
Accession-Level Differentiation of Urushiol Levels, and Identification of Cardanols in Nascent Emerged Poison Ivy Seedlings


Abstract
:1. Introduction
2. Results
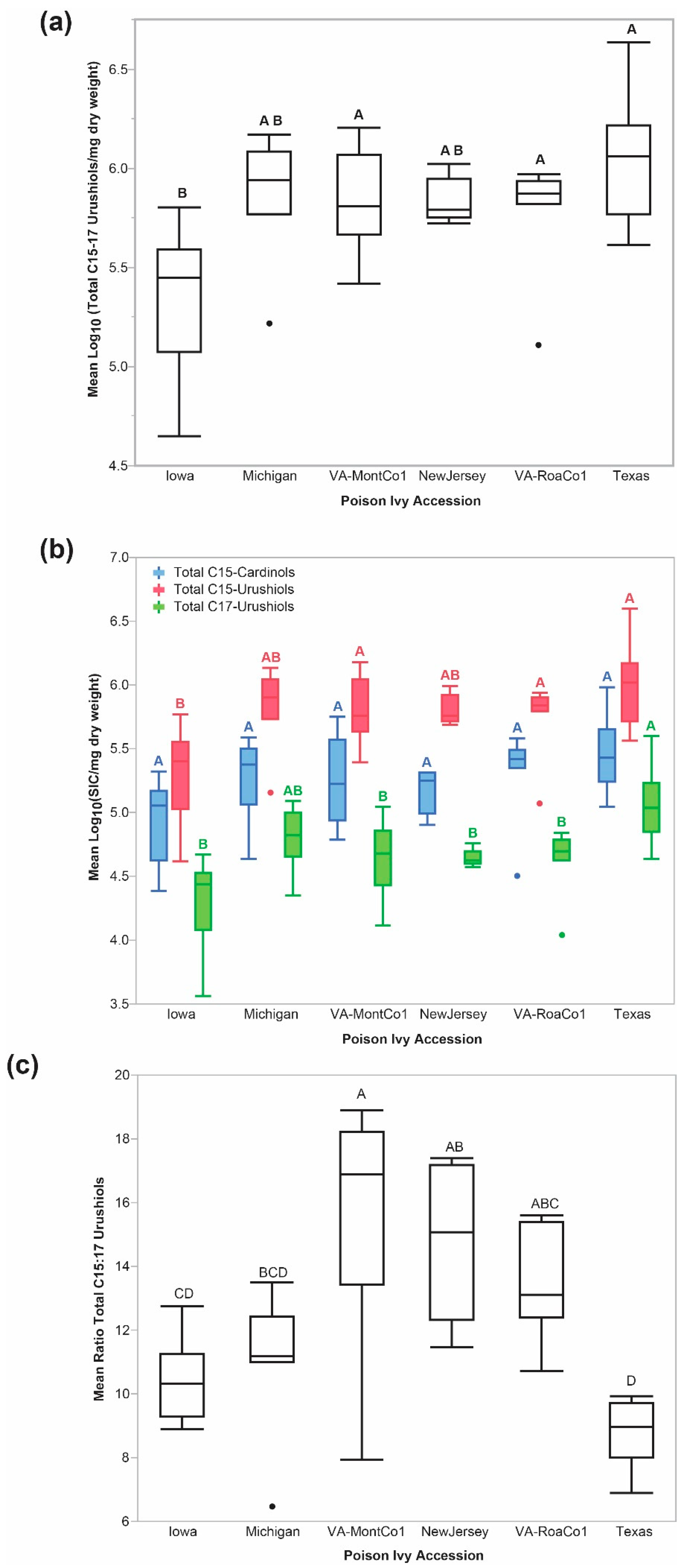
2.1. Accession-Level Differences in Steady State Total C15- and Total C17-Urushiol Accumulation Levels
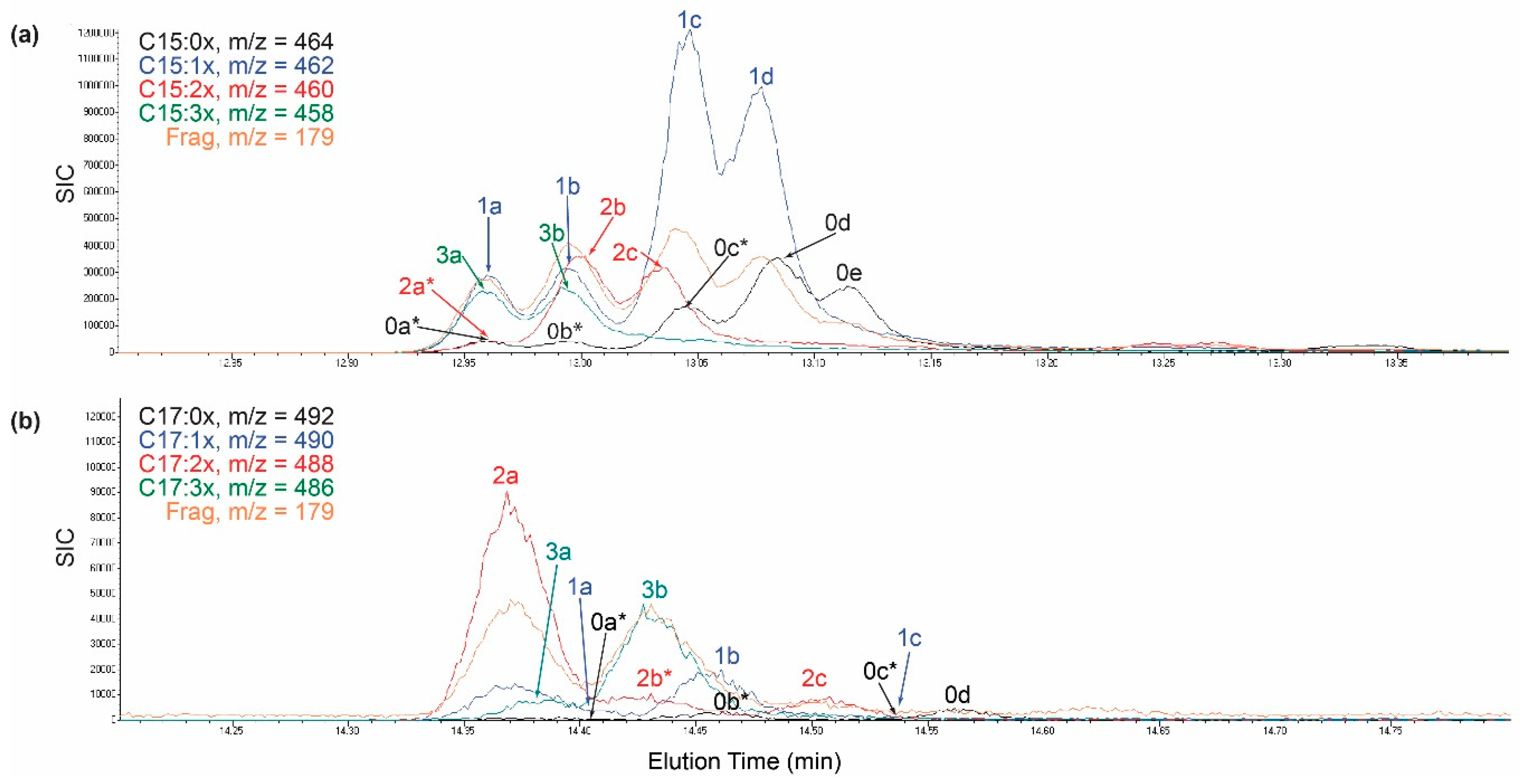
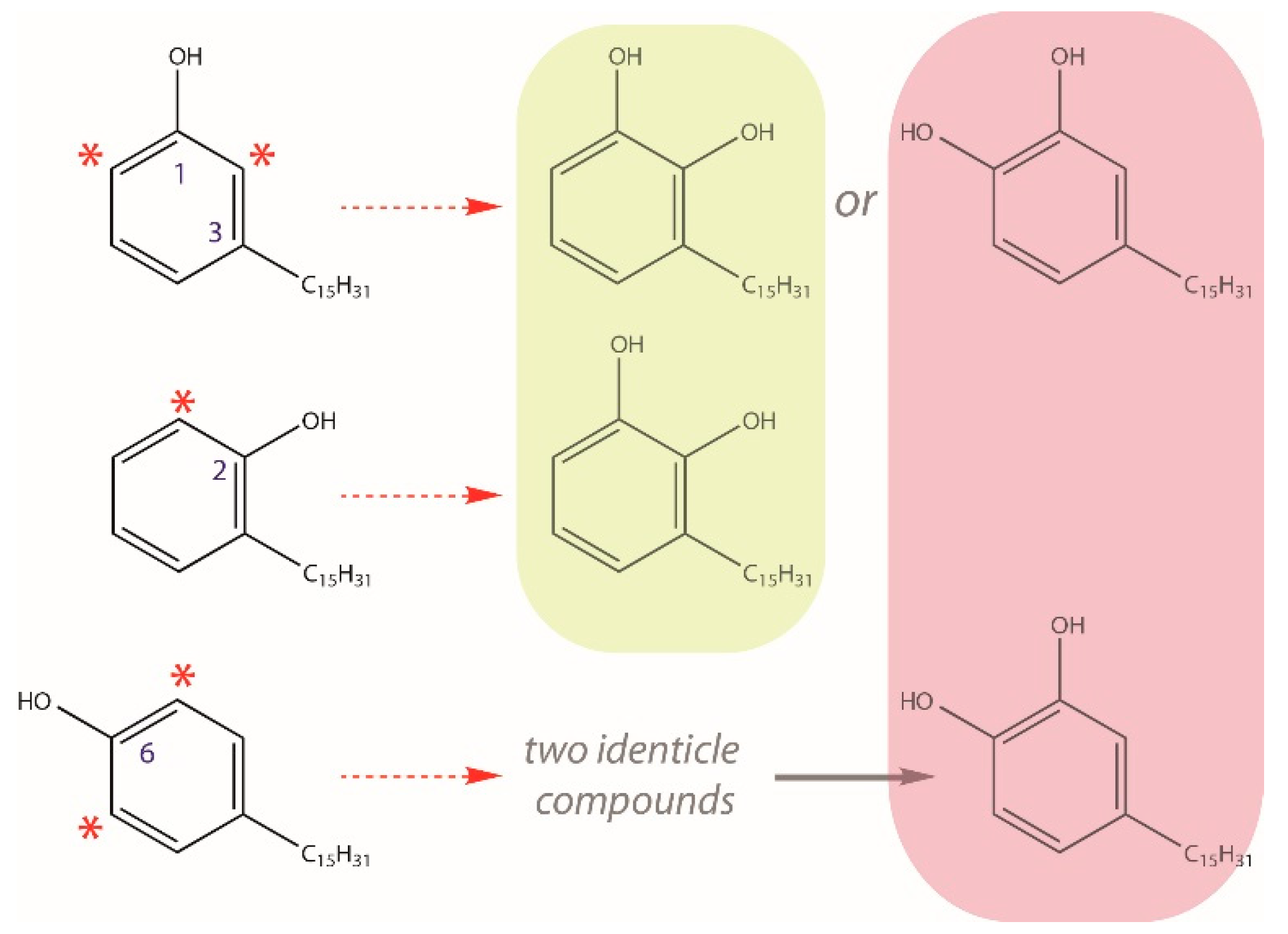
2.2. Identification of Novel C15- and C17-Urushiol Isomers
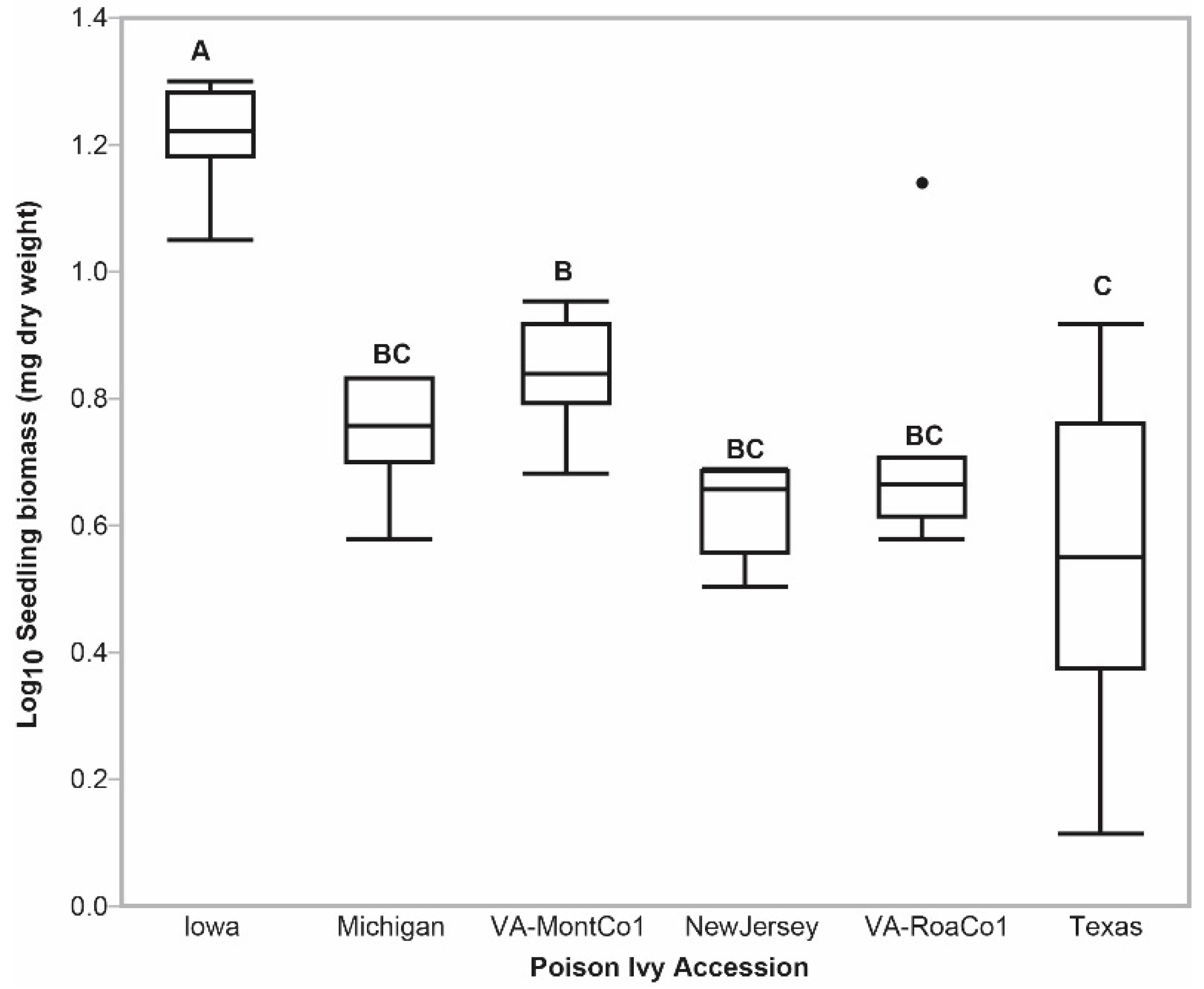
2.3. Urushiol Accumulation Imposed a Metabolic Load on Nascent Emerged Seedling Vigor
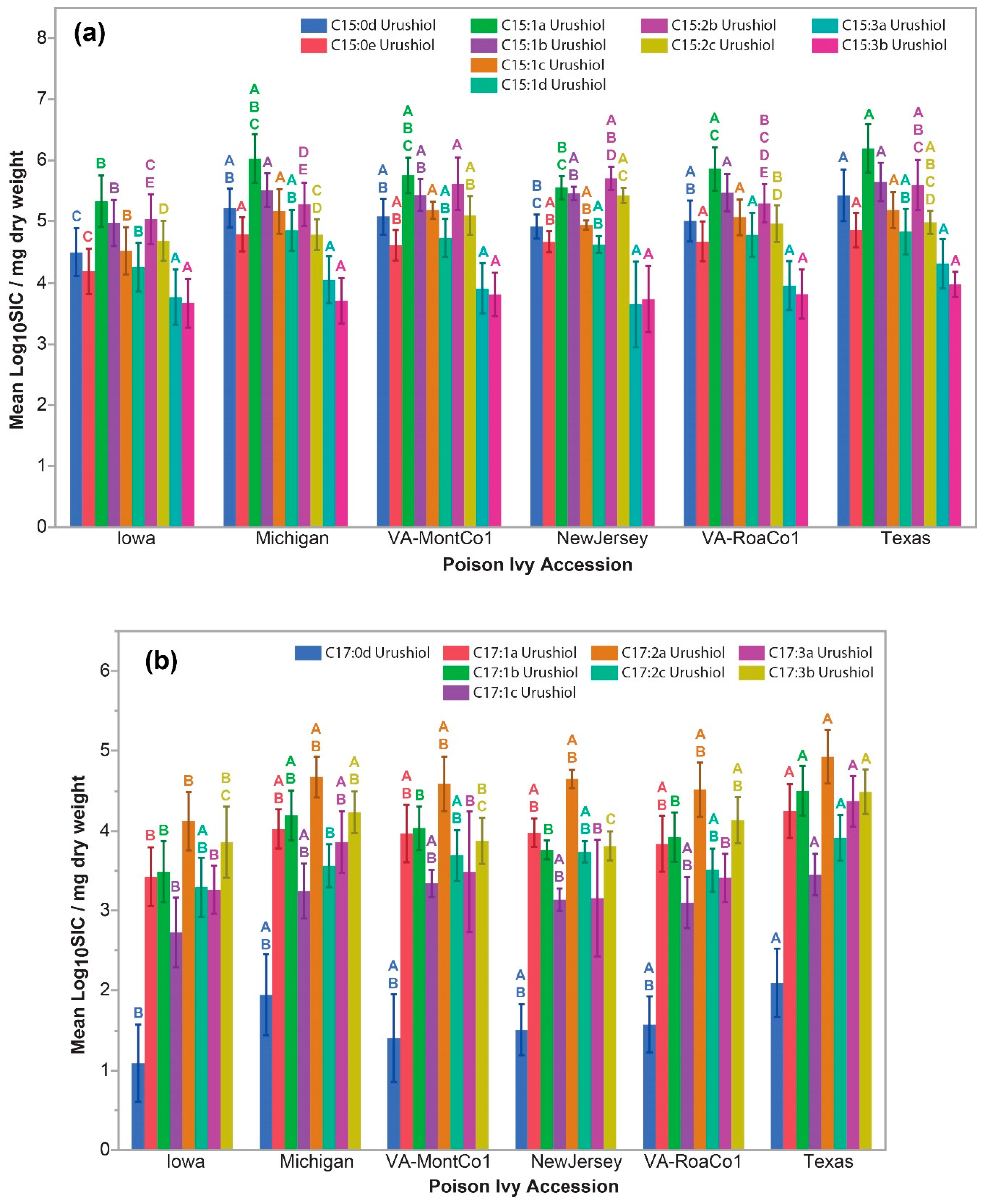
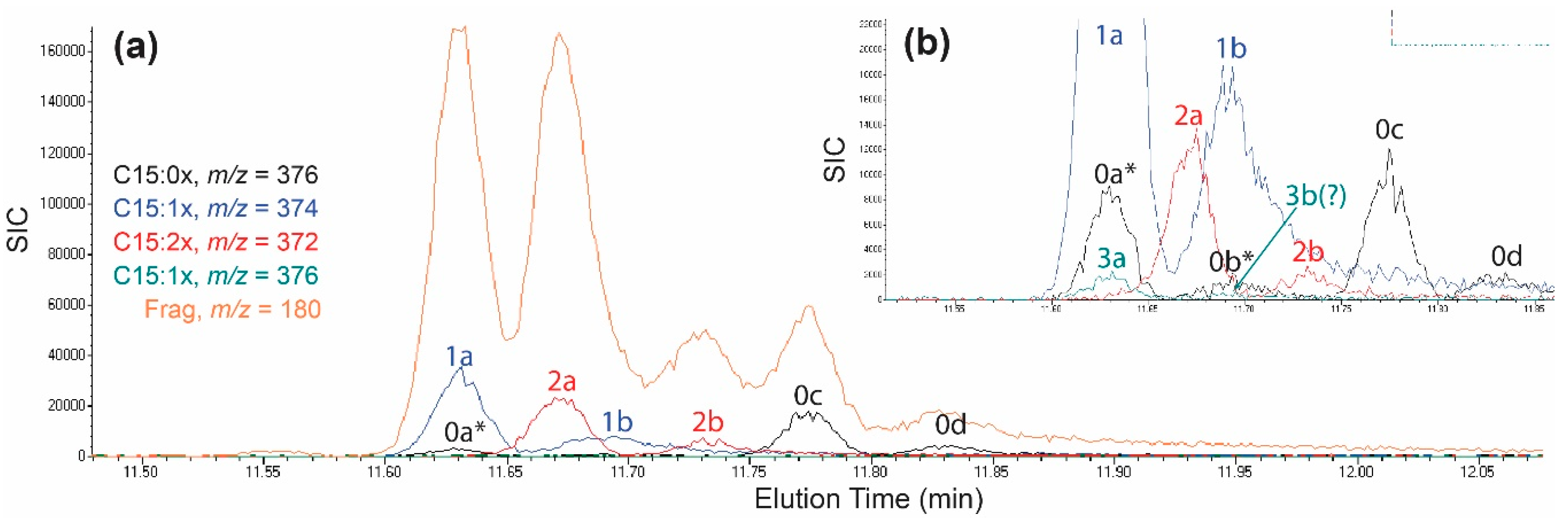
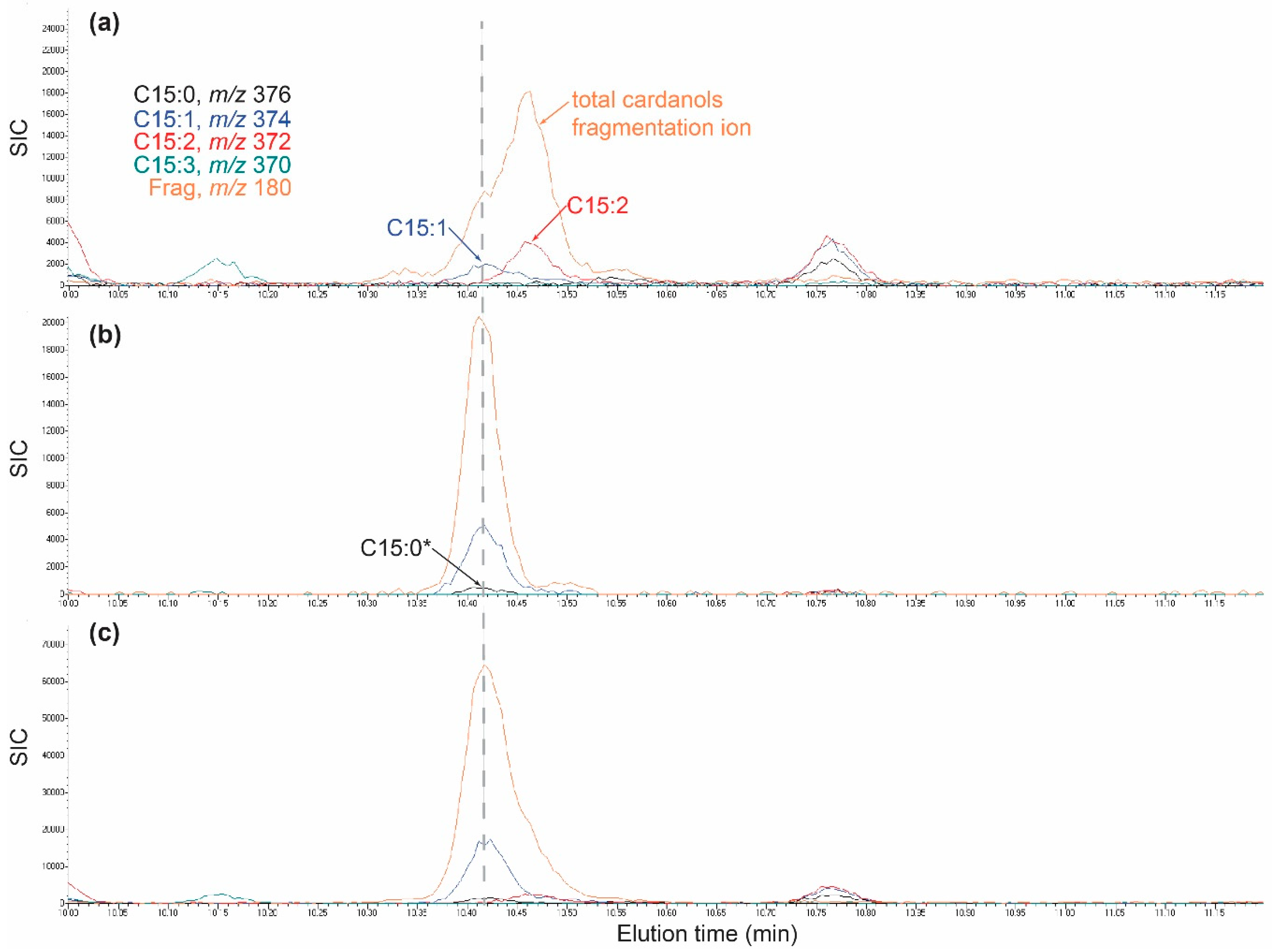
2.4. Identification of C15-Cardanol Congeners and Isomers in Poison Ivy Seedlings
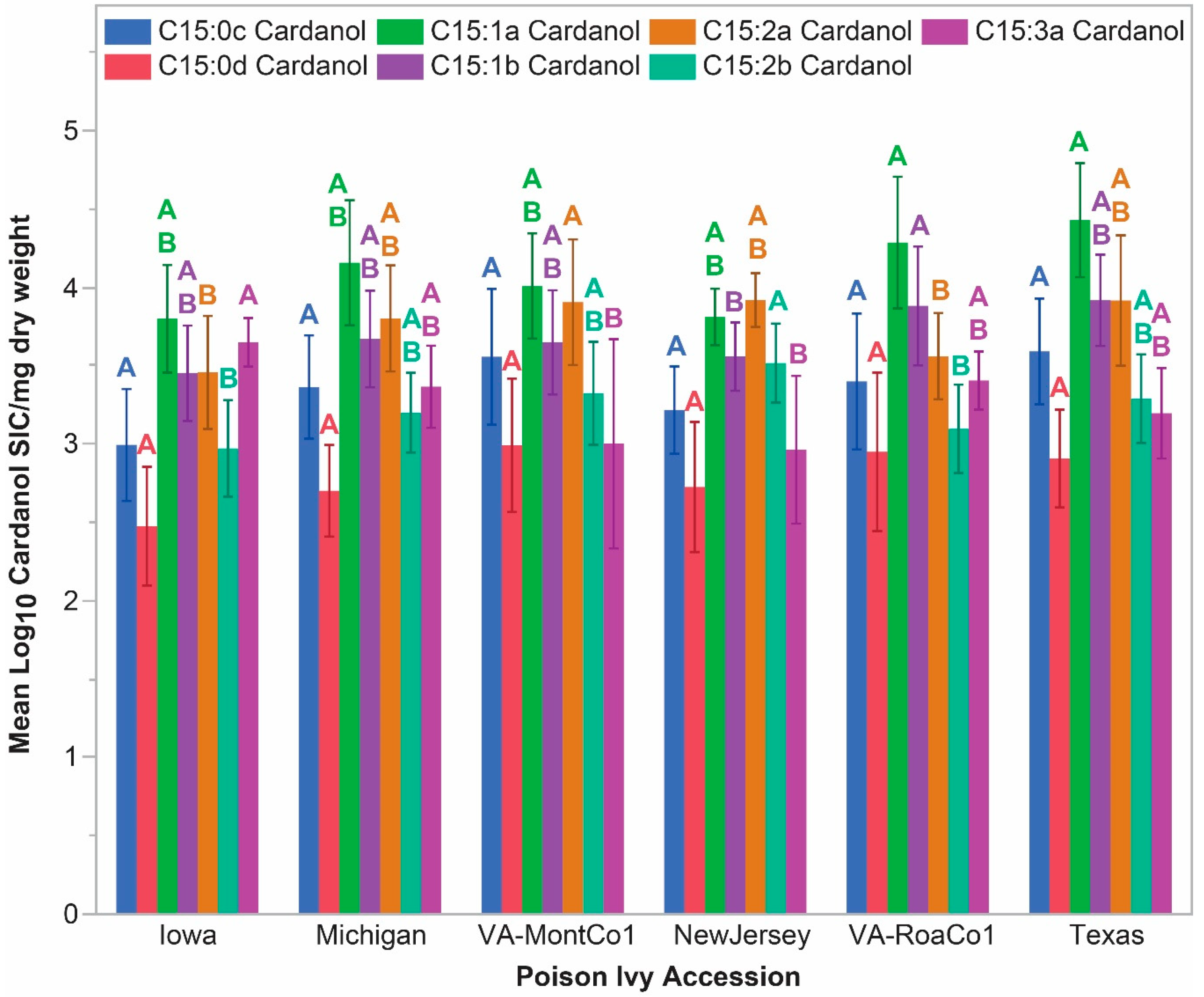
2.5. Significant Accession-Level C15-Cardanol Accumulation Levels
3. Discussion
4. Materials and Methods
4.1. Poison Ivy Seedling Germination and Harvest
4.2. Alkylphenol Extraction and Assay
