarangense and Evaluation of their Involvement in Protecting Keratinocytes against Oxidative Stress via Activation of the Nrf-2 Pathway

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
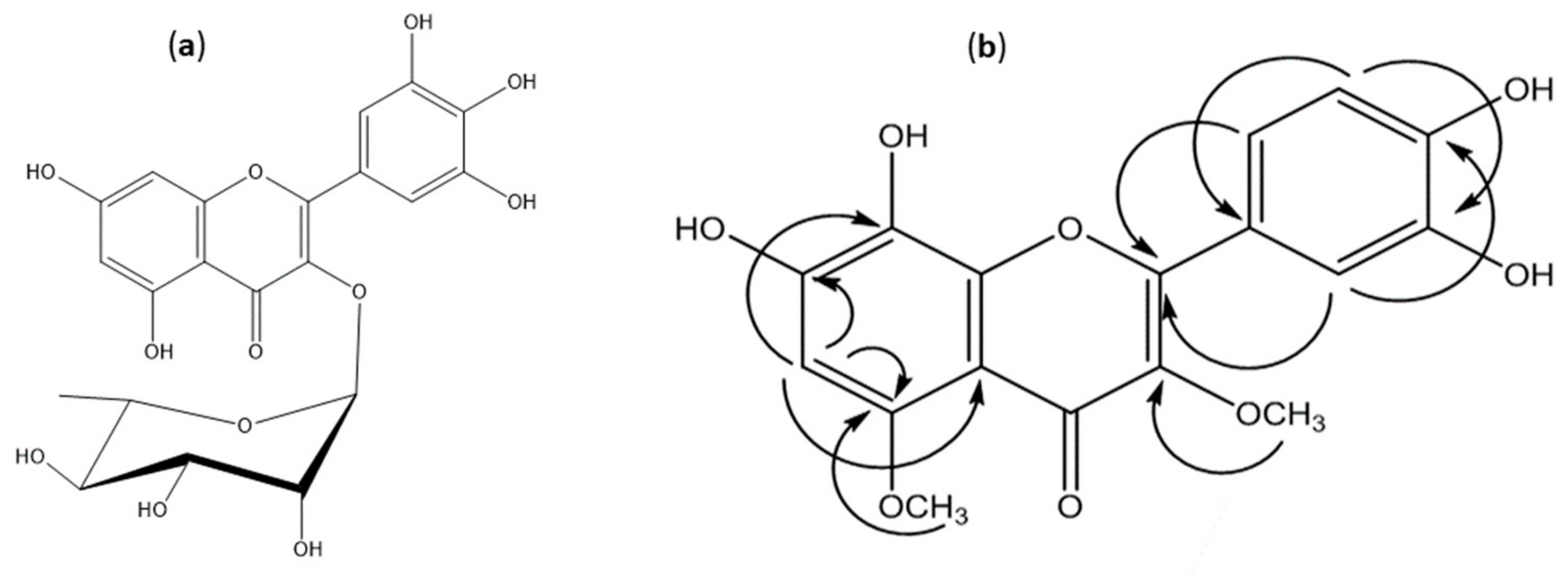
2.1. Compound Isolation
2.1.1. Myricitrin (Myricetin-3-O-α-rhamnoside) (Compound 1)
2.1.2. 7,8,3′,4′-Tetrahydroxy-3,5-dimethoxyflavone (3,5-di-O-Methyl Gossypetin) (Compound 2)
2.2. In Vitro Antioxidant Activity
2.3. Antioxidant Activity in a Cell-Based Model
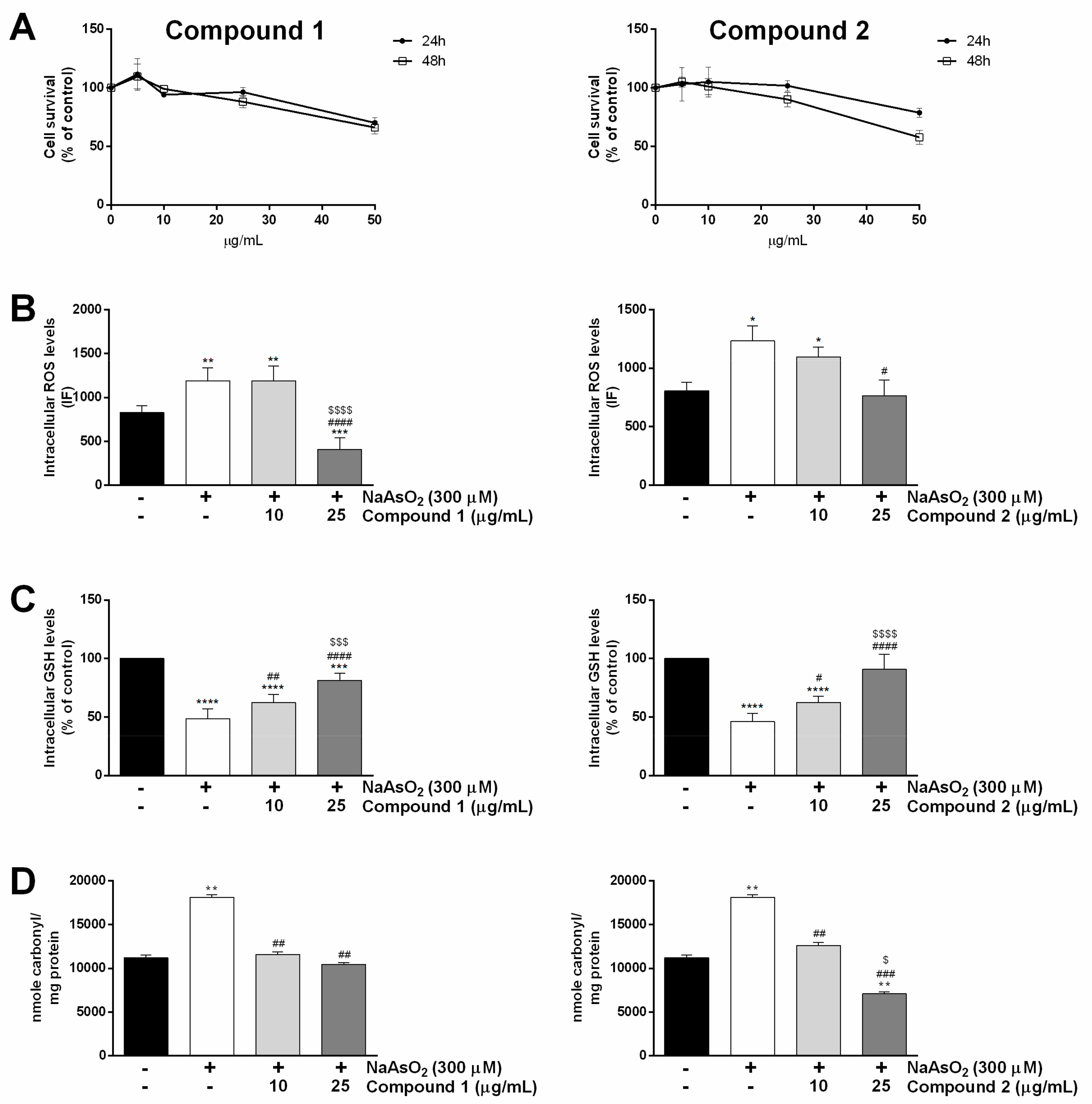
2.3.1. Biocompatibility in Human Keratinocytes
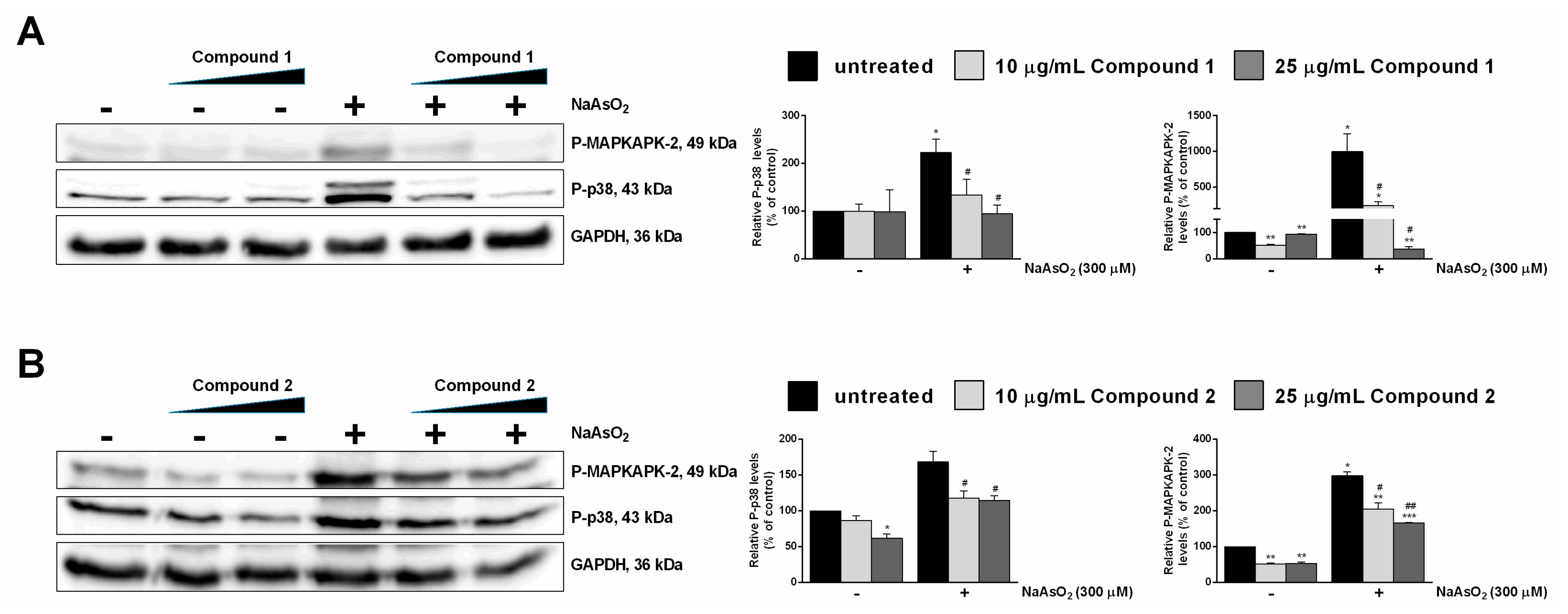
2.3.2. Antioxidative Effect of Compounds 1 and 2 in HaCaT Cells
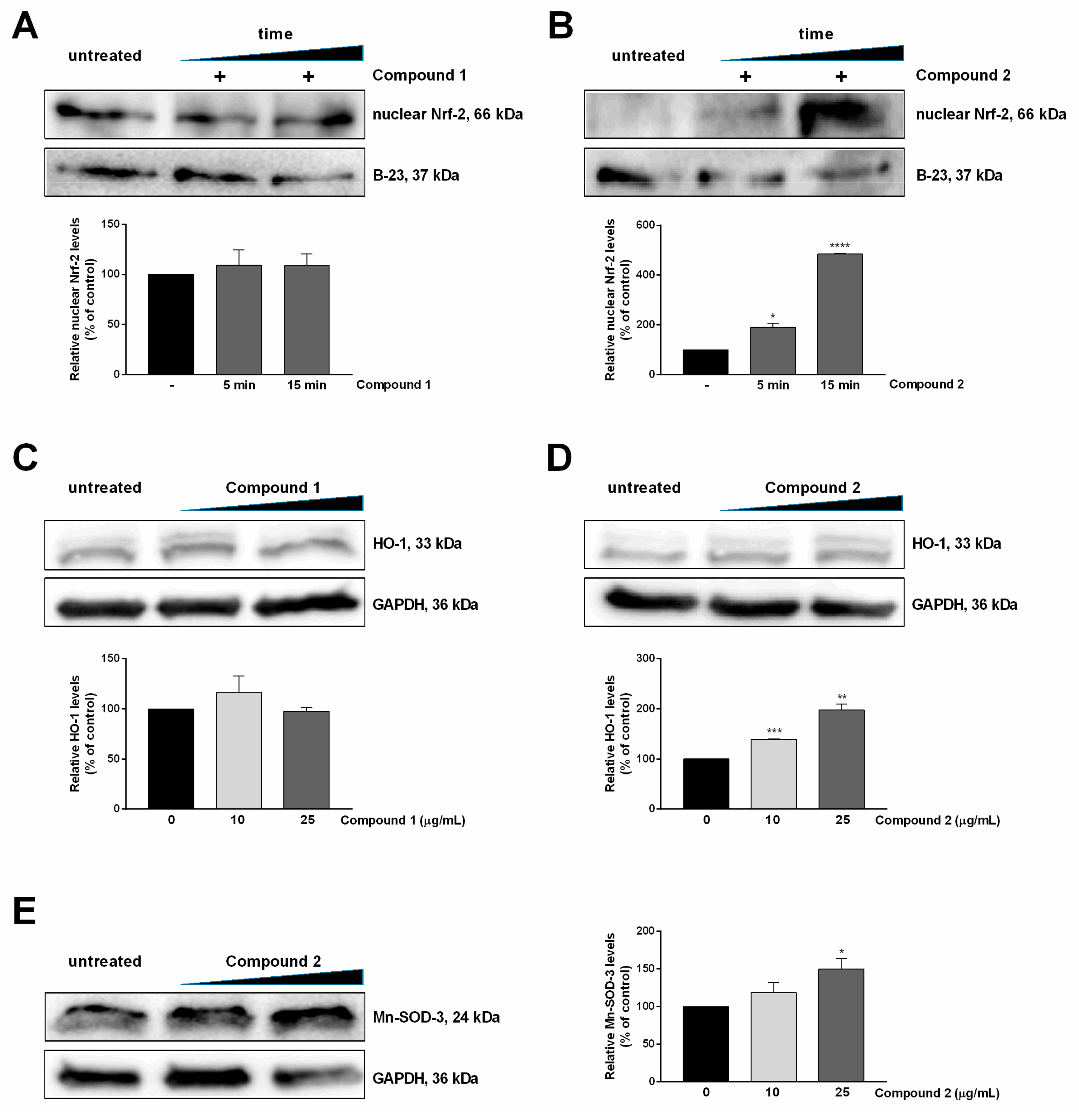
Compound 2 Stimulates Nrf-2 Nuclear Translocation and Nrf2-ARE Transcriptional Activity
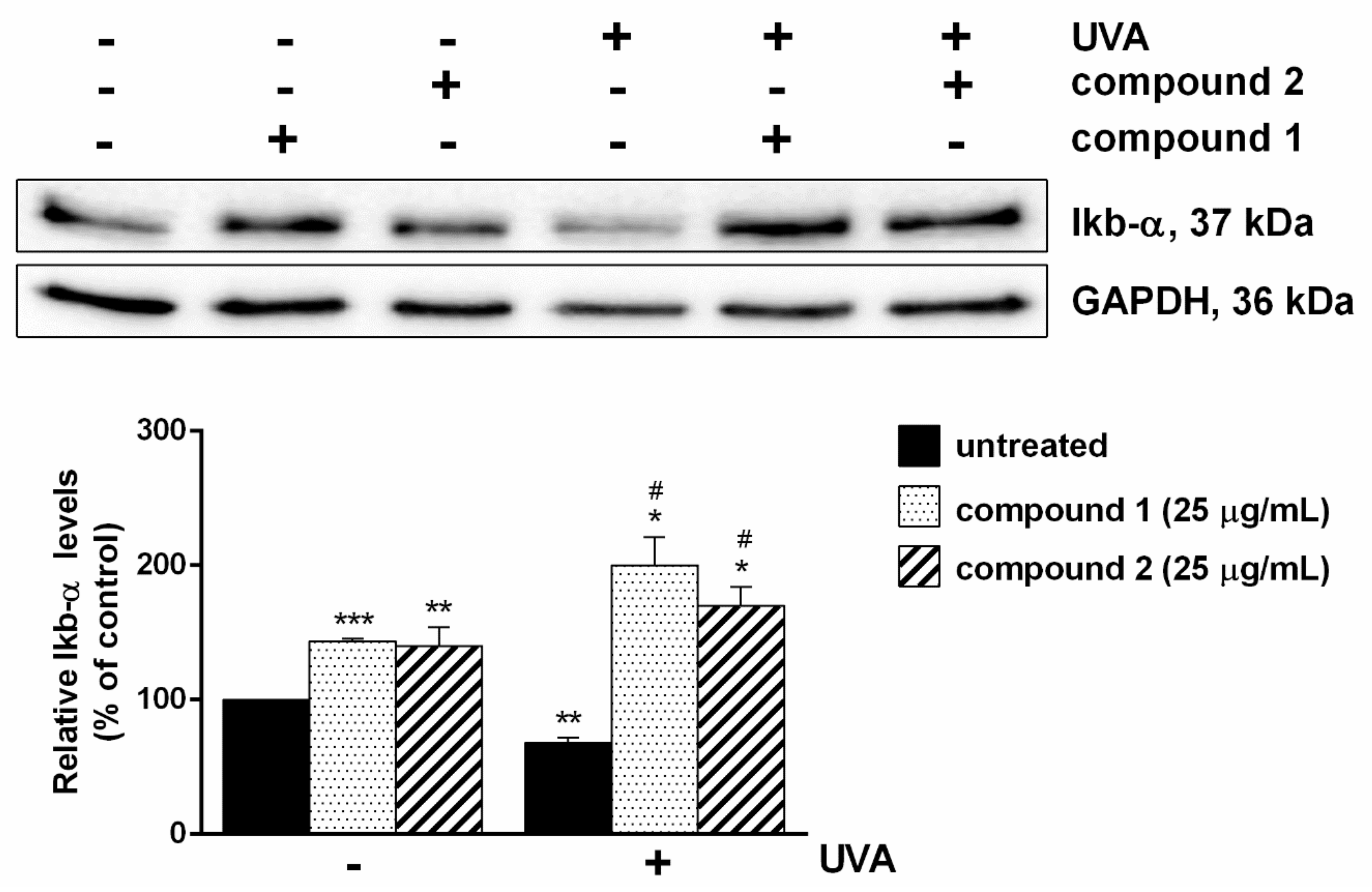
2.4. Pure Compounds as a Shield against UVA-Mediated Inflammation
3. Materials and Methods
3.1. Extraction and Isolation of Compounds
3.2. In Vitro Antioxidant Activity
3.3. Cell Culture
3.4. Induction of Oxidative Stress with Sodium Arsenite
3.5. DCFDA Assay
3.6. DTNB Assay
3.7. DNPH Assay
3.8. Induction of Inflammation by UVA Light Treatment
3.9. Western Blotting Analysis
3.10. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiolog. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A. Mitochondrial metabolism of reactive oxygen species. Biochemistry 2005, 70, 200–214. [Google Scholar] [CrossRef]
