Morphology and Molecular Identification of Twelve Commercial Varieties of Kiwifruit


Abstract
:1. Introduction
2. Results
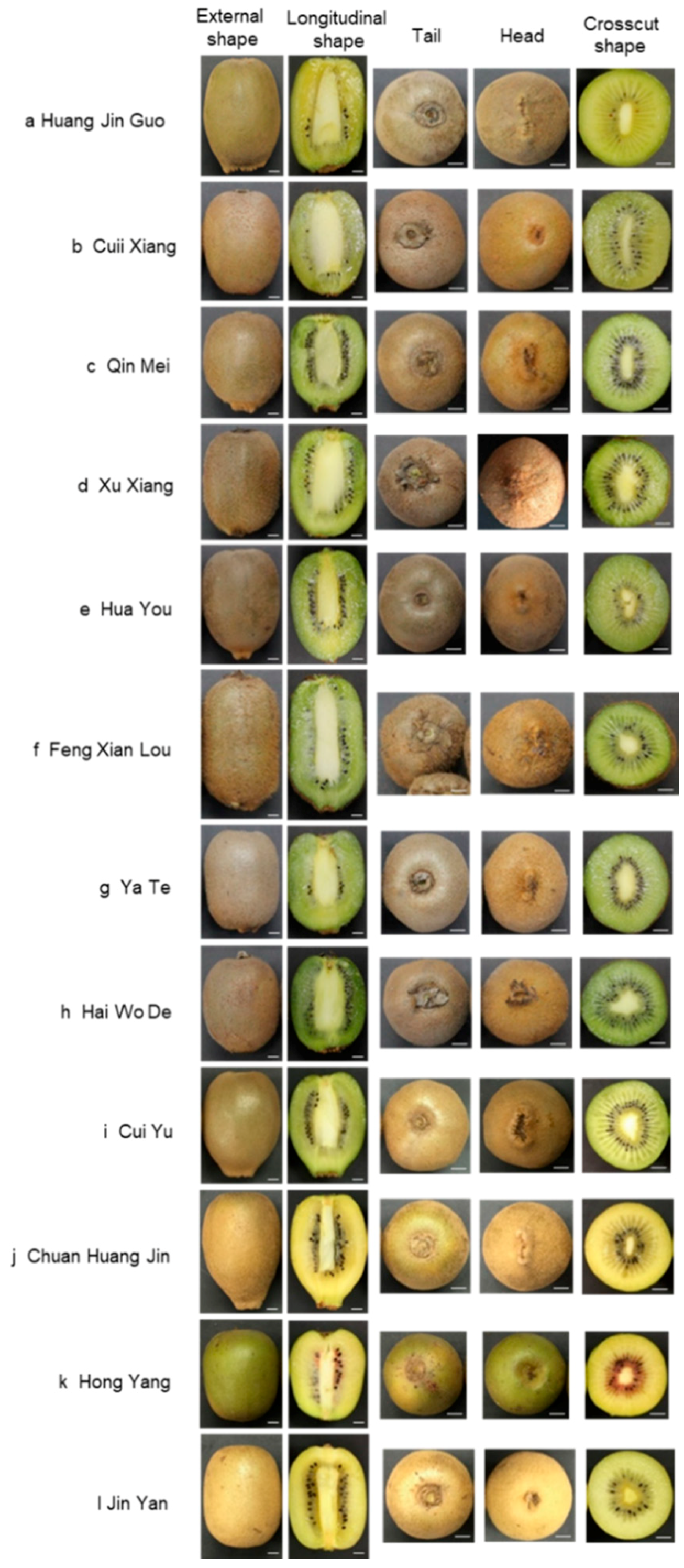
2.1. Morphological Identification
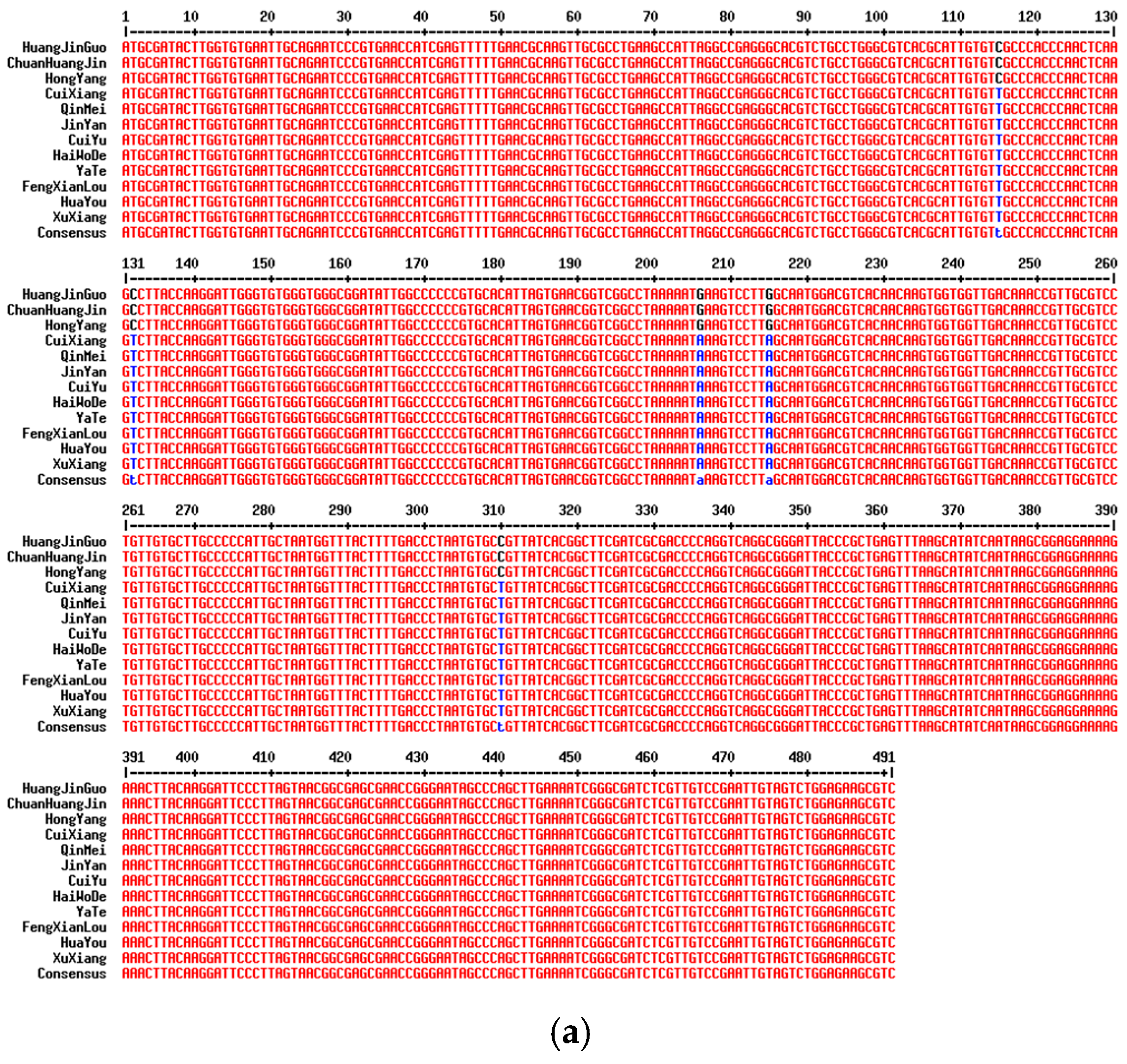