4.3. Statistical Analysis of Urushiol and Cardanol Congener Steady State Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Barkley, F.A. A monographic study of Rhus and its immediate allies in North and Central America, including the West Indies. Annu. Missouri Bot. Gard. 1937, 37, 265–498. [Google Scholar] [CrossRef]
- Gillis, W.T. The systematics and ecology of poison-ivy and the poison-oaks (Toxicodendron, Anacardiaceae). Rhodora 1971, 73, 72–159, 161–237, 370–443, 465–540. [Google Scholar]
- Epstein, W.L. Plant-induced dermatitis. Ann. Emerg. Med. 1987, 16, 950–955. [Google Scholar] [CrossRef]
- Epstein, W.L. Occupational poison ivy and oak dermatitis. Dermatol. Clin. 1994, 3, 511–516. [Google Scholar] [CrossRef]
- Pariser, D.M.; Ceilley, R.I.; Lefkovits, A.M.; Katz, B.E.; Paller, A.S. Poison ivy, oak and sumac. Dermatol. Insights 2003, 4, 26–28. [Google Scholar]
- Hill, G.A.; Mattacotti, V.; Graha, W.D. The toxic principle of the poison ivy. J. Am. Chem. Soc. 1934, 56, 2736–2738. [Google Scholar] [CrossRef]
- Majima, R. Uber den hauptbestandteil des Japan-lacs, VIII. Mitteilung: Stellung der doppelbindungen in der seitenkette des urushiols und beweisfuhrung, dab das urushiol eine mischung ist. Ber. Dtsch. Chem. Ges. 1922, 55, 172–191. [Google Scholar] [CrossRef]