- Van Wyk, B.-E.; Wink, M. Phytomedicines, Herbal Drugs, and Poisons; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- Van Wyk, B.-E.; Wink, M. Medicinal Plants of the World; CABI: Wallingford, UK, 2017. [Google Scholar]
- Sobeh, M.; Youssef, F.S.; Esmat, A.; Petruk, G.; El-Khatib, A.H.; Monti, D.M.; Ashour, M.L.; Wink, M. High resolution UPLC-MS/MS profiling of polyphenolics in the methanol extract of Syzygium samarangense leaves and its hepatoprotective activity in rats with CCl4-induced hepatic damage. Food Chem. Toxicol. 2018, 113, 145–153. [Google Scholar] [CrossRef]
- Hu, Y.-K.; Wang, L.; Li, Y.-Y.; Li, M.-J.; Xu, W.; Zhao, Y.; Li, F.; Zhao, Y. Five new triterpenoids from Syzygium samarangense (Bl.) Merr. et Perry. Phytochem. Lett. 2018, 25, 147–151. [Google Scholar] [CrossRef]
- Mabry, T.; Markham, K.; Thomas, M. The Systematic Identification of Flavonoids; Springer Science and Business Media, Springer: New York, NY, USA, 1970. [Google Scholar]
- Barakat, H.H.; El-Raey, M.; Nada, S.A.; Zeid, L.; Nawwar, M. Constitutive phenolics and hepatoprotective activity of Eugenia supra-axillaris leaves. Egypt. J. Chem. 2011, 54, 313–323. [Google Scholar]
- El-Kashak, W.A.; Hamed, A.R.; El-Raey, M.; Elshamy, A.I.; Abd-Ellatef, G.E.F. Antiproliferative, antioxidant and antimicrobial activities of phenolic compounds from Acrocarpus fraxinifolius. J. Chem. Pharmaceut. Res. 2016, 8, 520–528. [Google Scholar]