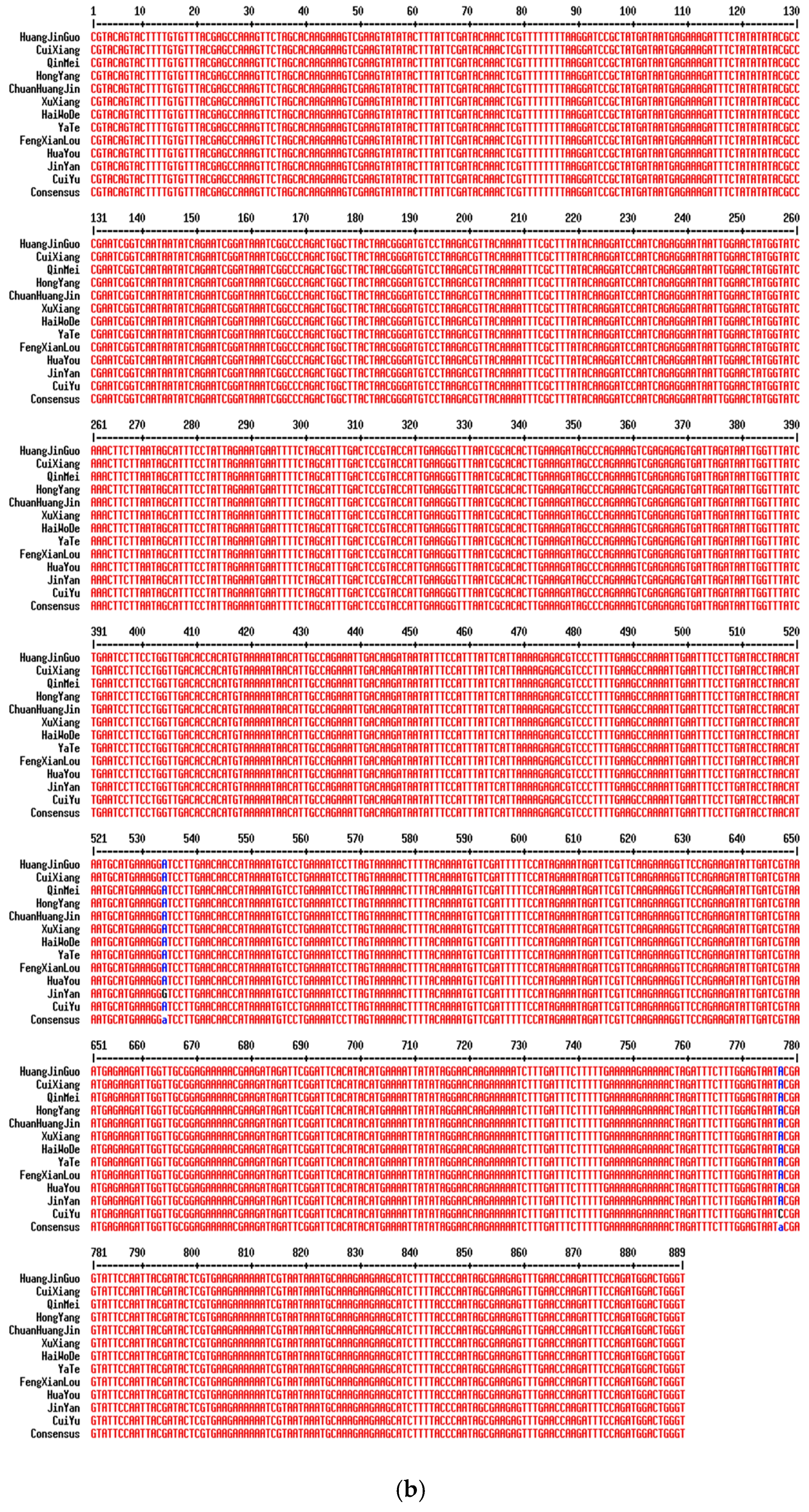
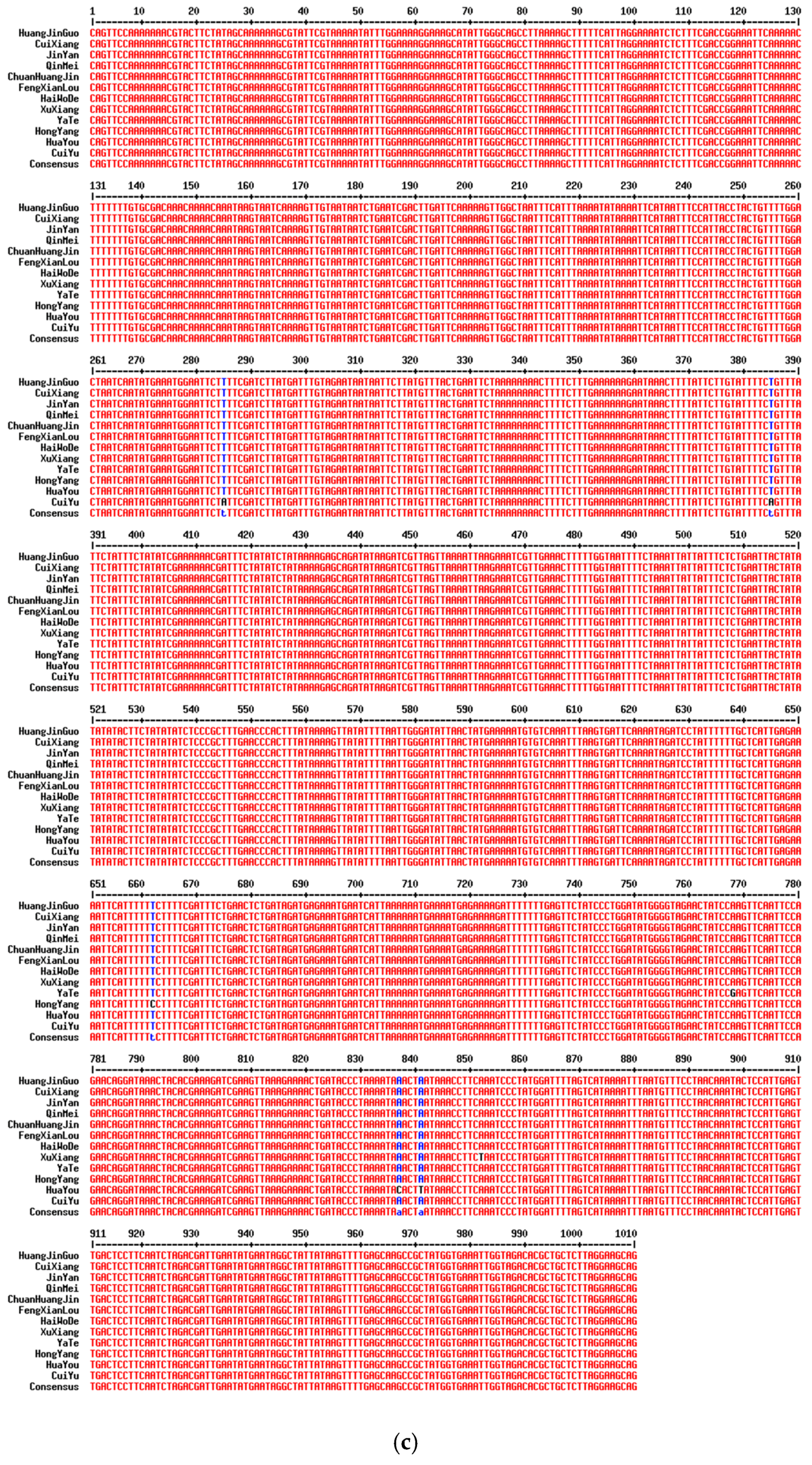
2.2. Analysis of Variable Sites in Different Commercial Varieties
2.3. K2P Genetic Distance Analysis
2.4. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Samples Collection and DNA Extraction
4.2. PCR Amplification, Cloning, and Sequencing
4.3. Sequence Alignments and Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hunter, D.C.; Greenwood, J.; Zhang, J.L.; Skinner, M.A. Antioxidant and ‘Natural Protective’ Properties of Kiwifruit. Curr. Top. Med. Chem. 2011, 11, 1811–1820. [Google Scholar] [CrossRef] [PubMed]
- Analysis of global kiwifruit yield and production area distribution. Available online: http://www.chyxx.com/industry/201602/389018.html (accessed on 22 December 2018).