- Majima, R.; Cho, S. Ueber einen hauptbestandteil des japanischen lackes. Ber. Dtsch. Chem. Ges. 1907, 40, 632. [Google Scholar] [CrossRef]
- Symes, W.F.; Dawson, C.R. Separation and structural determination of the olefinic components of poison ivy urushiol, cardanol and cardol. Nature 1953, 171, 841–842. [Google Scholar] [CrossRef]
- Sunthankar, S.V.; Dawson, C.R. The structural identification of the olefinic components of Japanese Lac urushiol. J. Am. Chem. Soc. 1954, 76, 5070–5074. [Google Scholar] [CrossRef]
- Markiewitz, K.H.; Dawson, C.R. On the isolation of the allegenically active components of the toxic principle of poison ivy. J. Org. Chem. 1965, 30, 1610–1613. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Adawadkar, P.D.; Benigni, D.A.; Watson, E.S.; Little, T.L., Jr. Analogues of poison ivy urushiol. Synthesis and biological activity of disubstituted n-alkylbenzenes. J. Med. Chem. 1986, 29, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Baer, H.; Watkins, R.C.; Kurtz, A.P.; Byck, J.S.; Dawson, C.R. Delayed contact sensitivyt to catechols III. The relationship of side-chain length to sensitizing potency of catechols chemically related to the active principles of poison ivy. J. Immunol. 1967, 99, 370–375. [Google Scholar] [PubMed]
