Effects of Puerarin on the Ovariectomy-Induced Depressive-Like Behavior in ICR Mice and Its Possible Mechanism of Action

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
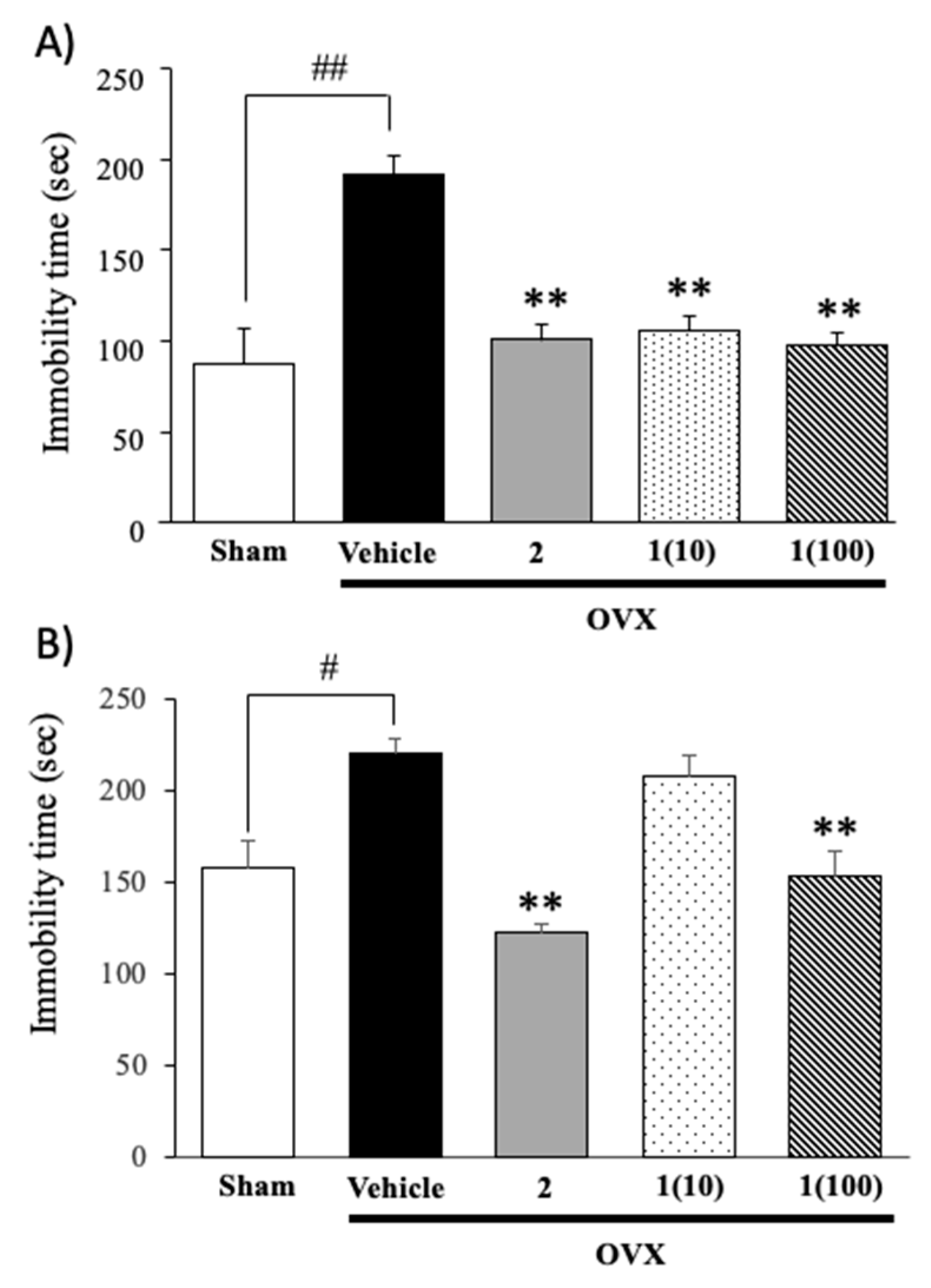
2.1. Effects of Puerarin (1) and 17β-Estradiol (2) on Ovariectomy-Induced Depressive-Like Behavior
2.2. Changes in Uterine Weights and Volumes after the Administration of Puerarin (1) to OVX Animals
2.3. Administration of Puerarin (1) Reverses the Ovariectomy-Induced Increase in Serum Corticosterone Levels
2.4. Effects of Puerarin (1) and 17β-Estradiol (2) on Ovariectomy-Induced Changes in the Hippocampal Expression of Genes Encoding BDNF, ERβ, and ERα
2.5. Puerarin (1) Inhibited the Ovariectomy-Induced Suppression of Neurogenesis in the Dentate Gyrus Area in the Hippocampus
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Materials and Isolation of Puerarin (1)
4.3. Animals
4.4. Surgical Procedure
4.5. Drug Administration
4.6. Behavioral Studies
4.6.1. Locomotor Test
4.6.2. Forced Swimming Test (FST)
4.6.3. Tail Suspension Test (TST)
4.7. Uterine Hypertrophy Test
4.8. Neurochemical Studies
4.8.1. Measurement of Serum Corticosterone Levels
4.8.2. Quantitative Real-Time Polymerase Chain Reaction (QPCR)
4.9. Analysis of Neurogenesis in the Dentate Gyrus
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bastos, C.P.; Pereira, L.M.; Ferreira-Vieira, T.H.; Drumond, L.E.; Massensini, A.R.; Moraes, M.F.; Pereira, G.S. Object recognition memory deficit and depressive-like behavior caused by chronic ovariectomy can be transitorialy recovered by the acute activation of hippocampal estrogen receptors. Psychoneuroendocrinology 2015, 57, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.T.; Mapstone, M.; Staskiewicz, J.; Maki, P.M. Reconciling subjective memory complaints with objective memory performance in the menopausal transition. Menopause 2012, 197, 735–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudielka, B.M.; Schmidt-Reinwald, A.K.; Hellhammer, D.H.; Kirschbaum, C. Psychological and endocrine responses to psychosocial stress and dexamethasone/corticotropin-releasing hormone in healthy postmenopausal women and young controls: The impact of age and a two-week estradiol treatment. Neuroendocrinology 1999, 70, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Patacchioli, F.R.; Simeoni, S.; Monnazzi, P.; Pace, M.; Capri, O.; Perrone, G. Menopause, mild psychological stress and salivary cortisol: Influence of long-term hormone replacement therapy (HRT). Maturitas 2006, 55, 150–155. [Google Scholar] [CrossRef]
- Farquhar, C.; Marjoribanks, J.; Lethaby, A.; Suckling, J.A.; Lamberts, Q. Long term hormone therapy for perimenopausal and postmenopausal women. Cochrane Database Syst. Rev. 2009, 15, CD004143. [Google Scholar] [CrossRef]
- Narod, S.A. Hormone replacement therapy and the risk of breast cancer. Nat. Rev. Clin. Oncol. 2001, 8, 669–676. [Google Scholar] [CrossRef]