- Garcia, C.V.; Quek, S.Y.; Stevenson, R.J.; Winz, R.A. Characterisation of bound volatile compounds of a low flavour kiwifruit species: Actinidia eriantha. Food Chem. 2012, 134, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, I.; Yamashita, Y.; Yamanaka, M.; Shimohashi, A.; Fukuda, T.; Oota, T. Varietal difference in vitamin C content in the fruit of kiwifruit and other Actinidia species. J. Agric. Food Chem. 2004, 52, 5472–5475. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B-Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ding, C.B.; Zhang, L.; Yang, R.W.; Zhou, Y.H.; Tang, L. Identification of the genus Epimedium with DNA barcodes. J. Med. Plants Res. 2011, 5, 6413–6417. [Google Scholar] [CrossRef]
- Liu, J.X.; Shi, L.C.; Han, J.P.; Li, G.; Lu, H.; Hou, J.Y.; Zhou, X.T.; Meng, F.Y.; Downie, S.R. Identification of species in the angiosperm family Apiaceae using DNA barcodes. Mol. Ecol. Resour. 2014, 14, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; Fazekas, A.J.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar]
- Yao, H.; Song, J.Y.; Ma, X.Y.; Liu, C.; Li, Y.; Xu, H.X.; Han, J.P.; Duan, L.S.; Chen, S.L. Identification of Dendrobium Species by a Candidate DNA Barcode Sequence: The Chloroplast psbA-trnH Intergenic Region. Planta Med. 2009, 75, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Xie, C.X.; Liu, C.; Song, J.Y.; Yao, H.; Luo, K.; Zhu, Y.J.; Gao, T.; Pang, X.H.; Qian, J.; et al. Species Identification of Medicinal Pteridophytes by a DNA Barcode Marker, the Chloroplast psbA-trnH Intergenic Region. Biol. Pharm. Bull. 2010, 33, 1919–1924. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, A.J.; Burgess, K.S.; Kesanakurti, P.R.; Graham, S.W.; Newmaster, S.G.; Husband, B.C.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C. Multiple multilocus DNA barcodes from the plastid genome discriminate plant species equally well. PLoS ONE 2008, 3, e2802. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhan, A.; Cao, L.; Meng, F.; Xu, W. Selection of a marker gene to construct a reference library for wetland plants, and the application of metabarcoding to analyze the diet of wintering herbivorous waterbirds. PeerJ 2016, 4, e2345. [Google Scholar] [CrossRef] [PubMed]
- Li, M.G.; Wunder, J.; Bissoli, G.; Scarponi, E.; Gazzani, S.; Barbaro, E.; Saedler, H.; Varotto, C. Development of COS genes as universally amplifiable markers for phylogenetic reconstructions of closely related plant species. Cladistics 2008, 24, 727–745. [Google Scholar] [CrossRef]
- Bruni, I.; De Mattia, F.; Galimberti, A.; Galasso, G.; Banfi, E.; Casiraghi, M.; Labra, M. Identification of poisonous plants by DNA barcoding approach. Int. J. Leg. Med. 2010, 124, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Wang, Z.L.; Lin, L.J.; Lin, L.; Li, W.; Yuan, D.M. Molecular Phylogenetic Analysis of Wild Cerasus Plants in South China. J. Nucl. Agric. Sci. 2018, 32, 2126–2134. [Google Scholar]