- Johnson, R.A.; Baer, H.; Kirkpatrick, C.H.; Dawson, C.R.; Khurana, R.G. Comparison of the contact allergenicity of the four pentadecylcatechols derived from poison ivy urushiol in human subjects. J. Allergy Clin. Immunol. 1972, 49, 27–35. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons: Chichester, UK, 1997; p. 466. [Google Scholar]
- Giessman, T.A. The biosynthesis of phenolic plant products. In The Biogenesis of Natural Compounds, 2nd ed.; Bernfeld, P., Ed.; Pergamon Press Ltd.: Oxford, UK, 1967; p. 1209. [Google Scholar]
- You, I.S.; Ghosal, D.; Gunsalus, I.C. Nucleotide sequence analysis of the Pseudomonas putida PpG7 salicylate hydroxylase gene (nahG) and its 3′-flanking region. Biochemistry 1991, 30, 1635–1641. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, M.T.; Pfundstein, B.; Haubner, R.; Wurtele, G.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Characterization of alkyl phenols in cashew (Anacardium occidentale) products and assay of their antioxidant capacity. Food Chem. Toxicol. 2006, 44, 188–197. [Google Scholar] [CrossRef]
- Cojocaru, M.; Droby, S.; Glotter, E.; Goldman, A.; Gottlieb, H.E.; Jacoby, B.; Prusky, D. 5-(12-Heptadecenyl)-Resorcinol, the major component of the antifungal activity in the peel of mango fruit. Phytochemistry 1986, 25, 1093–1095. [Google Scholar] [CrossRef]
- Mohan, J.E.; Ziska, L.H.; Schlesinger, W.H.; Thomas, R.B.; Sicher, R.C.; George, K.; Clark, J.S. Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2. Proc. Natl. Acad. Sci. USA 2006, 103, 9086–9089. [Google Scholar] [CrossRef]
- Ziska, L.H.; Sicher, R.C.; George, K.; Mohan, J.E. Rising atmospheric carbon dioxide and potential impacts on the growth and toxicity of poison ivy (Toxicodendron radicans). Weed Sci. 2007, 55, 288–292. [Google Scholar] [CrossRef]
- Kuntze, O. Revisio Generum Plantarum; Arthur Felix: Leipzig, Germany, 1891. [Google Scholar]
- Jelesko, J.G.; Benhase, E.B.; Barney, J.N. Differential responses to light and nutrient availability by geographically isolated poison ivy accessions. Northeast. Nat. 2017, 24, 191–200. [Google Scholar] [CrossRef]
- Benhase, E.B.; Jelesko, J.G. Germinating and culturing axenic poison ivy seedlings. Hortscience 2013, 48, 1–5. [Google Scholar] [CrossRef]