- Malaivijitnond, S. Medical applications of phytoestrogens from the Thai herb Pueraria mirifica. Front. Med. 2012, 6, 8–21. [Google Scholar] [CrossRef]
- Peerakam, N.; Sirisa-ard, P.; Quoc Huy, N.; Van On, T.; Tuan Long, P.; Intharuksa, A. Isoflavonoids and phytoestrogens from Pueraria candollei var. mirifica related with appropriate ratios of ethanol extraction. Asian J. Chem. 2017, 30, 2086–2090. [Google Scholar] [CrossRef]
- Suthon, S.; Jaroenporn, S.; Charoenphandhu, N.; Suntornsaratoon, P.; Malaivijitnond, S. Anti-osteoporotic effects of Pueraria candollei var. mirifica on bone mineral density and histomorphometry in estrogen-deficient rats. J. Nat. Med. 2016, 70, 225–233. [Google Scholar] [CrossRef]
- Zhou, Y.X.; Zhang, H.; Peng, C. Puerarin: A review of pharmacological effects. Phytother. Res. 2014, 28, 961–975. [Google Scholar] [CrossRef]
- Wei, S.Y.; Chen, Y.; Xu, X.Y. Progress on the pharmacological research of puerarin: A review. Chin. J. Nat. Med. 2014, 12, 407–414. [Google Scholar] [CrossRef]
- Shi, W.; Yuan, R.; Chen, X.; Xin, Q.; Wang, Y.; Shang, X.; Cong, W.; Chen, K. Puerarin reduces blood pressure in spontaneously hypertensive rats by targeting eNOS. Am. J. Chin. Med. 2019, 47, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Chang, S.; Xiao, Z.; Qi, Y. The protective effect of puerarin on angiotensin II-induced aortic aneurysm formation by the inhibition of NADPH oxidase activation and oxidative stress-triggered AP-1 signaling pathways. Oncol. Lett. 2018, 16, 3327–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.-K.; Zhang, G.-H.; Zhong, D.-S.; He, J.-L.; Liu, X.; Chen, J.-S.; Wei, D.-N. Puerarin ameliorated the behavioral deficits induced by chronic stress in rats. Sci. Rep. 2017, 7, 6266. [Google Scholar] [CrossRef] [Green Version]
- Walf, A.A.; Frye, C.A. A review and update of mechanisms of estrogen in the hippocampus and amygdala for anxiety and depression behavior. Neuropsychopharmacology 2006, 31, 1097–1111. [Google Scholar] [CrossRef]
- Borrow, A.P.; Cameron, N.M. Estrogenic mediation of serotonergic and neurotrophic systems: Implications for female mood disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 54, 13–25. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Chatterjee, M.; Jaiswal, M.; Palit, G. Comparative evaluation of forced swim test and tail suspension test as models of negative symptom of schizophrenia in rodents. ISRN Psychiatry 2012, 595141. [Google Scholar] [CrossRef] [Green Version]
- Rachon, D.; Vortherms, T.; Seidlova-Wuttke, D.; Wuttke, W. Dietary daidzein and puerarin do not affect pituitary LH expression but exert uterotropic effects in ovariectomized rats. Maturitas 2007, 57, 161–170. [Google Scholar] [CrossRef]
