1. Introduction
Herbicides have been used widely in the field of agriculture to kill weed pests and increase crop yield production. However, most of the available herbicides are synthetic chemical compounds, causing serious pollution of the ecological system due to their large-scale usage and difficult degradation [
1,
2]. Meanwhile, herbicide resistance is increasing in weed control after a long history of the application of existing commercial herbicides [
3,
4,
5]. Therefore, the exploration of new herbicidal agents is of increasing importance.
To date, more than 400 phytotoxins have been isolated and structures identified from microbial sources, which possess different chemical characterizations [
6,
7]. With great structural diversity and novel mechanisms compared to synthetic herbicides, naturally occurring compounds from microbes have been ideal candidates for new herbicides. They have been used as natural herbicides directly, as leading compounds, or for simultaneous applications with a low dose of synthetic herbicides to achieve a better treatment outcome [
8]. Several compounds have been the product of successful commercial herbicides, or the prototype of new herbicides against several kinds of weeds worldwide. Glufosinate, also known as phosphinothricin, is produced by several species of
Streptomyces bacteria. It is one of the most successful commercial herbicides because it is a broad-spectrum herbicide, and is widely used to control important weeds such as
Ipomoea aquatica,
Sesbania aculeata (Wild.) Pers.,
Polygonum pensylvanicum and
Cyperus esculentus [
9,
10,
11,
12]. Hydantocidin, a spironucleoside from
Streptomyces hygroscopicus, and its synthetic analogues have been patented as herbicides [