- Aziz, M.; Sturtevant, D.; Winston, J.; Collakova, E.; Jelesko, J.G.; Chapman, K.D. MALDI-MS imaging of urushiols in poison ivy stem. Molecules 2017, 22, 711. [Google Scholar] [CrossRef] [PubMed]
- Baer, H.; Hooton, M.; Fales, H.; Wu, A.; Schaub, F. Catecholic and other constituents of the leaves of Toxicodendron radicans and variation of urushiol concentrations within one plant. Phytochemistry 1980, 19, 799–802. [Google Scholar] [CrossRef]
- Gross, M.; Baer, H.; Fales, H.M. Urushiols of poisonous Anacardiaceae. Phytochemistry 1975, 14, 2263–2266. [Google Scholar] [CrossRef]
- Billets, S.; Craig, J.C., Jr.; Corbett, M.D.; Vickery, J.F. Component analysis of the urushiol content of poison ivy and poison oak. Phytochemistry 1976, 15, 533–535. [Google Scholar] [CrossRef]
- Craig, J.C.; Waller, C.W.; Billets, S.; Elsohly, M.A. New GLC analysis of urushiol congeners in different plant parts of poison ivy, Toxicodendron radicans. J. Pharmacol. Sci. 1978, 67, 483–485. [Google Scholar] [CrossRef]