- Osman, S.M.; El Kashak, W.A.; Wink, M.; El Raey, M.A. New isorhamnetin derivatives from Salsola imbricata Forssk. leaves with distinct anti-inflammatory activity. Pharmacog. Mag. 2016, 12, S47. [Google Scholar]
- Takemura, O.S.; Iinuma, M.; Tosa, H.; Miguel, O.G.; Moreira, E.A.; Nozawa, Y. A flavone from leaves of Arrabidaea chicaf. cuprea. Phytochemistry 1995, 38, 1299–1300. [Google Scholar] [CrossRef]
- Hussein, S.A.; Hashem, A.N.; Seliem, M.A.; Lindequist, U.; Nawwar, M.A. Polyoxygenated flavonoids from Eugenia edulis. Phytochemistry 2003, 64, 883–889. [Google Scholar] [CrossRef]
- Singh, N.; Kumar, D.; Sahu, A.P. Arsenic in the environment: Effects on human health and possible prevention. J. Environ. Biol. 2007, 28, 359. [Google Scholar] [PubMed]
- Hughes, M.F. Arsenic toxicity and potential mechanisms of action. Toxicol. Lett. 2002, 133, 1–16. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Büchter, C.; Ackermann, D.; Havermann, S.; Honnen, S.; Chovolou, Y.; Fritz, G.; Kampkötter, A.; Wätjen, W. Myricetin-mediated lifespan extension in Caenorhabditis elegans is modulated by DAF-16. Int. J. Molec. Sci. 2013, 14, 11895. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Raiola, A.; Del Giudice, R.; Barone, A.; Frusciante, L.; Rigano, M.M.; Monti, D.M. An ascorbic acid-enriched tomato genotype to fight UVA-induced oxidative stress in normal human keratinocytes. J. Photochem. Photobiol. B Biol. 2016, 163, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Pallauf, K.; Duckstein, N.; Hasler, M.; Klotz, L.