13,
14].
Cichorine is a novel phytotoxin active against knapweed, corn, and soybeans. It was discovered from several fungi including
A. nidulans and
Alternaria cichorii, which produces foliar blight in the important pest Russian knapweed. We formulated the hypothesis that cichorine is an important lead compound for new herbicide development because its compact, functionalized isoindolin-1-one framework is extremely attractive in the field of drug discovery [
15]. Therefore, it is highly important to discover structural analogues of cichorine for new herbicides. In our previous research, a cluster of seven genes including non-reducing polyketide synthase AN6448 was confirmed as a requirement for the biosynthesis of cichorine in the strain
A. nidulans. Interestingly, cichorine produced by this pathway could be the precursor to other interesting compounds, as the deletion of AN6448 led to no production of the NRPS peptide aspercryptin. This provides us with a powerful platform to discover new analogues of cichorine with good herbicide activity. The occurrence of many tailoring enzymes in the strain
A. nidulans makes us believe that some cichorine-derived compounds could be biologically synthesized in this fungus [
16]. In order to isolate and identify the cichorine derivatives, we selected the engineered
A. nidulans LO8030, in which the gene clusters responsible for the biosynthesis of most of the known secondary metabolites in the wild type were deleted to reduce metabolite background in
A. nidulans and minimize the rediscovery of known compounds. The gene deletion of the PKS gene AN6448∆ (Δ