- Loutchanwoot, P.; Vortherms, T.; Jarry, H. Evaluation of in vivo estrogenic potency of natural estrogen-active chemical, puerarin, on pituitary function in gonadectomized female rats. Life Sci. 2016, 165, 75–82. [Google Scholar] [CrossRef]
- Boue, S.M.; Wiese, T.E.; Nehls, S.; Burow, M.E.; Elliott, S.; Carter-Wientjes, C.H.; Shih, B.Y.; Mclachlan, J.A.; Cleveland, T.E. Evaluation of the estrogenic effects of legume extracts containing phytoestrogens. J. Agric. Food Chem. 2003, 51, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, O.O.; Zhou, J.; Samathanam, C.; Powell-Braxton, L.; Bondy, C.A. Insulin-like growth factor 1 is required for G2 progression in the estradiol-induced mitotic cycle. Proc. Natl. Acad. Sci. USA 1999, 96, 3287–3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balog, M.; Miljanovic, M.; Blazetic, S.; Labak, I.; Ivic, V.; Viljetic, B.; Borbely, A.; Papp, Z.; Blazekovic, R.; Vari, S.G.; et al. Sex-specific chronic stress response at the level of adrenal gland modified sexual hormone and leptin receptors. Croat. Med. J. 2015, 56, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fester, L.; Prange-Kiel, J.; Jarry, H.; Rune, G.M. Estrogen synthesis in the hippocampus. Cell Tissue Res. 2011, 345, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Baudry, M.; Bi, X.; Aguirre, C. Progesterone-estrogen interactions in synaptic plasticity and neuroprotection. Neuroscience 2013, 239, 280–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zárate, S.; Stevnsner, T.; Gredilla, R. Role of Estrogen and Other Sex Hormones in Brain Aging. Neuroprotection and DNA Repair. Front. Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bean, L.A.; Ianov, L.; Foster, T.C. Estrogen receptors, the hippocampus, and memory. Neuroscientist 2014, 20, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Voudouri, K.; Berdiaki, A.; Tzardi, M.; Tzanakakis, G.N.; Nikitovic, D. Insulin-Like Growth Factor and Epidermal Growth Factor Signaling in Breast Cancer Cell Growth: Focus on Endocrine Resistant Disease. Anal. Cell. Pathol. (Amst.) 2015, 2015, 975495. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.L. Cross-talk between peptide growth factor and estrogen receptor signaling pathways. Biol. Reprod. 1998, 58, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Mizuki, D.; Matsumoto, K.; Tanaka, K.; Thi Le, X.; Fujiwara, H.; Ishikawa, T.; Higuchi, Y. Antidepressant-like effect of Butea superba in mice exposed to chronic mild stress and its possible mechanism of action. J. Ethnopharmacol. 2014, 156, 16–25. [Google Scholar] [CrossRef]