- Symes, W.F.; Dawson, C.R. Poison ivy “urushiol”. J. Am. Chem. Soc. 1954, 76, 2959–2963. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Adawadkar, P.D.; Ma, C.-Y.; Turner, C.E. Separation and characterization of poison ivy and poison oak urushiol components. J. Nat. Prod. 1982, 45, 532–538. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors, because the compounds were destructively analyzed in the GC-MS, and thus were never isolated as purified compounds. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample: | TMS-Derivatized Cardanol Parent and Fragmentation Ions: | |||
|---|---|---|---|---|
| C15:0 | C15:1 | C15:2 | C15:0d | |
| Cardanols isolated from Cashew [18] | 376 (77) 361 (5) 180 (100) 179 (25) | 374 (42) 359 (2) 180 (100) 179 (22) | 372 (34) 357 (4) 180 (100) 179 (39) | 376 (77) 361 (5) 180 (100) 179 (25) |
| HR32-4a | [C15:0a*] 376 (3) 361 (0) 180 (100) 179 (39) | 374 (23) 359 (2) 180 (100) 179 (39) | Substantial overlap with C15:1a. | B.L.D. |
| C15:1 cardanol authentic standard | [C15:0a*] 376 (2) 361 (0) 180 (100) 179 (31) | 374 (25) 359 (2) 180 (100) 179 (31) | N.A. | N.A. |
| HR32-4a with 1ng authentic C15:1 cardanol spike | [C15:0a*] 376 (2) 361 (0) 180 (100) 179 (28) | 374 (24) 359 (2) 180 (100) 179 (28) | Substantial overlap with C15:1a. | B.L.D. |