pkbA) from LO8030 was conducted because the PKS gene is responsible for the first step in the biosynthesis of cichorine, and therefore it is crucial for the biosynthesis of any cichorine-derived compound. High-performance liquid chromatography coupled with a UV detector (HPLC-UV) profile analysis of the ethyl acetate (EtOAc) extract from the parental and Δ
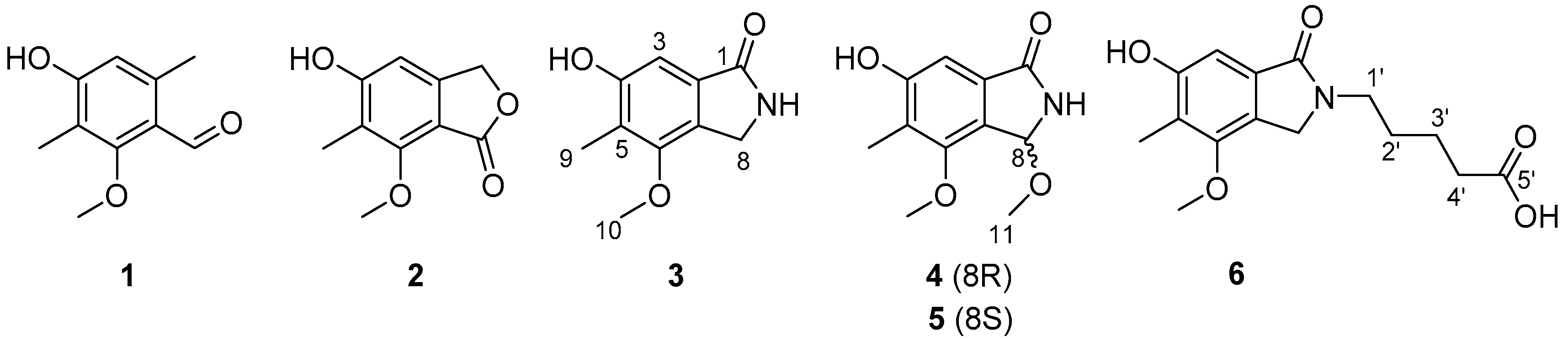
pkbA strains resulted in the discovery of six compounds, including 3,6-dimethyl-4-hydroxy-2-methoxybenzaldehyde (
1) [
17], nidulol (
2) [
18], cichorine (
3) [
15], 8-methoxycichorine (
4), 8-
epi-methoxycichorine (
5), and
N-(4’-carboxybutyl) cichorine (
6). Among the metabolites, compounds (
4), (
5), and (
6) were determined as three new cichorine analogues with potent phytotoxicity. Here, we report the isolation, purification, structure determination, and biological assay of the isolate compounds by pathway inactivation.
2. Results and Discussion
The AN6448 deletion mutant was generated by the homologous transformation of the
A. nidulans strain LO8030. The AN6448 gene was replaced by a pyrG selectable marker from
A. fumigatus 293, as seen in
Figure S1A. The resulting transformants were confirmed by diagnostic PCR, as shown in
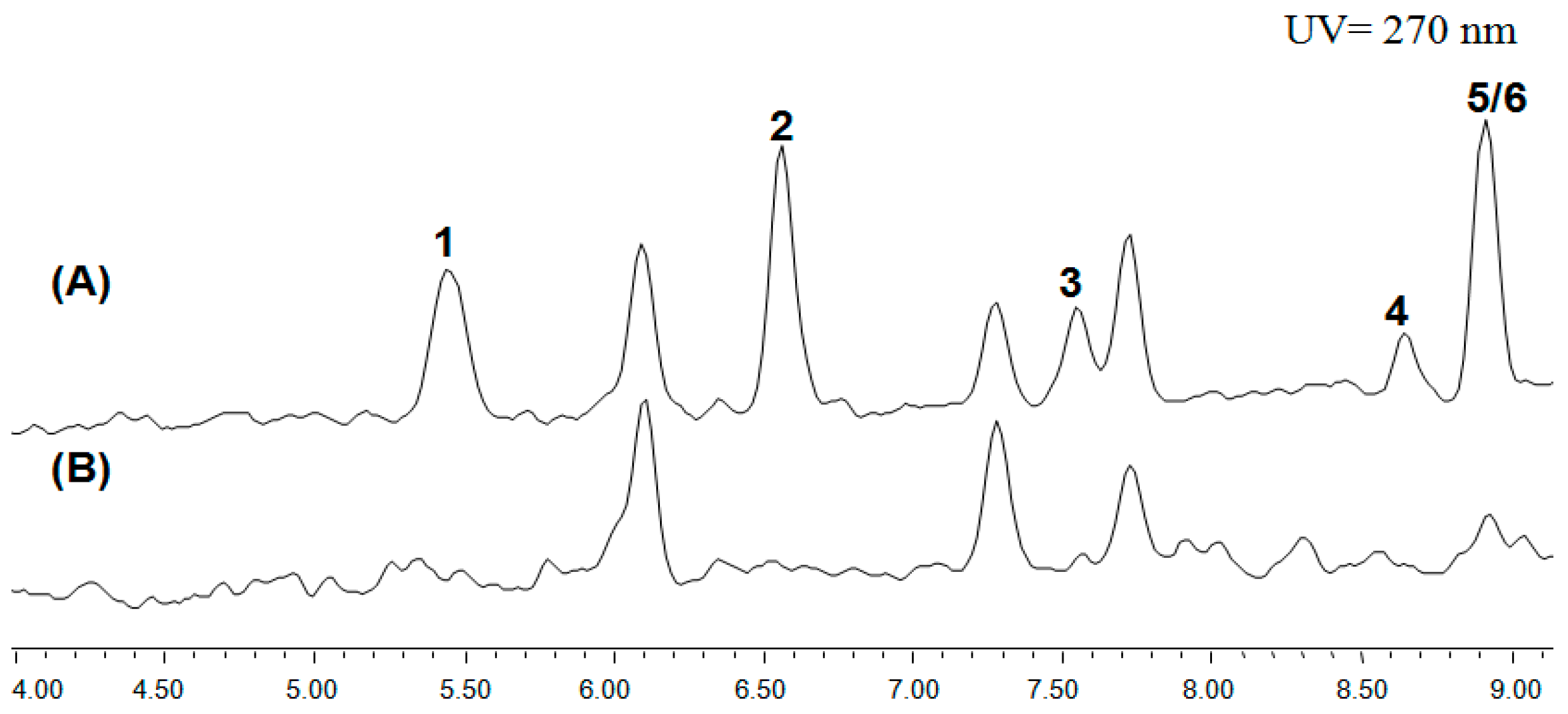
Figure S1B. The HPLC results showed that the cichorine was drastically eliminated in the positive deletion mutant in comparison with the wild-type LO8030, which was designated as the control in
Figure 1. This was consistent with the previous study, which confirmed the involvement of the AN6448 gene in cichorine biosynthesis. Consequently, we discovered six compounds (
1–
6), three of which are new ones (
4–
6) in the ΔAN6448 mutant, as shown in
Figure 2.
The molecular formula of 8-methoxycichorine (
4) was deduced to be C
11H
13NO