-O.; Rimbach, G. Flavonoids as putative inducers of the transcription factors Nrf2, FoxO, and PPARγ. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768. [Google Scholar] [CrossRef] [PubMed]
- Hur, W.; Gray, N.S. Small molecule modulators of antioxidant response pathway. Curr. Opin. Chem. Biol. 2011, 15, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.-C.; Tsai, Y.-C.; Huang, P.-J.; Ou, T.-T.; Korivi, M.; Hsu, L.-S.; Chang, S.-H.; Wu, C.-R.; Yang, H.-L. The dermato-protective effects of lucidone from Lindera erythrocarpa through the induction of Nrf2-mediated antioxidant genes in UVA-irradiated human skin keratinocytes. J. Funct. Foods 2015, 12, 303–318. [Google Scholar] [CrossRef]
- Keum, Y.-S. Regulation of Nrf2-mediated phase II detoxification and anti-oxidant genes. Biomolecul. Therapeut. 2012, 20, 144. [Google Scholar] [CrossRef]
- Jones, R.M.; Desai, C.; Darby, T.M.; Luo, L.; Wolfarth, A.A.; Scharer, C.D.; Ardita, C.S.; Reedy, A.R.; Keebaugh, E.S.; Neish, A.S. Lactobacilli modulate epithelial cytoprotection through the Nrf2 pathway. Cell. Rep. 2015, 12, 1217–1225. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Lo, H.-W.; Korivi, M.; Tsai, Y.-C.; Tang, M.-J.; Yang, H.-L. Dermato-protective properties of ergothioneine through induction of Nrf2/ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Rad. Biol. Med. 2015, 86, 102–117. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Köhle, C.; Bock, K.W. Activation of coupled Ah receptor and Nrf2 gene batteries by dietary phytochemicals in relation to chemoprevention. Biochem. Pharmacol. 2006, 72, 795–805. [Google Scholar] [CrossRef]
- Niestroy, J.; Barbara, A.; Herbst, K.; Rode, S.; van Liempt, M.; Roos, P.H. Single and concerted effects of benzopyrene and flavonoids on the AhR and Nrf2-pathway in the human colon carcinoma cell line Caco-2. Toxicol. In Vitro 2011, 25, 671–683. [Google Scholar] [CrossRef]
- Schadich, E.; Hlaváč, J.; Volná, T.; Varanasi, L.; Hajdúch, M.; Džubák, P. Effects of ginger phenylpropanoids and quercetin on Nrf2-ARE pathway in human BJ fibroblasts and HaCaT keratinocytes. BioMed Res. Int. 2016. [Google Scholar] [CrossRef]
- Petruk, G.; Del Giudice, R.; Rigano, M.M.; Monti, D.M. Antioxidants from plants protect against skin photoaging. Oxid. Med. Cell. Longev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zheng, Y. Myricetin protects keratinocyte damage induced by UV through IκB/NFκb signaling pathway. J. Cosmet. Dermatol. 2017, 16, 444–449. [Google Scholar] [CrossRef]
- Sobeh, M.; Braun, M.S.; Krstin, S.; Youssef, F.S.; Ashour, M.L.; Wink, M. Chemical profiling of the essential oils of Syzygium aqueum, Syzygium samarangense and Eugenia uniflora and their discrimination using chemometric analysis. Chem. Biodiv. 2016, 13, 1537–1550. [Google Scholar] [CrossRef]