- Lee, S.C.; Wang, C.H.; Yen, C.E.; Chang, C. DNA barcode and identification of the varieties and provenances of Taiwan’s domestic and imported made teas using ribosomal internal transcribed spacer 2 sequences. J. Food Drug Anal. 2017, 25, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Enan, M.R.; Ahmed, A. Cultivar-level phylogeny using chloroplast DNA barcode psbK-psbI spacers for identification of Emirati date palm (Phoenix dactylifera L.) varieties. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Suokas, M.; Haggman, H. Novel approaches based on DNA barcoding and high-resolution melting of amplicons for authenticity analyses of berry species. Food Chem. 2010, 123, 494–500. [Google Scholar] [CrossRef]
- He, Y.; Hou, P.; Fan, G.; Song, Z.; Arain, S.; Shu, H.; Tang, C.; Yue, Q.H.; Zhang, Y. Authentication of Angelica anomala Ave-Lall cultivars through DNA barcodes. Mitochondrial DNA 2012, 23, 100–105. [Google Scholar] [CrossRef] [PubMed]
- de Paula Careta, F.; Paneto, G.G. Recent patents on high-throughput single nucleotide polymorphism (SNP) genotyping methods. Recent Pat. DNA Gene Seq. 2012, 6, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Nagasaki, H.; Yonemaru, J.; Ebana, K.; Nakajima, M.; Shibaya, T.; Yano, M. Fine definition of the pedigree haplotypes of closely related rice cultivars by means of genome-wide discovery of single-nucleotide polymorphisms. BMC Genom. 2010, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jiang, C.; Liu, L.; Yu, S.; Cui, Z.; Chen, M.; Lin, S.; Wang, S.; Huang, L. Convenient, sensitive and high-throughput method for screening botanic origin. Sci. Rep. 2014, 4, 5395. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; Kim, M.K.; Kim, Y.J.; Lee, H.N.; Jin, H.Z.; Chen, J.; Yang, D.C. Molecular authentication of the Oriental medicines Pericarpium Citri Reticulatae and Citri Unshius Pericarpium using SNP markers. Gene 2012, 494, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Maiden, M.C.J.; van Rensburg, M.J.J.; Bray, J.E.; Earle, S.G.; Ford, S.A.; Jolley, K.A.; McCarthy, N.D. MLST revisited: The gene-by-gene approach to bacterial genomics. Nat. Rev. Microbiol. 2013, 11, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed]
- Ferri, G.; Alu, M.; Corradini, B.; Licata, M.; Beduschi, G. Species Identification Through DNA “Barcodes”. Genet. Test. Mol. Biomark. 2009, 13, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haider, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Philos. Trans. R. Soc. B-Biol. Sci. 2005, 360, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Taxonomy—Wanted: A barcode for plants. Science 2007, 318, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Newmaster, S.G.; Fazekas, A.J.; Steeves, R.A.D.; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Resour. 2008, 8, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Cowan, R.S.; Hollingsworth, P.M.; van den Berg, C.; Madrinan, S.; Petersen, G.; Seberg, O.; Jorgsensen, T.; Cameron, K.M.; Carine, M. A proposal for a standardised protocol to barcode all land plants. Taxon 2007, 56, 295–299. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.G.; Jiao, K.L.; Zhu, Y.J.; Wang, H.F.; Jiang, M.Y.; Wang, H.Z. Molecular identification of species of Physalis (Solanaceae) using a candidate DNA barcode: The chloroplast psbA-trnH intergenic region. Genome 2018, 61, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.C.; Liao, B.S.; Song, J.Y.; Pang, X.H.; Han, J.P.; Chen, S.L. A fast SNP identification and analysis of intraspecific variation in the medicinal Panax species based on DNA barcoding. Gene 2013, 530, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Duan, Z.; Yang, J.; Ma, X.; Zhan, R.; Xu, H.; Chen, W. SNP typing for germplasm identification of Amomum villosum Lour. Based on DNA barcoding markers. PLoS ONE 2014, 9, e114940. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.P.; Tranel, P.J. Identification and Validation of Amaranthus Species-Specific SNPs within the ITS Region: Applications in Quantitative Species Identification. Crop. Sci. 2018, 58, 304–311. [Google Scholar] [CrossRef]
- Huang, S.X.; Ding, J.; Deng, D.J.; Tang, W.; Sun, H.H.; Liu, D.Y.; Zhang, L.; Niu, X.L.; Zhang, X.; Meng, M. Draft genome of the kiwifruit Actinidia chinensis. Nat. Commun. 2013, 4, 2640. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, S.M.; Crowhurst, R.; Hilario, E.; Nardozza, S.; Fraser, L.; Peng, Y.Y.; Gunaseelan, K.; Simpson, R.; Tahir, J.; Deroles, S.C. A manually annotated Actinidia chinensis var. chinensis (kiwifruit) genome highlights the challenges associated with draft genomes and gene prediction in plants. BMC Genom. 2018, 19, 257. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Qi, X.; Chen, J.; Sun, L.; Zhong, Y.; Fang, J.; Hu, C. The complete chloroplast genome sequence of Actinidia arguta using the PacBio RS II platform. PLoS ONE 2018, 13, e0197393. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tang, P.; Li, Z.; Li, D.; Liu, Y.; Huang, H. The First Complete Chloroplast Genome Sequences in Actinidiaceae: Genome Structure and Comparative Analysis. PLoS ONE 2015, 10, e0129347. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G.J.N. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
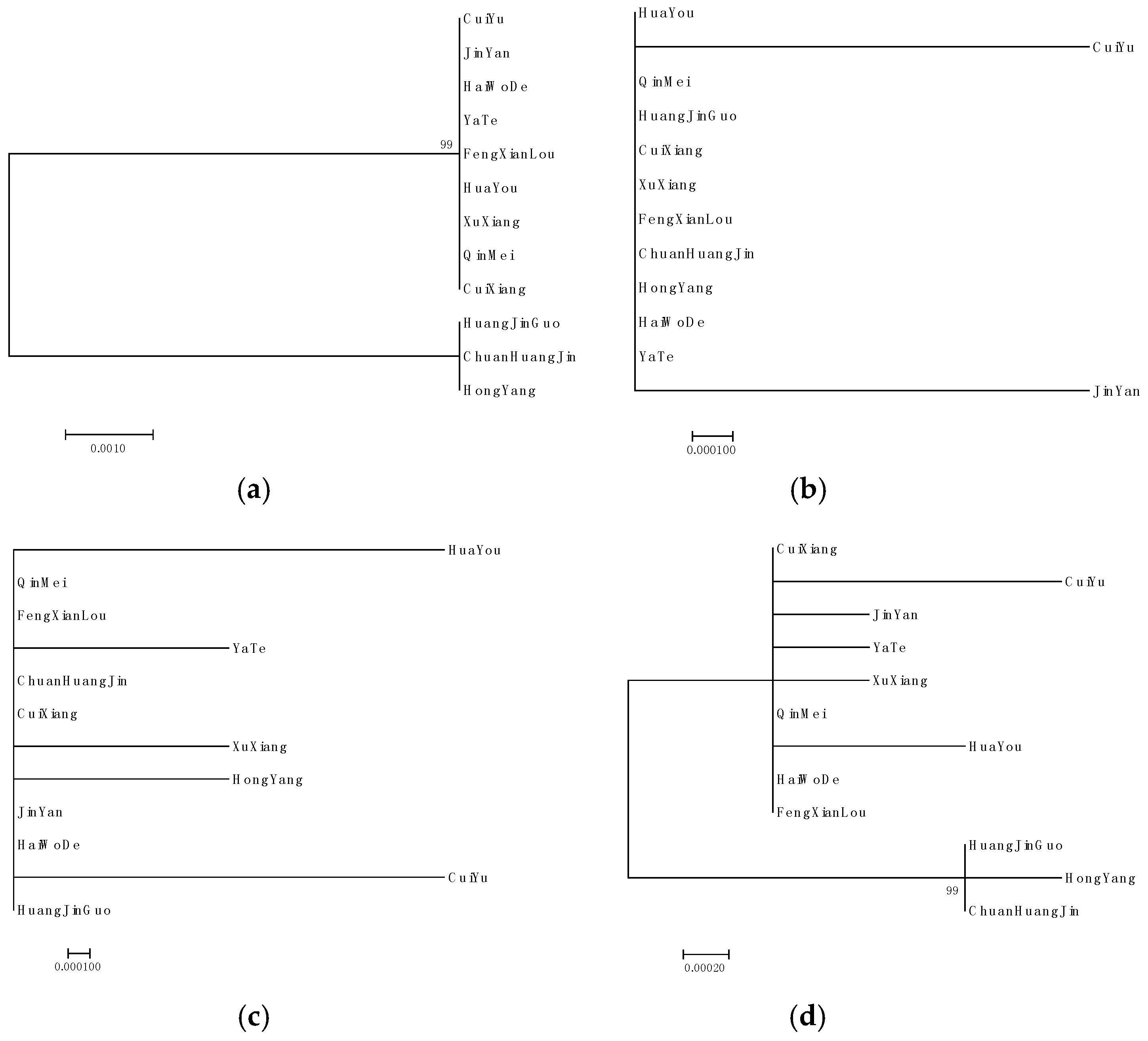{kind=link}
| Variety | SNP Site Information (bp) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ITS2 | matK | rpl32 | ||||||||||||
| 115 | 132 | 206 | 215 | 310 | 534 | 777 | 285 | 385 | 662 | 768 | 837 | 841 | 852 | |
| HuangJinGuo | C | C | G | G | C | A | A | T | T | T | A | A | A | A |
| ChuanHuangJin | C | C | G | G | C | A | A | T | T | T | A | A | A | A |
| HongYang | C | C | G | G | C | A | A | T | T | C | A | A | A | A |
| CuiiXiang | T | T | A | A | T | A | A | T | T | T | A | A | A | A |
| QinMei | T | T | A | A | T | A | A | T | T | T | A | A | A | A |
| JinYan | T | T | A | A | T | G | A | T | T | T | A | A | A | A |
| CuiYu | T | T | A | A | T | A | C | A | A | T | A | A | A | A |
| HaiWoDe | T | T | A | A | T | A | A | T | T | T | A | A | A | A |