4 by high-resolution electrospray ionization (HRESIMS) analysis. The six degrees of unsaturation indicated the presence of one aromatic ring. The UV spectrum showed the absorption at 215, 259, and 298 nm, which is in accordance with the appearance of the aromatic ring in compound
4. The
13C-NMR data of this compound showed a carbonyl carbon signal at the δ
C value of 172.5 and a benzene ring with carbon signals at δ
C values of 159.8, 156.5, 132.7, 123.6, 122.3, and 104.6, as shown in
Table 1. In addition to the downfield carbon signals, the
1H-NMR and
13C-NMR data also showed an
N-substituted aliphatic methine (δ
H 6.04, δ
C 85.3), a methyl group (δ
H 2.13, δ
C 9.53), and two methoxy groups (δ
H 3.18, δ
C 52.2 and δ
H 3.96, δ
C 60.1, respectively). By a series of
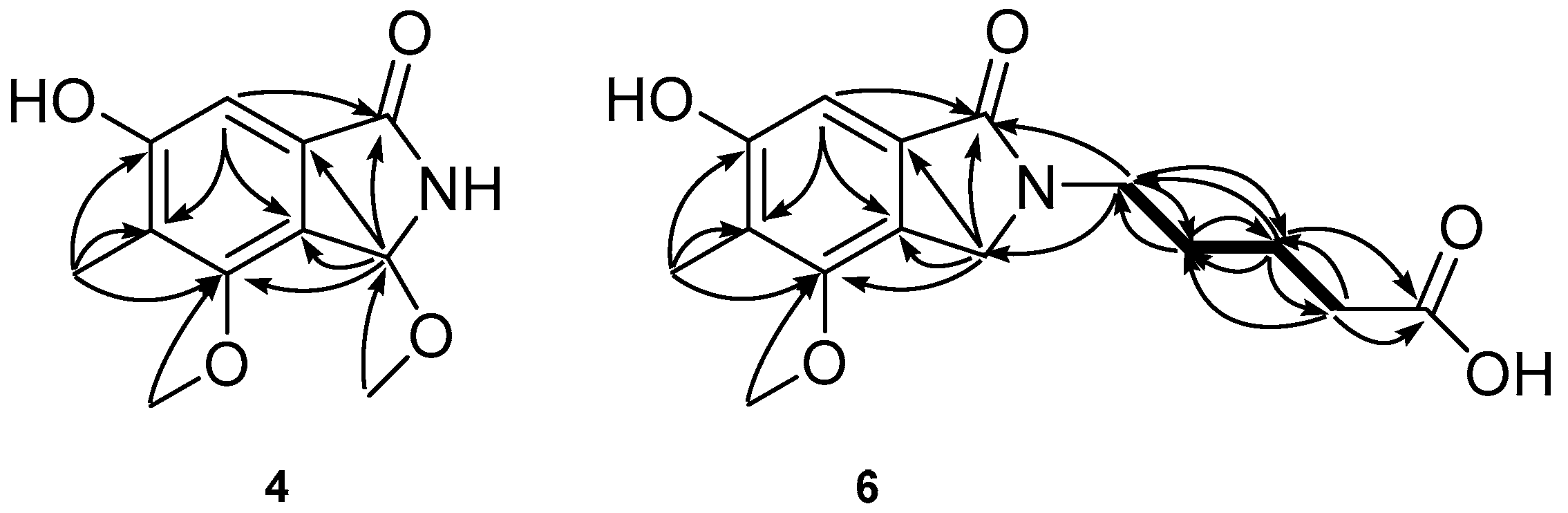
1H detected heteronuclear multiple bond for correlations (HMBC) between H-3 to C-1/C-5/C-6/C-7, H-8 to C-1/C-2/C-6/C-7, 5-CH
3 protons to C-4/C-5/C-6, 6-OCH
3 protons to C-6, and 8-OCH
3 protons to C-8 (
Figure 3), the planar structure of compound
4 was determined as 8-methoxycichorine, which was the derivative of cichorine (
3). The configuration of the asymmetric carbon center at C-8 is discussed below.
The molecular formula of 8-
epi-methoxycichorine (
5) was deduced to be C
11H
13NO
4 by HRESIMS analysis, which was identical to
4. The planar structure of this compound was determined to be the same as
4 based on the
1H-NMR and
13C-NMR results (
Table 1). Although the comparison of
1H-NMR and
13C-NMR chemical shifts between compounds
4 and
5 had no obvious difference, the optical rotations of both compounds were the opposite (the values for
4 and
5 were +5.3 and −4.1, respectively), suggesting these compounds exhibited an epimeric relationship at C-8.