- Du, X.; Pang, T.Y. Is dysregulation of the HPA-axis a core pathophysiology mediating co-morbid depression in neurodegenerative diseases? Front. Psychiatry 2015, 6, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menke, D. Is the HPA Axis as Target for Depression Outdated, or Is There a New Hope? Front. Psychiatry 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.D.; Rovis, T.; Chung, W.C.; Handa, R.J. Novel action of estrogen receptor-beta on anxiety-related behaviors. Endrocrinology 2005, 146, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Oyala, M.G.; Portillo, W.; Reyna, A.; Foradori, C.D.; Kudwa, A.; Hinds, L.; Handa, R.J.; Mani, S.K. Anxiolytic effects and neuroanatomical targets of estrogen receptor-β (ERβ) activation by a selective ERβ agonist in female mice. Endocrinology 2012, 153, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhu, X.; Liu, S.; Nicholson, R.C.; Ni, X. Phytoestrogens induce differential estrogen receptor beta-mediated responses in transfected MG-63 cells. Endocrine 2008, 34, 29–35. [Google Scholar] [CrossRef]
- Ninan, I. Synaptic regulation of affective behaviors; role of BDNF. Neuropharmacology 2014, 76, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Numakawa, T.; Odaka, H.; Adachi, N. Actions of brain-derived neurotrophic factor and glucocorticoid stress in neurogenesis. Int. J. Mol. Sci. 2017, 18, 2312. [Google Scholar] [CrossRef] [Green Version]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A key factor with multipotent impact on brain signaling and synaptic plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Tolwani, R.J.; Buckmaster, P.S.; Varma, S.; Cosgaya, J.M.; Wu, Y.; Suri, C.; Shooter, E.M. BDNF overexpression increases dendrite complexity in hippocampal dentate gyrus. Neuroscience 2002, 114, 795–805. [Google Scholar] [CrossRef]
- Kang, H.; Schuman, E.M. Long-lasting neurotrophin-induced enhancement of synaptic transmission in the adult hippocampus. Science 1995, 267, 1658–1662. [Google Scholar] [CrossRef]
- Sohrabji, F.; Miranda, R.C.; Toran-Allerand, C.D. Identification of a putative estrogen response element in the gene encoding brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 1995, 92, 11110–11114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, A.; Delattre, A.M.; Pereira, S.I.; Carolino, R.G.; Szawka, R.E.; Anselmo-Franci, J.A.; Zanata, S.M.; Ferraz, A.C. 17β-estradiol replacement in young, adult and middle-aged female ovariectomized rats promotes improvement of spatial reference memory and an antidepressant effect and alters monoamines and BDNF levels in memory- and depression-related brain areas. Behav. Brain Res. 2012, 227, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, H.; Cohen, R.S.; Pandey, S.C. Effects of estrogen treatment on expression of brain-derived neurotrophic factor and cAMP response element-binding protein expression and phosphorylation in rat amygdaloid and hippocampal structures. Neuroendocrinology 2005, 81, 294–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gass, P.; Reichardt, H.M.; Strekalova, T.; Henn, F.; Tronche, F. Mice with targeted mutations of glucocorticoid and mineralocorticoid receptors: Models for depression and anxiety? Physiol. Behav. 2001, 73, 811–825. [Google Scholar] [CrossRef]