- Ghareeb, M.A.; Mohamed, T.; Saad, A.M.; Refahy, L.A.-G.; Sobeh, M.; Wink, M. HPLC-DAD-ESI-MS/MS analysis of fruits from Firmiana simplex (L.) and evaluation of their antioxidant and antigenotoxic properties. J. Pharm. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Donadio, G.; Lanzilli, M.; Isticato, R.; Monti, D.M. Alternative use of Bacillus subtilis spores: protection against environmental oxidative stress in human normal keratinocytes. Sci. Rep. 2018, 8, 1745. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Compound 1 | Compound 2 | ||
|---|---|---|---|---|
| δH | δC | δH | δC | |
| 2 | 159.56 | 155.61 | ||
| 3 | 136.44 | 141.29 | ||
| 4 | 179.80 | 176.64 | ||
| 5 | 163.36 | 154.64 | ||
| 6 | 6.20 (d, J = 2.1 Hz) | 100.02 | 6.45 (s) | 97.24 |
| 7 | 166.27 | 152.93 | ||
| 8 | 6.36 (d, J = 2.1 Hz) | 94.88 | 127.47 | |
| 8a | 158.68 | 146.94 | ||
| 4a | 105.95 | 108.79 | ||
| 3-OCH3 | 3.76 (s) | 60.36 | ||
| 5-OCH3 | 3.83 (s) | 56.56 | ||
| 1′ | 122.07 | 123.42 | ||
| 2′ | 6.95 (s) | 109.70 | 7.70 (d, J = 2.0 Hz) | 116.76 |
| 3′ | 147.01 | 146.39 | ||
| 4′ | 138.05 | 149.58 | ||
| 5′ | 147.01 | 6.89 (d, J = 8.4) | 116.42 | |
| 6′ | 6.95 (s) | 109.70 | 7.61 (dd, J = 8.4 & 2.0 Hz) | 122.45 |
| 1′′ | 5.23 (d, J = 1.5 Hz) | 103.78 | ||
| 2′′ | 4.22 (dd, J = 1.5 & 3.4 Hz) | 72.04 | ||
| 3′′ | 3.79 (dd, J = 9.5 & 3. 4 Hz) | 72.19 | ||
| 4′′ | 3.35 (t, J = 9.5 Hz) | 73.51 | ||
| 5′′ | 3.52 (dd, J = 9.8 & 6.2 Hz) | 72.28 | ||