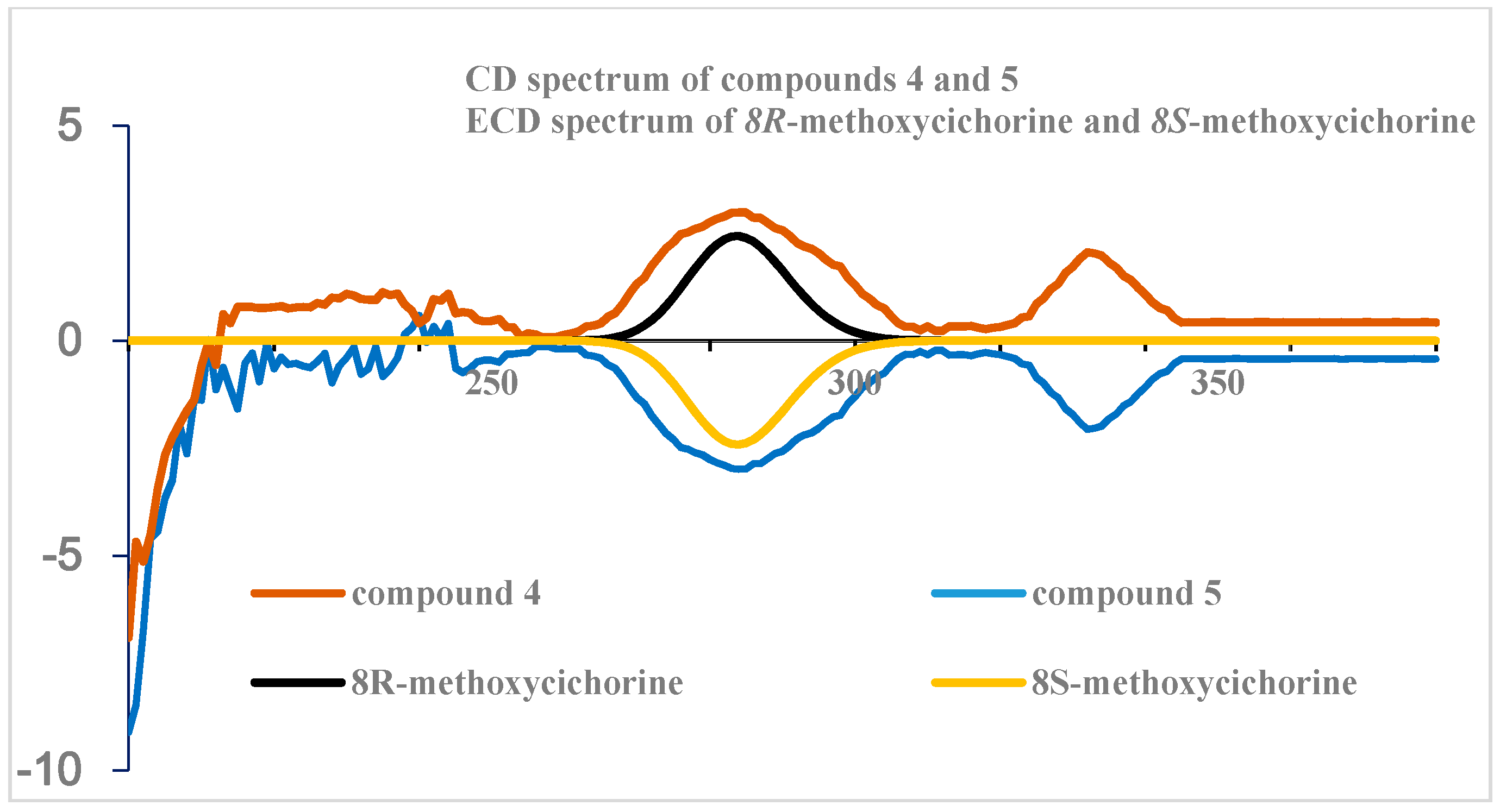
The absolute configuration at C-8 was eventually defined by circular dichroism (CD) measurements and ECD calculations. The CD profiles of
4 and
5 had opposite profiles at approximately 284 nm that were highly consistent with the ECD calculation for the model of 8
R and 8
S, respectively, as shown in
Figure 4. Additionally, the ECD spectrum for compound
4 is in accordance with the 3-methoxyporriolide [
19].