| VA-MontCo-1 Seedling | [C15:0a*] 376 (2) 361 (0) 180 (100) 179 (27) | 374 (20) 359 (1) 180 (100) 179 (27) | 372 (14) 357 (2) 180 (100) 179 (40) | 376 (30) 361 (2) 180 (100) 179 (31) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lott, A.A.; Baklajian, E.R.; Dickinson, C.C.; Collakova, E.; Jelesko, J.G. Accession-Level Differentiation of Urushiol Levels, and Identification of Cardanols in Nascent Emerged Poison Ivy Seedlings. Molecules 2019, 24, 4213. https://doi.org/10.3390/molecules24234213
Lott AA, Baklajian ER, Dickinson CC, Collakova E, Jelesko JG. Accession-Level Differentiation of Urushiol Levels, and Identification of Cardanols in Nascent Emerged Poison Ivy Seedlings. Molecules. 2019; 24(23):4213. https://doi.org/10.3390/molecules24234213
Chicago/Turabian StyleLott, Aneirin A., Emily R. Baklajian, Christopher C. Dickinson, Eva Collakova, and John G. Jelesko. 2019. "Accession-Level Differentiation of Urushiol Levels, and Identification of Cardanols in Nascent Emerged Poison Ivy Seedlings" Molecules 24, no. 23: 4213. https://doi.org/10.3390/molecules24234213