| YaTe | T | T | A | A | T | A | A | T | T | T | G | A | A | A |
| FengXianLou | T | T | A | A | T | A | A | T | T | T | A | A | A | A |
| HuaYou | T | T | A | A | T | A | A | T | T | T | A | C | T | A |
| XuXiang | T | T | A | A | T | A | A | T | T | T | A | A | A | T |
| Gene Amplification Region | Primer Name | Sequence (5′ →3′) | Length of Amplified Fragment | Amplification Procedure |
|---|---|---|---|---|
| rbcL | rbcL-F rbcL-R | ATGTCACCACAAACAGA TCGCATGTACCTGCAGTA | 743 bp | 94 °C 5 min; [94 °C 30S, 54.5 °C 30S, 72 °C 45S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| matK | matK-F matK-R | CGTACAGTACTTTTGTGTTTAC ACCCAGTCCATCTGGAAATC | 889 bp | 94 °C 5 min; [94 °C 30S, 56 °C 30S, 72 °C 55S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| psbA-trnH | psbA-trnH-F psbA-trnH-R | TATGCATGAACGTAATGCT GCATGGTGGATTCACAAT | 502 bp | 94 °C 5 min; [94 °C 30S, 55 °C 30S, 72 °C 30S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| ITS2 | ITS2-F ITS2-R | ATGCGATACTTGGTGTG GACGCTTCTCCAGACTA | 491 bp | 94 °C 5 min; [94 °C 30S, 55 °C 30S, 72 °C 30S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| rpoB | rpoB-F rpoB-R | AAGTGCATTGTTGGAACTGG CCCAGCATCACAATTCC | 512 bp | 94 °C 5 min; [94 °C 30S, 55 °C 30S, 72 °C 35S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| rpoC1 | rpoC1-F rpoC1-R | CAAAGAGGGAAGAT TAAGCATATCTTGAGT | 529 bp | 94 °C 5 min; [94 °C 30S, 54.5 °C 30S, 72 °C 40S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| rpl32_trnL(UAG) | rpl32 trnL(UAG) | CAGTTCCAAAAAAACGTACTTC CTGCTTCCTAAGAGCAGCGT | 1010 bp | 94 °C 5 min; [94 °C 30S, 57.8 °C 30S, 72 °C 45S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| trnL | trnL-F trnL-R | CGAAATCGGTAGACGCTACG CCATTGAGTCTCTGCACCTATC | 193 bp | 94 °C 5 min; [94 °C 30S, 56.5 °C 30S, 72 °C 30S]* 35 cycles; 72 °C 10 min; 4 °C 10 min |
| ycf1b | ycf1b-F ycf1b-R | TCTCGACGAAAATCAGATTGTTGTGAAT ATACATGTCAAAGTGATGGAAAA | No result | 94 °C 5 min; [94 °C 30S, 56.5 °C 30S, 72 °C 45S]* 35 cycles; 72 °C 10 min; 4 °C 10 min 56.5 °C |
| At103 | At103-F At103-R | CTTCAAGCCMAAGTTCATCTTCTA ATCATTGAGGTACATNGTMACATA | No result | 94 °C 5 min; [94 °C 30S, 54 °C 30S, 72 °C 45S]* 35 cycles; 72 °C 10 min; 4 °C 10 min 56.5 °C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Q.; Zhang, H.; Yan, F.; Yan, C.; Wei, S.; Lai, J.; Wang, Y.; Zhang, B. Morphology and Molecular Identification of Twelve Commercial Varieties of Kiwifruit. Molecules 2019, 24, 888. https://doi.org/10.3390/molecules24050888
Xie Q, Zhang H, Yan F, Yan C, Wei S, Lai J, Wang Y, Zhang B. Morphology and Molecular Identification of Twelve Commercial Varieties of Kiwifruit. Molecules. 2019; 24(5):888. https://doi.org/10.3390/molecules24050888
Chicago/Turabian StyleXie, Qiaoli, Hongbo Zhang, Fei Yan, Chunxia Yan, Shuguang Wei, Jianghua Lai, Yunpeng Wang, and Bao Zhang. 2019. "Morphology and Molecular Identification of Twelve Commercial Varieties of Kiwifruit" Molecules 24, no. 5: 888. https://doi.org/10.3390/molecules24050888