The molecular formula of
N-(4’-carboxybutyl) cichorine (
6) was deduced to be C
15H
19NO
5 by the HRESIMS analysis. The NMR data of this compound was very similar to that of cichorine (
3), with the most noticeable difference being the replacement of the carboxybutyl group at NH, which had signals at δ
C values of 43.2, 35.3, 28.9, 23.6, and 177.3, and δ
H values of 3.61, 2.33, 1.73, and 1.62. This interpretation was readily confirmed by a combination of 2D NMR experiments, in which all of the proton–proton and carbon–proton correlations obtained and supported the placement of a carboxybutyl group at the N position (
Figure 3). The carboxylic signal 177.3 in
13C-NMR was assigned to C-5’ based on the HMBC correlations between H-3’ (δ
H 1.62) to C-5’ (δ
C 177.3) and H-4’ (δ
H 2.33) to C-5’ (δ
C 177.3). Thus,
N-(4’-carboxybutyl)cichorine (
6) was structurally elucidated to be an
N-carboxybutyl derivative of
3.
The plausible biogenetic pathway for cichorine (
3) proposed by the Kawai Ken-ichi group is as follows [
18]. The first step is the formation of 3-methylorsellinate through the acetate-malonate pathway, followed by cyclization and the introduction of the C
1 unit at the C-5 position, followed by
O-methylation at C-6. The oxidation of the methyl group at C-1 of 3-methylorsellinate and cyclization makes compound
3. Compound
3 is the key intermediate, and transforms into
4,
5, and
6 by
O-methylation at C-8 and the introduction of a carboxybutyl group at the N position in this study, respectively.
The phytotoxic activities of zinniol-related compounds have been reported, and the relationship between their structure and phytotoxicity have been studied. However, the relationship between the structure and phytotoxicity of cichorine-derived compounds with an isoindolinone skeleton has rarely been discussed. Previous studies showed that the alkylation of the 6-OH group of cichorine (
3) abolished the phytotoxicity in a leaf-spot assay [
20]. We therefore investigated the activity of isolated compounds by a leaf-spot assay. Compounds
1 and
2 were inactive. The activities of
3–
6 are presented in
Table 2. Compounds
4 and
5, which have the methoxy group at C-8, exhibited more potential activity than cichorine (
3), indicating that structural modification of position C-8 in cichorine (
3) is important for improving the phytotoxicity of this type of compound. Compounds
4 and
5 exhibited similar activities toward the tested plant. Interestingly, compound
6 showed stronger phytotoxicity than cichorine (
3), as seen in
Table 2, suggesting that the
N-alkyl and carboxylic acid groups contributed to the phytotoxicity. However, it should be kept in mind that the relationship between the structure and phytotoxicity of cichorine-derived compounds depends on the species and organisms of the plants. For example, in a seedling-growth assay against stone leek and lettuce,
N-alkyl and hydroxyl groups enhanced the activity, but sulfonation almost abolished the activity when the 6-OH group of cichorine (
3) was alkylated. These results are important for the further modification of cichorine to discover the potential phytotoxins as herbicide candidates.
3. Materials and Methods
3.1. General Experimental Procedures
Optical rotations were measured on a JASCO P-1020 polarimeter (JASCO, Tokyo, Japan) using a 1 cm cell. UV spectra were acquired with a Hitachi U-3010 spectrophotometer (Hitachi, Tokyo, Japan). NMR spectra were recorded on a JEOL 400 spectrometer (JEOL, Tokyo, Japan). Proton and carbon NMR spectra were measured in MeOH-d4 and CDCl3 solutions at 400 and 100 MHz, respectively. High-resolution electrospray ionization mass spectrometry (HRESIMS) data were acquired using the Agilent 1290 Infinity LC System (Agilent, Palo Alto, USA) coupled with the Agilent 6540 Ultra High Definition Accurate-Mass Q-TOF LC/MS (Agilent, Palo Alto, USA). HPLC measurements were performed on an Essentia Pre LC-16P (Essentia, Kyoto, Japan) equipped with a UV detector. All solvents used were of spectroscopic grade or distilled from glass prior to use.