| CH3-rh | 0.98 (d, J = 6.2 Hz) | 17.82 | ||
| DPPH | FRAP | |
|---|---|---|
| Sample | (EC50 µg/mL) | (mM FeSO4 Equivalent/mg Sample) |
| Compound 1 | 3.21 | 22.9 |
| Compound 2 | 3.89 | 21.08 |
| Total extract [5] | 5.80 | 10 |
| Ascorbic acid | 2.94 | - |
| Quercetin | - | 23.18 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobeh, M.; Petruk, G.; Osman, S.; El Raey, M.A.; Imbimbo, P.; Monti, D.M.; Wink, M. Isolation of Myricitrin and 3,5-di-O-Methyl Gossypetin from Syzygium samarangense and Evaluation of their Involvement in Protecting Keratinocytes against Oxidative Stress via Activation of the Nrf-2 Pathway. Molecules 2019, 24, 1839. https://doi.org/10.3390/molecules24091839
Sobeh M, Petruk G, Osman S, El Raey MA, Imbimbo P, Monti DM, Wink M. Isolation of Myricitrin and 3,5-di-O-Methyl Gossypetin from Syzygium samarangense and Evaluation of their Involvement in Protecting Keratinocytes against Oxidative Stress via Activation of the Nrf-2 Pathway. Molecules. 2019; 24(9):1839. https://doi.org/10.3390/molecules24091839
Chicago/Turabian StyleSobeh, Mansour, Ganna Petruk, Samir Osman, Mohamed A. El Raey, Paola Imbimbo, Daria Maria Monti, and Michael Wink. 2019. "Isolation of Myricitrin and 3,5-di-O-Methyl Gossypetin from Syzygium samarangense and Evaluation of their Involvement in Protecting Keratinocytes against Oxidative Stress via Activation of the Nrf-2 Pathway" Molecules 24, no. 9: 1839. https://doi.org/10.3390/molecules24091839