- Bechstedt, S.; Lu, K.; Brouhard, G.J. Doublecortin recognizes the longitudinal curvature of the microtubule end and lattice. Curr. Biol. 2014, 24, 2366–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, J.L.; Markham, K.R.; Dziedzic, S.Z.; Pope, G.S. Puerarin 6″-O-β-apiofuranoside, a C-glycosylisoflavone O-glycoside from Pueraria mirifica. Phytochemistry 1986, 25, 1772–1775. [Google Scholar] [CrossRef]
- Monthakantirat, O.; Sukano, W.; Umehara, K.; Noguchi, H.; Chulikhit, Y.; Matsumoto, K. Effect of miroestrol on ovariectomy-induced cognitive impairment and lipid peroxidation in mouse brain. Phytomedicine 2014, 21, 1249–1255. [Google Scholar] [CrossRef]
- Mizuki, D.; Qi, Z.; Tanaka, K.; Fujiwara, H.; Ishikawa, T.; Higuchi, Y.; Matsumoto, K. Butea superba-induced amelioration of cognitive and emotional deficits in olfactory bulbectomized mice and putative mechanisms underlying its actions. J. Pharmacol. Sci. 2014, 124, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Khamphukdee, C.; Monthakantirat, O.; Chulikhit, Y.; Buttachon, S.; Lee, M.; Silva, A.M.S.; Sekeroglu, N.; Kijjoa, A. Chemical constituents and antidepressant-like effects in ovariectomized mice of the ethanol extract of Alternanthera Philoxeroides. Molecules 2018, 23, 2202. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: The sample of puerarin is not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Uterine Weight/Body Weight (g/kg) | Uterine Volume/Body Volume (cm3/kg) |
|---|---|---|
| Sham-operated | 7.68 ± 0.31 | 10.32 ± 1.39 |
| OVX + vehicle | 1.66 ± 0.11 ## | 1.37 ± 0.31 ## |
| OVX + 2 | 5.50 ± 0.50 ** | 4.79 ± 0.23 * |
| OVX + 1 (10 mg/kg) | 1.57 ± 0.15 ‡ | 2.35 ± 0.34 |
| OVX + 1 (100 mg/kg) | 3.15 ± 0.48 ** | 4.94 ± 0.81 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tantipongpiradet, A.; Monthakantirat, O.; Vipatpakpaiboon, O.; Khampukdee, C.; Umehara, K.; Noguchi, H.; Fujiwara, H.; Matsumoto, K.; Sekeroglu, N.; Kijjoa, A.; et al. Effects of Puerarin on the Ovariectomy-Induced Depressive-Like Behavior in ICR Mice and Its Possible Mechanism of Action. Molecules 2019, 24, 4569. https://doi.org/10.3390/molecules24244569
Tantipongpiradet A, Monthakantirat O, Vipatpakpaiboon O, Khampukdee C, Umehara K, Noguchi H, Fujiwara H, Matsumoto K, Sekeroglu N, Kijjoa A, et al. Effects of Puerarin on the Ovariectomy-Induced Depressive-Like Behavior in ICR Mice and Its Possible Mechanism of Action. Molecules. 2019; 24(24):4569. https://doi.org/10.3390/molecules24244569
Chicago/Turabian StyleTantipongpiradet, Ariyawan, Orawan Monthakantirat, Onchuma Vipatpakpaiboon, Charinya Khampukdee, Kaoru Umehara, Hiroshi Noguchi, Hironori Fujiwara, Kinzo Matsumoto, Nazim Sekeroglu, Anake Kijjoa, and et al. 2019. "Effects of Puerarin on the Ovariectomy-Induced Depressive-Like Behavior in ICR Mice and Its Possible Mechanism of Action" Molecules 24, no. 24: 4569. https://doi.org/10.3390/molecules24244569