3.2. Molecular Genetic Methods
The gene deletion was performed using established gene targeting procedures [
21]. The 1267 bp upstream fragment and 928 bp downstream of the PKS
pkbA (AN6448) involved in cichorine biosynthesis were amplified from
A. nidulans LO8030 genomic DNA using PCR. Primers used in this study are listed in
Table S1. The pyrG gene was amplified, as a selectable marker cassette from
A. fumigatus 293 genomic DNA. The two amplified flanking sequences and the pyrG gene were fused together by PCR using nested primers. Production of protoplasts and transformations were carried out as previously described. The strain LO8030 carrying four mutations (i.e., pyroA4, riboB2, pyrG89, and nkuA::argB) was used as the recipient strain. For selection of transformants, 0.5 µg/mL pyridoxine and 2.5 µg/mL riboflavin were added to selection plates. For further verification, diagnostic PCR was performed two more times, as described using the primers located inside the targeted gene [
16], as shown in
Table S1.
3.3. Fermentation and Isolation
The A. nidulans strain LO8030 and ΔpkbA were cultured on YAG agar plates (5 g yeast extract, 20.0 g d-glucose, 15.0 g agar in 1 L distilled water, and 1 mL Hutner’s trace element solution) which were supplemented with riboflavin (2.5 mg/L), pyridoxin (0.5 mg/L), uracil (1 g/L), and uridine (10 mM). The fermentation was conducted at 37 °C for 7 days. Then, the plates were extracted with MeOH three times. The solvent was evaporated to obtain an organic extract. The extract was separated by C18 reversed-phase vacuum flash chromatography using a sequential mixture of MeOH and H2O as eluents (eight fractions in gradient, H2O‒MeOH, from 100:0 to 0:100), acetone, and finally EtOAc. On the basis of the results of the 1H-NMR analysis, the fraction eluted with H2O‒MeOH (60:40) and H2O‒MeOH (40:60) were chosen for separation. The fraction eluted with H2O‒MeOH (60:40) (3.32 g) was separated by semipreparative reversed-phase HPLC (from 70:30 to 40:60 in 40 min with H2O‒MeOH as the solvent system, 2.0 mL/min) to afford compounds 1 (2.0 mg), 2 (2.6 mg), and 3 (5.9 mg), respectively. The fraction eluted with H2O‒MeOH (40:60) (0.46 g) was separated by LH-20 (H2O‒MeOH, 40:60, 3 drops/min) followed with purification by reversed-phase HPLC analysis with H2O-MeCN as eluent to afford a mixture of 4 and 5 (3.6 mg), and compound 6 (0.9 mg). Compounds 4 (1.1 mg) and 5 (0.9 mg) were separated by a chiral column (Phenomenex lux cellulose-3, H2O–ACN as solvent system, from 90:10–0:100 in 30 min, 1.0 mL/min).
3,6-dimethyl-4-hydroxy-2-methoxybenzaldehyde (1): pale yellow amorphous solid; UV (EtOH) λmax (log ε): 232 (4.10), 284 (4.15) nm; 1H-NMR (400 MHz, CDCl3) δ: 10.35 (1H, s, 1-CHO), 6.97 (1H, br s, 4-OH), 6.52 (1H, s, 5-H), 3.83 (3H, s, 2-OMe), 2.54 (3H, s, 6-Me), 2.18 (3H, s, 3-Me). Compound 1 was identified by comparison of the 1H-NMR with the authentic sample.
nidulol (2): pale yellow amorphous solid; UV (EtOH) λmax (log ε): 215 (4.55), 256 (3.80), 303 (3.47) nm; HRESIMS, m/z 195.0654 [M + H]+ (Calcd. for C10H11O4, 195.0657). Compound 2 was identified by comparison of the 1H-, 13C-NMR chemical shifts and HRESIMS with the authentic sample.
cichorine (3): pale yellow amorphous solid; UV (MeOH) λmax (log ε): 212 (4.00), 252 (3.32), 295 (3.01) nm; HRESIMS, m/z 194.0815 [M + H]+ (Calcd. for C10H12NO3, 194.0817). Compound 3 was identified by comparison of the 1H-, 13C-NMR chemical shifts and HRESIMS with the authentic sample.
8-methoxycichorine (
4): yellow amorphous solid; m.p. 287 °C;
+ 5.3 (
c 0.5, MeOH); UV (MeOH)
λmax (log ε) 215 (4.07), 259 (3.37), 298 (3.10) nm; CD (MeOH) λ (∆ε) 284 (+3.05), 352 (+2.06) nm; IR (ZnSe) ν
max 3394 (br), 1665, 1572 cm
−1;
1H- and
13C-NMR data, see
Table 1; HRESIMS,
m/
z 246.0751 [M + Na]
+ (Calcd. for C
11H
13NO
4Na, 246.0742).
8-epi-methoxycichorine (
5): yellow amorphous solid; m.p. 287 °C;
− 4.1 (
c 0.5, MeOH); UV (MeOH)
λmax (log ε) 215 (4.07), 259 (3.37), 298 (3.10) nm; CD (MeOH) λ (∆ε) 284 (−3.02), 352 (−2.08) nm; IR (ZnSe) ν
max 3393 (br), 1666, 1579 cm
−1;
1H- and
13C-NMR data, see
Table 1; HRESIMS,
m/
z 246.0751 [M + Na]
+ (Calcd. for C
11H
13NO
4Na, 246.0742).
N-(4’-carboxybutyl)cichorine (
6): white amorphous solid; m.p. 470 °C; UV (MeOH)
λmax (log ε) 213 (4.03), 254 (3.33), 295 (3.05) nm; IR (ZnSe) ν
max 3374, 1778, 1662, 1572 cm
−1;
1H- and
13C-NMR data, see
Table 1; HRESIMS,
m/
z 294.1348 [M + H]
+ (Calcd. for C
15H
20 NO
5, 294.1341).
3.4. Computational Analysis
The ground-state geometries were optimized with density functional theory (DFT) calculations using Turbomole 6.5 with the basis set def-SV(P) for all atoms at the DFT level, using the B3LYP functional. The ground states were further confirmed by a harmonic frequency calculation. The calculated ECD data corresponding to the optimized structures were obtained using time-dependent density-functional theory (TDDFT) with the basis set def2-TZVPP for all atoms at the DFT level, using the B3LYP functional. The ECD spectra were simulated by overlapping each traction, where
σ is the width of the band at 1/e height. t
Ei and
Ri are the excitation energies and rotatory strengths for transition
i, respectively. In the current work, the value of σ was fixed at 0.09 eV.
3.5. Phytotoxic Bioassay
The phytotoxic activities of the cichorine analogues were tested on the cut leaves using a moist chamber technique as described in reference [
15]. The leaves were from
Zea mays and
Medicago polymorpha L., respectively. All the used leaves were cut from three or four-week-old plants with the size 1.5 cm × 1.5 cm. All the tested compounds were prepared with the concentration at 5 × 10
−3 M. The pricks were made using the tip of a 10 µL Hamilton syringe, and a droplet (10 µL) of test solution was applied in the center of the leaves from the upper side. The diameters of the lesions were measured after 48 h of incubation in a moist, sterile chamber at 25 °C. The compounds were prepared in 2% EtOH. The cichorine was chosen as the positive control and 2% EtOH was tested as the negative control.
3.6. Statistical Analysis
Experiments were carried out at least in triplicate, and results were expressed as the mean ± standard deviation (SD). Statistical analysis was performed using the SPSS statistical package for the Social Sciences software. A p value of < 0.05 was considered to be statistically significant.







{kind=link}
{kind=link}
{kind=link}
{kind=link}